Глаз | Часть 1

Содержание:
Описание

Глаз часто сравнивают с фотоаппаратом. Более уместно было бы сравнить его с телевизионной камерой, установленной на треноге, с автоматической системой слежения — машиной, которая самофокусируется, автоматически подстраивается к интенсивности света, имеет самоочищающуюся линзу и присоединена к компьютеру со столь развитыми возможностями параллельной обработки информации, что инженеры еще только начинают обсуждать сходные стратегии для конструируемой ими аппаратуры.
Гигантская работа по преобразованию света, падающего на две сетчатки, в осмысленную зрительную сцену часто странным образом игнорируется, как будто все необходимое нам для того, чтобы видеть, — это изображение внешнего мира, четко сфокусированное на сетчатке. Хотя получение резких изображений и важная задача, она скромна по сравнению с работой нервной системы — сетчатки и мозга. Как мы увидим в этой главе, вклад сетчатки уже сам по себе впечатляет.
Преобразуя свет в нервные сигналы, она начинает извлекать из окружающей среды то, что полезно, и отбрасывать то, что излишне. Никакое человеческое изобретение, включая управляемые компьютером камеры, пока еще не может соперничать с глазом. Эта глава посвящена главным образом нейронной части глаза — сетчатке, но я начну с краткого описания глазного яблока, аппарата, который содержит сетчатку и создает на ней четкое изображение внешнего мира.
↑ Глазное яблоко
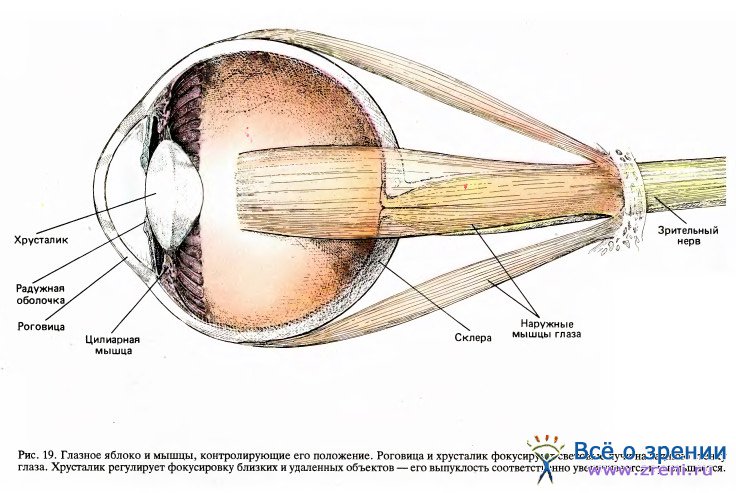
Совокупная функция несетчаточных частей глаз заключается в том, чтобы обеспечить на двух сетчатках сфокусированное четкое изображение внешнего мира. Каждый глаз устанавливается в глазнице в определенное положение шестью маленькими наружными мышцами, упоминавшимися в главе 2. То, что каждый глаз имеет именно шесть мышц, не случайно; они разбиваются на три пары, причем мышцы каждой пары работают в противофазе, обеспечивая движения в трех ортогональных (перпендикулярных) плоскостях. Для обоих глаз задача слежения за объектом должна выполняться с точностью до нескольких угловых минут — иначе видимое изображение будет двоиться. (Чтобы понять, насколько такое двоение может быть мучительным, попробуйте посмотреть на что-нибудь, надавив на край одного из глаз указательным пальцем.) Столь точные движения требуют для своей реализации набора тонко настроенных рефлексов, включая те, которые контролируют положение головы.
