Импульсы, синапсы и нейронные сети | Часть 2

Содержание:
Описание
↑ Зрительный путь
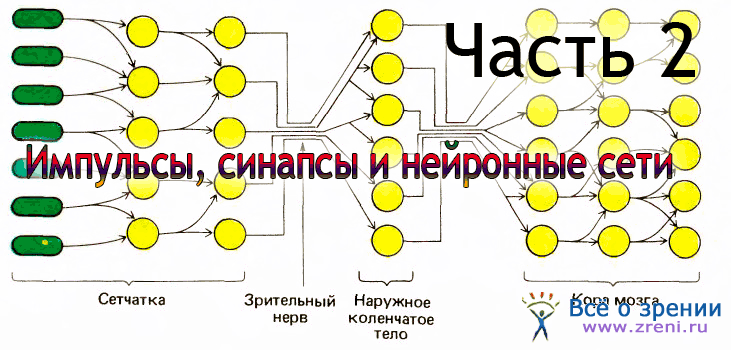
Теперь мы можем приспособить нашу первоначальную схему (рис. 13) к специальному случаю зрительного пути.
Как показано на рис. 14, рецепторы и следующие два уровня находятся в сетчатке. Рецепторами служат палочки и колбочки; зрительный нерв — общий выход всей сетчатки — представляет собой пучок аксонов клеток третьего уровня, называемых ганглиозными клетками сетчатки. Между рецепторами и ганглиозными клетками имеются промежуточные нейроны, из которых наиболее важны биполярные клетки. Зрительный нерв направляется к расположенной глубоко в мозгу промежуточной станции — наружному коленчатому телу. Отсюда после всего лишь одного набора синапсов выходные сигналы направляются к стриарной коре, содержащей три или четыре уровня.

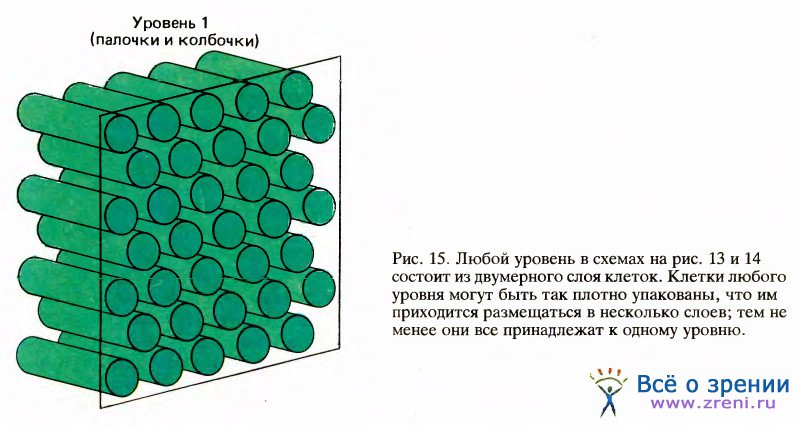
Каждую колонку приведенной схемы можно представить себе как пластину из клеток в поперечном разрезе. Если, например, вы будете смотреть налево от слоя, расположенного на странице справа, то увидите все клетки предыдущего слоя. Каждая клеточная колонка на рисунке представляет собой двумерный массив клеток, подобно тому как это показано для палочек и колбочек на рис. 15.
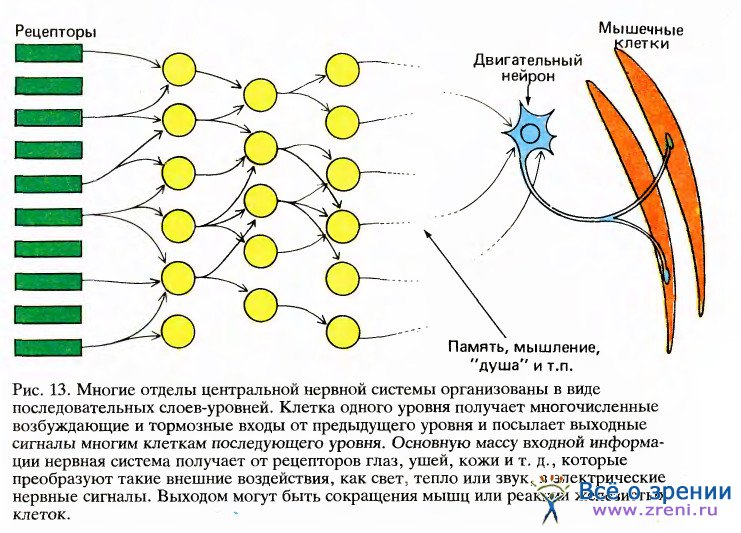
Начиная здесь рассуждать об отдельных уровнях, мы сразу же должны вспомнить о нашем сравнении с генеалогией. В сетчатке, как мы увидим в следующей статье, минимальное число уровней от рецепторов до выхода составляет, несомненно, три, но из-за наличия двух других типов клеток некоторая часть информации проходит непрямыми путями с четырьмя или пятью уровнями от входа до выхода. Для удобства на схеме эти окольные пути, несмотря на их важность, не показаны; кроме того, упрощены также связи между нейронами. Когда я говорю о ганглиозных клетках сетчатки как об «уровне 3 или 4», это не значит, что я забыл, сколько их есть.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Для того чтобы понять характер передачи сигналов в подобного рода сети, мы можем для начала рассмотреть поведение одиночной ганглиозной клетки сетчатки. Из анатомии мы знаем, что такая клетка имеет входы от многих биполярных клеток — возможно от 12, 100 или 1000, каждая из которых тоже в свою очередь получает сигналы от сходного числа рецепторов.