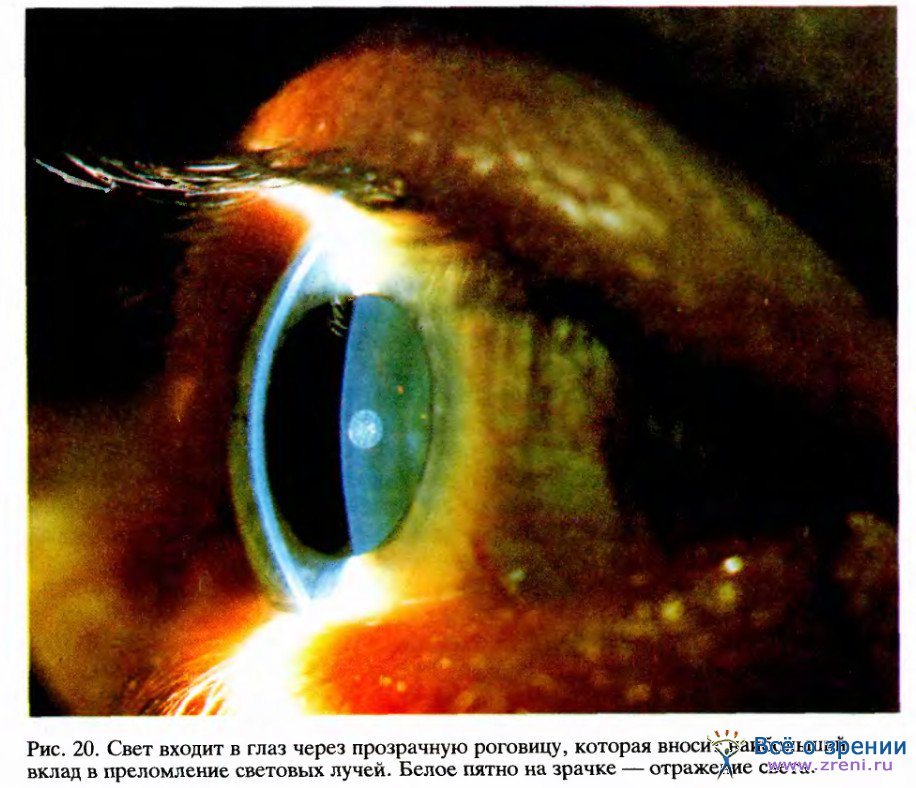
Роговица (прозрачная передняя часть глаза) и хрусталик вместе образуют эквивалент линзы фотоаппарата. Приблизительно две трети общего преломления света, необходимого для фокусировки, происходит на границе воздух — роговица, где свет входит в глаз. Оставшуюся треть фокусирующей способности реализует хрусталик, но его главная задача — обеспечить необходимое регулирование для фокусировки на объектах, расположенных на разных расстояниях от глаза. Фокусируя фотоаппарат, мы изменяем расстояние от линзы до фотопленки; в глазу же изменяется не расстояние от хрусталика до сетчатки, а форма эластичного студенистого хрусталика — путем натяжения или ослабления прикрепленных к его краю сухожилий таким образом, что для близких объектов он делается более выпуклым, а для удаленных — более плоским. Эти изменения формы осуществляет совокупность радиальных мышц, называемых цилиарными мышцами. (Когда мы достигаем примерно 45 лет, хрусталик становится более жестким и мы постепенно утрачиваем способность фокусировать. Чтобы обойти это существенное возрастное неудобство, Бенджамин Франклин изобрел бифокальные очки.) Рефлекс, приводящий к сокращению цилиарных мышц и делающий хрусталик более выпуклым, определяется зрительным входом и тесно связан с рефлексом, контролирующим сопутствующий поворот глаз.