Как общее правило, все клетки, посылающие сигналы к какой-то одной клетке данного уровня (как, например, биполярные клетки, «питающие» одну ганглиозную клетку), тесно группируются вместе. В сетчатке нейроны, соединенные с какой-либо одной клеткой следующего уровня, обычно лежат в зоне диаметром от 1 до 2 миллиметров, и уж во всяком случае не разбросаны по всей сетчатке. Иными словами, внутри сетчатки нет соединений длиннее примерно 1— 2 миллиметров.
Если бы мы располагали подробным описанием всех связей в такой структуре и достаточными познаниями в клеточной физиологии — например, знали, какие синапсы возбуждающие, а какие тормозные, то мы в принципе могли бы вывести заключение о происходящих здесь преобразованиях информации. Однако в отношении сетчатки и коры мозга имеющиеся в нашем распоряжении сведения отнюдь не удовлетворяют этому условию. Поэтому наиболее эффективный подход к решению этой задачи заключался до сих пор в микроэлектродной регистрации активности клеток и в сопоставлении их входных сигналов и выходных реакций. В зрительной системе это равносильно поискам ответа на вопрос: что происходит в клетке, будь то ганглиозная клетка сетчатки или клетка коры, когда глазу предъявляют тот или иной зрительный стимул.
При попытке активировать светом клетку уровня 3 (ганглиозную), нашим первым интуитивным поступком, вероятно, было бы осветить все питающие ее палочки и колбочки ярким светом, направленным в глаз. Именно так полагало большинство исследователей конца 40-х годов, когда физиологи еще только начинали осознавать важность синаптического торможения и никто не понимал, что тормозные синапсы столь же многочисленны, как и возбуждающие. Из-за торможения результат всякой стимуляции решающим образом зависит от того, куда в точности попадает свет, и от того, какие связи являются тормозными, а какие возбуждающими.
Если мы хотим сильно активировать ганглиозную клетку, то едва ли не самое глупое, что можно сделать, это стимулировать все соединенные с ней палочки и колбочки. Стимуляция большим световым пятном или, как предельный случай, освещение всей сетчатки рассеянным светом обычно приводит к тому, что импульсация клетки не учащается и не замедляется, т. е. фактически ничего не происходит: клетка просто сохраняет импульсацию на уровне покоя порядка 5—10 импульсов в секунду.
Для повышения частоты разрядов мы должны освещать некоторое частичное подмножество рецепторов, а именно те из них, которые связаны с данной клеткой (через биполярные нейроны) таким образом, что их влияние окажется возбуждающим. Освещение только одного такого рецептора едва ли может дать сколько-нибудь заметный эффект, но если бы мы осветили все рецепторы с возбуждающим эффектом, то мы были бы вправе ожидать суммирования их влияний и активации клетки — именно так фактически и происходит. Как мы увидим позже, наилучшим стимулом для большинства ганглиозных клеток сетчатки оказывается небольшое световое пятнышко строго определенного размера, вспыхивающее в строго определенном месте. Это, кроме всего прочего, говорит о том, какую важную роль в функции сетчатки играет торможение.
↑ Произвольное движение
Хотя книга будет посвящена в основном начальным, сенсорным уровням нервной системы, я хочу привести два примера, касающихся движения, чтобы дать представление о том, что могут делать конечные уровни схемы на рис. 13.