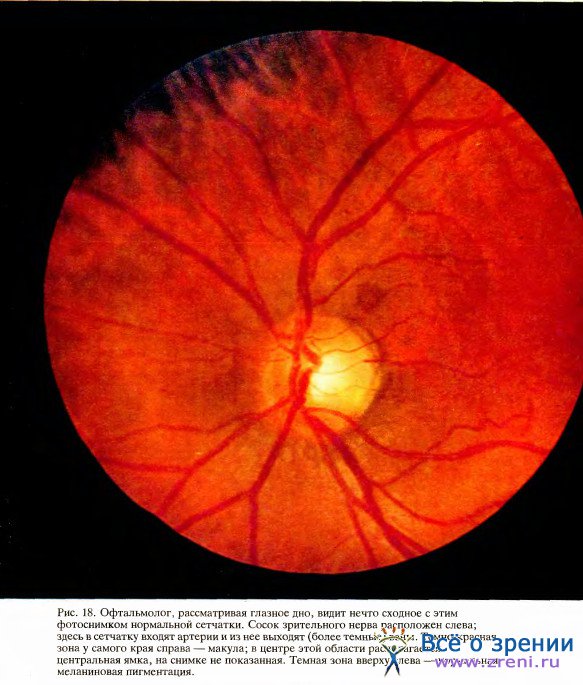
Две другие совокупности мышечных волокон изменяют диаметр зрачка и таким образом регулируют количество света, поступающего в глаз, точно так же, как в фотографическом аппарате это делает диафрагма. Система радиальных волокон, напоминающих спицы колеса, расширяет зрачок; другие, кольцевые волокна сужают его. Наконец, самоочистка передней поверхности роговицы достигается морганием век и смазкой из слезных желез. Роговица обильно снабжена нервами, чувствительными к прикосновению и боли, — вот почему малейшее раздражение пылинками вызывает рефлекс, который ведет к морганию и усиленному выделению слез.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Сетчатка
Вся сложная суперструктура, описанная выше, существует для того, чтобы могла работать сетчатка, которая сама является удивительной структурой. Она преобразует свет в нервные сигналы, позволяет нам видеть в условиях от звездной ночи до солнечного дня, различает длины волн, что дает нам возможность видеть цвета, и обеспечивает точность, достаточную, чтобы заметить человеческий волос или соринку с расстояния в несколько метров.
Сетчатка — это часть мозга, отделившаяся от него на ранних стадиях развития, но все еще связанная с ним посредством пучка волокон — зрительного нерва. Подобно многим другим структурам центральной нервной системы, сетчатка имеет форму пластинки, в данном случае толщиной приблизительно в четверть миллиметра. Она состоит из трех слоев тел нервных клеток, разделенных двумя слоями синапсов, образованных аксонами и дендритами этих клеток.
Слой клеток на задней поверхности сетчатки содержит светочувствительные рецепторы — палочки и колбочки. Палочки, значительно более многочисленные, чем колбочки, ответственны за наше зрение при слабом свете и отключаются при ярком освещении. Колбочки не реагируют на слабый свет, но ответственны за способность видеть тонкие детали и за цветовое зрение.
Число палочек и колбочек заметно изменяется в разных частях сетчатки. В самом центре, где способность нашего зрения различать тонкие детали максимальна, имеются только колбочки. Эту лишенную палочек зону диаметром примерно полмиллиметра называют центральной ямкой. Колбочки имеются по всей сетчатке, но наиболее плотно упакованы в центральной ямке.
Поскольку палочки и колбочки расположены на задней поверхности сетчатки, поступающий свет должен пройти через два других слоя, чтобы их стимулировать. Мы точно не знаем, почему сетчатка устроена таким странным образом — как бы перевернута. Одна из возможных причин — то, что позади рецепторов находится слой клеток, содержащих черный пигмент меланин (он имеется также в коже). Меланин поглощает прошедший через сетчатку свет, не давая ему отражаться назад и рассеиваться внутри глаза; он играет ту же роль, что и черная окраска внутренности фотокамеры. Клетки, содержащие меланин, способствуют также химическому восстановлению светочувствительного зрительного пигмента, который обесцвечивается на свету. Для выполнения обеих функций нужно, чтобы меланин находился поблизости от рецепторов. Если бы рецепторы лежали впереди, пигментные клетки должны были бы располагаться между ними и следующим слоем нервных клеток, в области, уже заполненной аксонами, дендритами и синапсами.
Как бы то ни было, слои перед рецепторами довольно прозрачны и, вероятно, не сильно вредят четкости изображения. Однако на центральном миллиметре, где наше зрение наиболее остро, последствия даже небольшого уменьшения четкости были бы катастрофическими, и эволюция, видимо, «постаралась» смягчить их — сместила другие слои к периферии, образовав здесь кольцо из утолщенной сетчатки и обнажив центральные колбочки так, что они оказались на самой поверхности. Образующееся маленькое углубление и есть центральная ямка.
Двигаясь от заднего слоя к переднему, мы попадаем в средний слой сетчатки, расположенный между палочками и колбочками, с одной стороны, и ганглиозными клетками — с другой. Этот слой содержит нейроны трех типов: биполярные, горизонтальные и амакриновые клетки. Биполярные клетки имеют входы от рецепторов, как показано на рис. 21, и многие из них передают сигналы непосредственно ганглиозным клеткам.
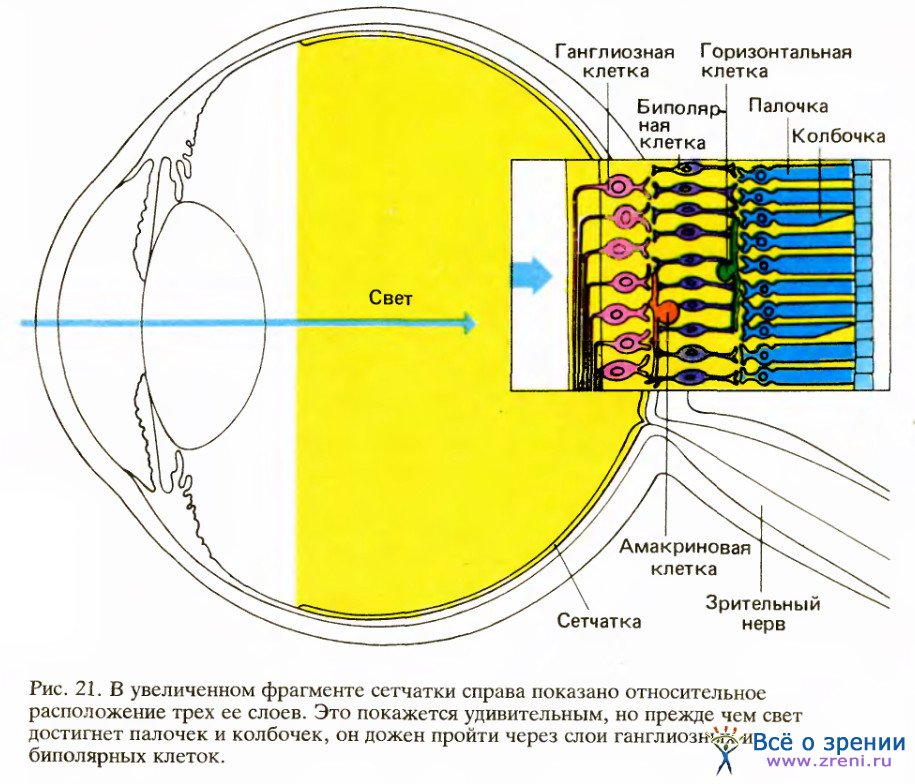
Горизонтальные клетки соединяют рецепторы и биполярные клетки сравнительно длинными связями, идущими параллельно сетчаточным слоям; сходным образом амакриновые клетки связывают биполярные клетки с ганглиозными.
Слой нейронов на передней стороне сетчатки содержит ганглиозные клетки, аксоны которых проходят по поверхности сетчатки, собираются в пучок у слепого пятна и покидают глаз, образуя зрительный нерв. В каждом глазу около 125 миллионов палочек и колбочек, но всего 1 миллион ганглиозных клеток. Ввиду такого различия возникает вопрос: каким образом может сохраняться детальная зрительная информация?
Изучение связей между клетками сетчатки может помочь разрешить эту проблему. Можно представить себе два пути информационного потока через сетчатку: прямой путь, идущий от фоторецепторов к биполярным и далее к ганглиозным клеткам, и непрямой путь, при котором между рецепторами и биполярами могут быть включены еще горизонтальные клетки, а между биполярами и ганглиозными клетками — амакриновые клетки (см. рис. 22, иллюстрирующий эти прямые и непрямые связи).