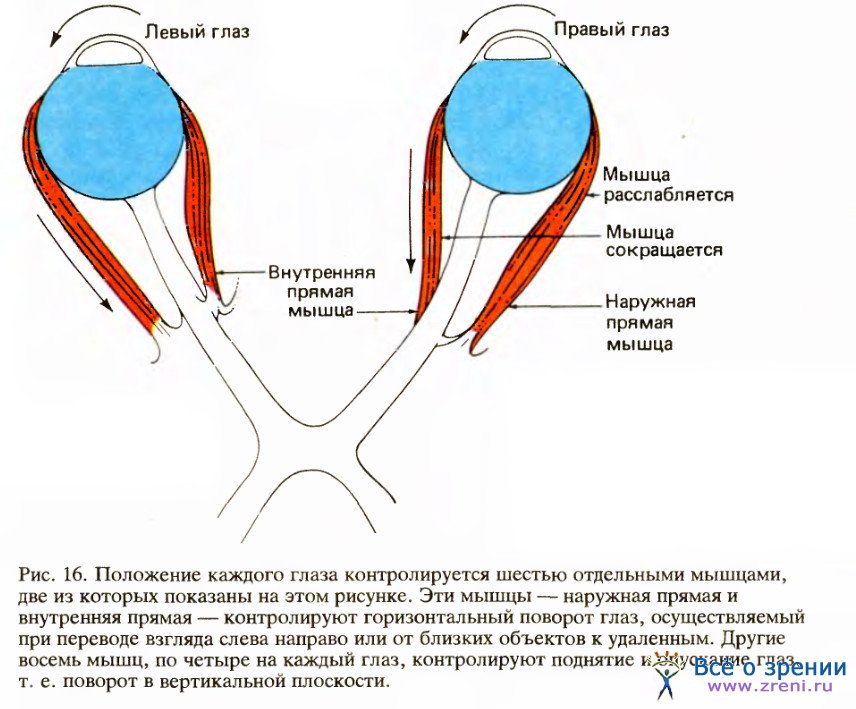
Рассмотрим сначала, как движутся наши глаза. Каждый глаз имеет почти сферическую форму и может свободно поворачиваться подобно шарику в гнезде. (Если бы глазу не надо было двигаться, он вполне мог бы возникнуть в процессе эволюции в виде прямоугольного ящика, похожего на старинный фотографический аппарат.) К каждому глазу прикреплены шесть наружных глазных мышц, которые управляют его движениями.
Для нас здесь не существенно, как все эти мышцы прикрепляются к глазу, но из рис. 16 легко видеть, что для поворота одного из глаз, скажем правого, внутрь к носу человек должен расслабить наружную прямую мышцу и сократить внутреннюю прямую.
Если бы каждая мышца не обладала все время некоторым тонусом, глаз бы свободно болтался в глазнице; поэтому любое движение глаза осуществляется путем сокращения одной мышцы и в точности такого же по величине расслабления мышцы-антагониста. (Это относится и к работе почти всех мышц тела.) Далее, всякое движение одного глаза почти всегда является частью более сложного комплекса движений. Если мы смотрим на объект, расположенный очень близко, оба глаза поворачиваются внутрь; если мы смотрим налево, правый глаз поворачивается внутрь, а левый — наружу; если же мы смотрим вверх или вниз, оба глаза вместе поворачиваются вверх или вниз.
Всеми этими движениями управляет мозг. Каждую глазную мышцу заставляет сокращаться импульсация двигательных (моторных) нейронов, расположенных в участке мозга, который называют мозговым стволом. Каждой из двенадцати мышц соответствует небольшое скопление из нескольких сотен двигательных нейронов ствола. Эти скопления называются глазодвигательными ядрами. Каждый моторный нейрон глазодвигательного ядра обслуживает несколько мышечных волокон одной из глазных мышц.
Эти нейроны в свою очередь получают сигналы от других возбуждающих волокон. Для такого движения, как конвергенция глаз, нужно, чтобы эти предшествующие нейроны посылали ответвления своих аксонов к соответствующим двигательным нейронам, обслуживающим обе внутренние прямые мышцы. Такая одиночная предшествующая клетка могла бы иметь ветвящийся надвое аксон, одна ветвь которого идет к одному глазодвигательному ядру, а другая — к его двойнику на противоположной стороне.
В то же самое время нужно, чтобы другая предшествующая нервная клетка (или клетки), аксон которой имеет тормозящие окончания на двигательных нейронах наружной прямой мышцы, вызывала ее строго соразмерное расслабление. Необходимо, чтобы оба набора предшествующих клеток разряжались совместно, обеспечивая одновременное сокращение и расслабление, а для этого мы могли бы иметь одну управляющую клетку (или группу клеток) на еще более раннем уровне, которая бы возбуждала обе группы. Таков один из механизмов, позволяющих получить координированные движения с участием многих мышц.