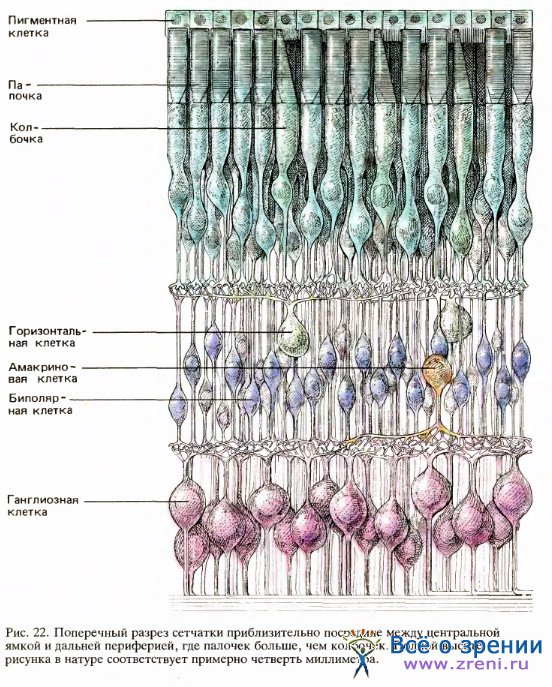
Эти связи уже были весьма обстоятельно изучены Рамон-и-Кахалом примерно в 1900 году. Прямой путь весьма специфичен, или компактен, в том смысле, что одна биполярная клетка имеет входы лишь от одного рецептора или от сравнительно небольшого их числа, а одна ганглиозная клетка — от одного или сравнительно немногих биполяров. Непрямой путь более диффузен, или «размыт», благодаря более широким боковым связям. Общая площадь, занятая рецепторами, связанными с одной ганглиозной клеткой по прямому и непрямому путям, составляет всего около миллиметра. Эта зона, как вы, возможно, помните из статьи Введение в нейронные структуры зрительной системы, является рецептивным полем ганглиозной клетки — областью сетчатки, световая стимуляция которой может влиять на импульсацию данной ганглиозной клетки.
Эта общая схема верна для всей сетчатки, но в деталях связей имеются большие различия между центральной ямкой, куда проецируется направление взора и где наша способность видеть тонкие детали максимальна, и периферией сетчатки, где острота зрения резко снижается. При переходе от центральной ямки к периферии сеть прямых путей от рецепторов к ганглиозным клеткам становится совершенно иной. В центральной ямке или около нее на прямом пути, как правило, одна колбочка связана с одной биполярной клеткой, а один биполяр — с одной ганглиозной клеткой. Однако по мере постепенного перехода внешним областям все больше рецепторов конвергируют на биполярах, а бипо-ляров — на ганглиозных клетках. Эта высокая степень конвергенции, которую мы видим в большей части сетчатки, вместе с весьма компактными путями в самом центре и около него позволяют понять, почему, несмотря на отношение 125:1 между числом рецепторов и числом волокон зрительного нерва, некоторая часть сетчатки (ее центр) может все-таки обеспечивать острое зрение.
Общая схема сетчаточных путей с их прямым и непрямым компонентами была известна многие годы, а их связь с остротой зрения поняли задолго до того, как удалось выяснить роль непрямого пути. Ее понимание внезапно стало возможным, когда начали изучать физиологию ганглиозных клеток.
↑ Рецептивные поля ганглиозных клеток сетчатки: выход глаза
При изучении сетчатки мы сталкиваемся с двумя главными проблемами. Во-первых, каким образом палочки и колбочки преобразуют поглощаемый ими свет в электрические и химические сигналы? Во-вторых, как последующие клетки двух других слоев — биполяры, горизонтальные, амакриновые и ганглиозные — интерпретируют эту информацию? Прежде чем обсуждать физиологию рецепторов и промежуточных клеток, я хочу забежать вперед и описать выходные сигналы сетчатки, представленные активностью ганглиозных клеток. Содержательной, удобной и компактной характеристикой нейрона, а тем самым и информации, доставляемой его выходными сигналами, может служить карта его рецептивного поля. Она может помочь нам понять, почему клетки промежуточных уровней соединены именно так, а не иначе, и объяснить назначение прямого и непрямого путей. Если мы узнаем, что ганглиозные клетки сообщают мозгу, мы значительно продвинемся в понимании работы сетчатки в целом.
Примерно в 1950 году Стивен Куффлер впервые зарегистрировал реакции ганглиозных клеток сетчатки на световые пятна у млекопитающего, а именно у кошки. Он работал тогда в Уилмеровском офтальмологическом институте при больнице Джонса Гопкинса. Ретроспективно можно сказать, что выбор животного оказался удачным, поскольку сетчатка кошки как будто бы не обладает ни сложностью реакций на движение, наблюдаемой у лягушки или кролика, ни особенностями, связанными с цветом, как у рыб, птиц или обезьян.

Куффлер использовал световой стимулятор, сконструированный С. Талботом. С помощью этого оптического прибора — видоизмененного медицинского офтальмоскопа — можно было равномерно освещать всю сетчатку постоянным слабым фоновым светом, а также проецировать маленькие, более яркие пятнышки, непосредственно наблюдая как стимул, так и кончик электрода. Фоновый свет позволял стимулировать либо палочки, либо колбочки, либо рецепторы обоих типов, поскольку при очень ярком освещении работают только колбочки, а при слабом — только палочки. Куффлер отводил реакции внеклеточными электродами, вводимыми через склеру (белая часть глаза) непосредственно в сетчатку с передней ее стороны. Находить ганглиозные клетки было нетрудно, так как они лежат под самой поверхностью сетчатки и довольно крупные.
При постоянном рассеянном фоновом свете и даже в абсолютной темноте большинство ганглиозных клеток сетчатки проявляет стационарную, несколько нерегулярную активность с частотой от 1—2 до примерно 20 импульсов в секунду. Так как можно было бы ожидать, что в полной темноте клетки должны молчать, эта импульсация сама по себе оказалась неожиданной.
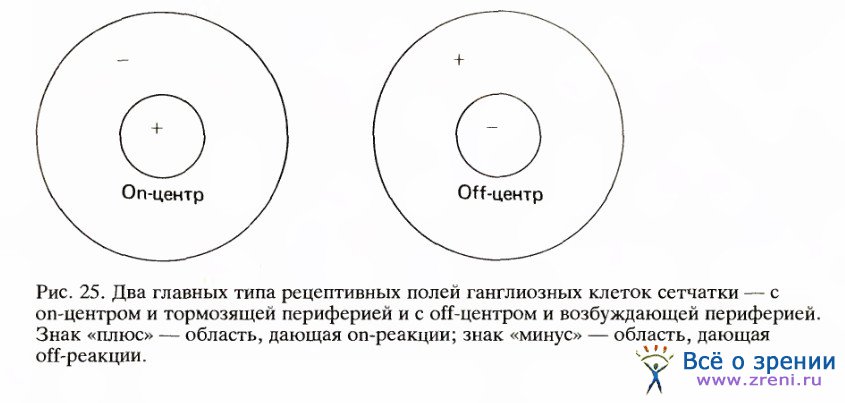
Применяя маленькое световое пятнышко, Куффлер смог отыскивать на сетчатке области, с которых он мог влиять на импульсацию ганглиозных клеток — увеличивать ее или подавлять. Такие области и были рецептивными полями соответствующих ганглиозных клеток. Как и следовало ожидать, рецептивное поле обычно окружало кончик электрода или находилось очень близко к нему. Вскоре выяснилось, что ганглиозные клетки бывают двух типов, и по причинам, которые я скоро объясню, Куффлер назвал их клетками с оn-центром и клетками с off-центром. Клетка с оп-центром разряжается с заметно повышенной частотой, если небольшое пятнышко света появляется где-то внутри определенной зоны в центре или около центра рецептивного поля. Если слушать разряды такой клетки через громкоговоритель, то сначала вы услышите спонтанную импульсацию, отдельные случайные щелчки, а затем, после включения света, возникает залп импульсов, напоминающий пулеметную очередь. Мы назовем эту форму реакции on-реакцией. Когда Куффлер перемещал световое пятнышко чуть подальше от центра рецептивного поля, свет подавлял спонтанную импульсацию клетки, а при выключении света клетка давала залп учащенных импульсов, длившийся около секунды. Такую последовательность — подавление импульсации во время воздействия света и разряд после его выключения — мы называем off-реакцией. Исследование рецептивного поля этого типа вскоре показало, что оно четко подразделено на круглую оп-зону и окаймляющую ее намного большую кольцеообразную off-зону.