Практически каждое осуществляемое нами движение — результат совместного сокращения многих мышц и расслабления многих других. Если вы захотите сжать пальцы в кулак, то мышцы передней поверхности вашего предплечья (на той стороне руки, где ладонь) сократятся, что можно ощутить, положив на предплечье другую руку. (Большинство людей, наверное, думают, что мышцы, сгибающие пальцы, находятся в кисти руки. Кисть действительно содержит некоторые мышцы, но это не сгибатели пальцев.) Как показано на рис. 17, мышцы предплечья, сгибающие пальцы, связаны с тремя косточками каждого пальца длинными сухожилиями, ход которых можно проследить на передней поверхности запястья.
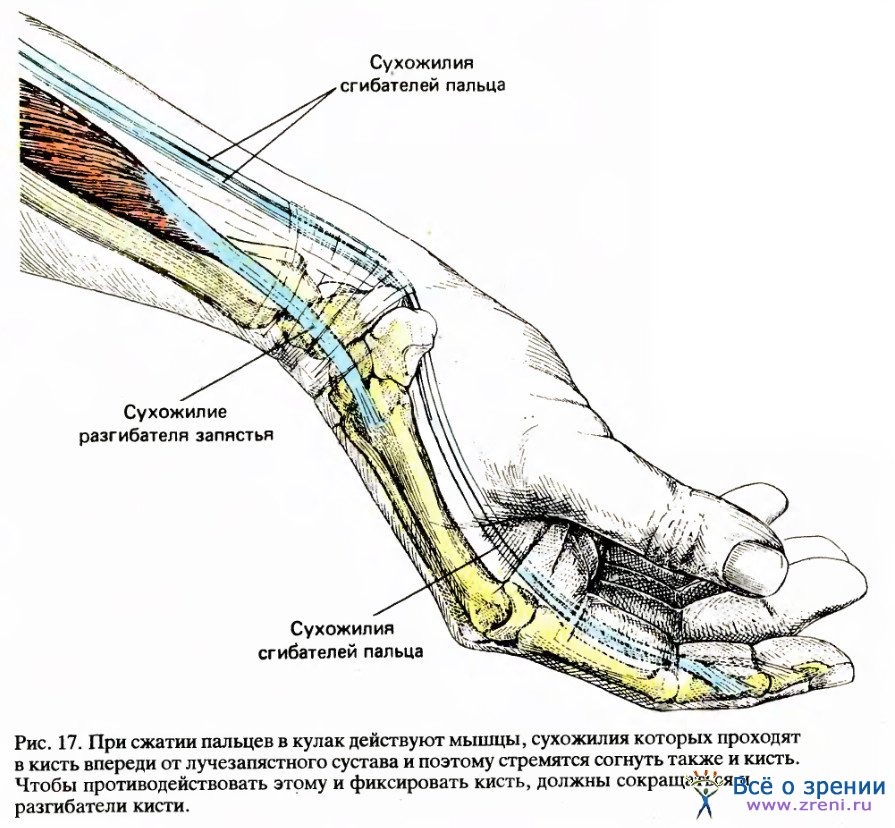
Некоторое удивление может вызвать то, что при сжимании кулака сокращаются также мышцы на тыльной стороне предплечья. Это может показаться совершенно излишним, если не учитывать, что, сжимая кулак, мы хотим жестко зафиксировать запястье в среднем положении: если бы сократились только сгибатели пальцев, их сухожилия, проходя по передней поверхности запястья, согнули бы также и его.
Нужно скомпенсировать эту тенденцию к нежелательному сгибанию запястья путем сокращения мышц, отгибающих запястье назад, а они находятся как раз на тыльной стороне предплечья. Важно то, что мы это делаем не думая. Более того, мы не обучаемся этому, посещая утренние лекции или оплачивая услуги тренера. Новорожденный ребенок хватает вас за палец и крепко держится за него настоящим кулаком без какой-либо предварительной тренировки.
По-видимому, в спинном мозгу у нас есть какие-то клетки исполнительного типа, которые посылают возбуждающие ветви как к сгибателям пальцев, так и к разгибателям запястья, и функция которых состоит в обслуживании процесса сжатия кулака. Видимо, связи этих клеток полностью «запаяны» еще до рождения, так же как и связи клеток, которые позволяют сводить глаза при разглядывании близких предметов, не думая об этом и не обучаясь этому.
---
Статья из книги: Глаз, мозг, зрение | Хьюбел Д.

Комментариев 0