Чем большая часть данной зоны, on- или off-, заполнялась стимулом, тем сильнее был ответ, так что максимальные оп-реакции получались на круглое пятно определенного диаметра, а максимальные off-реакции — на кольцо определенных размеров (с определенным внутренним и наружным диаметром). На рис. 24 приведены типичные записи реакций на такие стимулы.
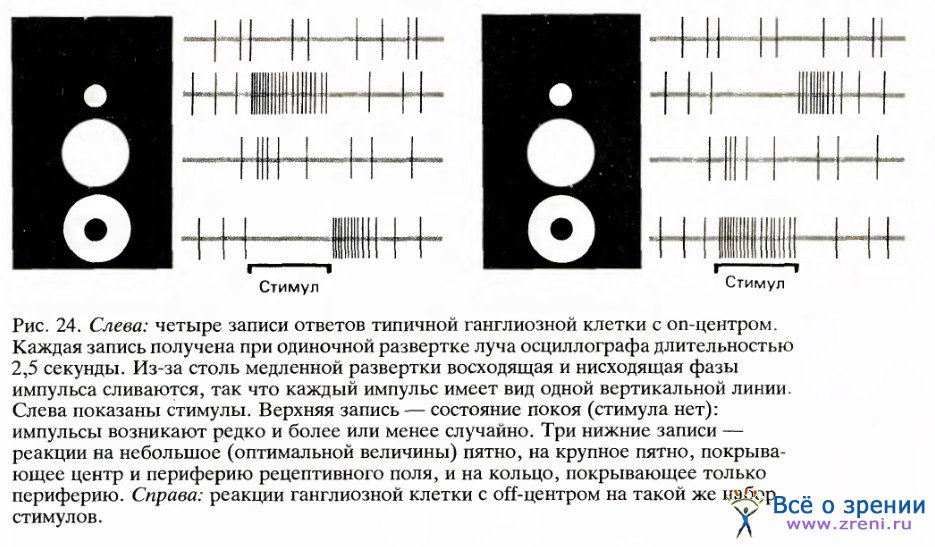
Центральная и периферическая зоны проявляли взаимный антагонизм: реакция на пятно в центре уменьшалась в результате вспышки второго пятна на периферии, как если бы клетку заставляли разряжаться одновременно быстрее и медленнее. Наиболее впечатляющая демонстрация этого взаимодействия между центром и периферией получалась при покрытии всего рецептивного поля одним большим пятном. Это вызывало намного более слабую реакцию, чем при заполнении пятном только центра; для некоторых же клеток эффекты стимуляции обеих зон полностью погашали друг друга.
Прямо противоположным было поведение клетки с off-центром. Ее рецептивное поле состояло из маленького центра, с которого получалась off-реакция, и периферии, дававшей оп-реакцию. Клетки обоих типов были перемешаны и встречались примерно одинаково часто. Клетки с off-центром разряжаются с наибольшей частотой в ответ на черное пятно на белом фоне, поскольку при этом освещается только периферия ее рецептивного поля. В природе темные объекты, по-видимому, столь же распространены, что и светлые; этим можно объяснить, почему информация от сетчатки передается клетками как с оп-центром, так и с off-центром.
Если постепенно увеличивать пятно, реакция возрастает, пока не заполнится центр рецептивного поля, а затем она начинает убывать, по мере того как захватывается все большая и большая доля периферии, как это можно видеть на приведенном графике (рис. 26).

При пятне, покрывающем поле целиком, либо слегка преобладает действие центра, либо реакция нулевая. Это позволяет понять, почему нейрофизиологи до Куффлера были столь неудачливыми: при регистрации активности от ганглиозных клеток они всегда использовали рассеянный свет — далеко не лучший стимул.
Можно представить себе удивление исследователей, когда магниевая вспышка, направленная прямо в глаз животного, вызывала столь слабые реакции или не вызывала их вовсе. Казалось бы, следовало ожидать, что освещение всех рецепторов, гарантируемое при такой вспышке, будет наиболее сильным, а не слабым стимулом. Ошибка здесь — в забвении того, насколько важны для нервной системы тормозные синапсы. Не располагая ничем, кроме схемы связей вроде изображенной на рис. 14, мы не можем предсказать влияние данного стимула на любую данную клетку, если не знаем, какие синапсы возбуждающие, а какие тормозные. В начале 50-х годов, когда Куффлер регистрировал реакции ганглиозных клеток, важность торможения в нервной системе только начинали осознавать.

Прежде чем перейти к описанию рецепторов и других клеток сетчатки, я хочу рассмотреть еще три вопроса относительно рецептивных полей. Первый из них касается общего понятия «рецептивное поле», а два других — некоторых особенностей рецептивных полей ганглиозных клеток сетчатки: их перекрывания и их размеров.
↑ Понятие рецептивного поля
Термин рецептивное поле в узком смысле означает просто совокупность рецепторов, посылающих данному нейрону сигналы через один или большее число синапсов. В зрительной системе это всего лишь некоторая область сетчатки, но со времен Куффлера и благодаря его работам этот термин постепенно стал использоваться в значительно более широком смысле. Ганглиозные клетки сетчатки исторически были первым примером нейронов, рецептивные поля которых обладают внутренней структурой: стимуляция разных частей рецептивного поля дает качественно различные реакции, а стимуляция значительной части поля может приводить к взаимному погашению эффектов от отдельных частей, а не к их сложению. Характеризуя рецептивное поле, в настоящее время обычно описывают его субструктуру, т. е., иными словами, указывают, как нужно стимулировать ту или другую его зону, чтобы вызвать реакцию клетки. Когда мы говорим о «картировании рецептивного поля клетки», мы часто подразумеваем не просто очерчивание его границ на сетчатке или на экране, стоящем перед животным, но также и описание его субструктуры. По мере дальнейшего продвижения в глубь центральной нервной системы, где рецептивные поля нейронов становятся все сложнее, соответственно будет возрастать и сложность их описаний.
Карты рецептивных полей особенно полезны тем, что позволяют предсказывать поведение клетки. Предположим, например, что в ганглиозном слое сетчатки мы стимулируем клетку с оn-центром с помощью светового прямо угольника, ширина которого точно соответствует центру рецептивного поля, а длина больше диаметра всего поля вместе с периферией. По карте для клетки с оn-центром, изображенной на рис. 25, мы могли бы предсказать, что такой стимул вызовет сильную реакцию, так как он покрывает весь центр и лишь небольшую долю антагонистического окружения.
Кроме того, основываясь на радиальной симметрии карты, мы сможем предсказать, что величина реакции клетки не будет зависеть от ориентации световой полосы. Оба предсказания подтверждаются в опыте.
↑ Перекрывание рецептивных полей
С перекрыванием рецептивных полей связан важный вопрос о том, что же делает в ответ на световой стимул некоторая популяция клеток, например выходных клеток сетчатки. Чтобы понять, что делают ганглиозные или любые другие клетки сенсорной системы, мы должны использовать два подхода к проблеме. Картируя рецептивное поле, мы задаемся вопросом, какой нужен стимул, чтобы вызвать реакцию одной клетки. Но мы также хотим знать, как тот или иной конкретный сетчаточный стимул влияет на всю популяцию ганглиозных клеток. Для ответа на второй вопрос следует сначала выяснить, что общего между собой имеют две соседние ганглиозные клетки, расположенные в сетчатке бок о бок.
Данное выше описание рецептивных полей ганглиозных клеток может ввести в заблуждение, если представлять их себе в виде мозаики неперекрыва-ющихся маленьких кружочков на сетчатке, чем-то вроде кафеля, которым выложен пол ванной комнаты. На самом деле соседние ганглиозные клетки получают входные сигналы от сильно перекрывающихся и обычно лишь незначительно разнящихся групп рецепторов (т. е. рецептивных полей), как это схематически показано на рис. 27.

Рассмотрев упрощенную схему на рис. 28, легко понять, чем это обусловлено: ганглиозные клетки, окрашенные красным и голубым, имеют входы от перекрывающихся областей, соответственно окрашенных на поперечном сечении. Вследствие дивергенции, при которой на каждом уровне одна клетка образует синапсы со многими другими клетками, один рецептор может оказывать влияние на сотни или тысячи ганглиозных клеток.
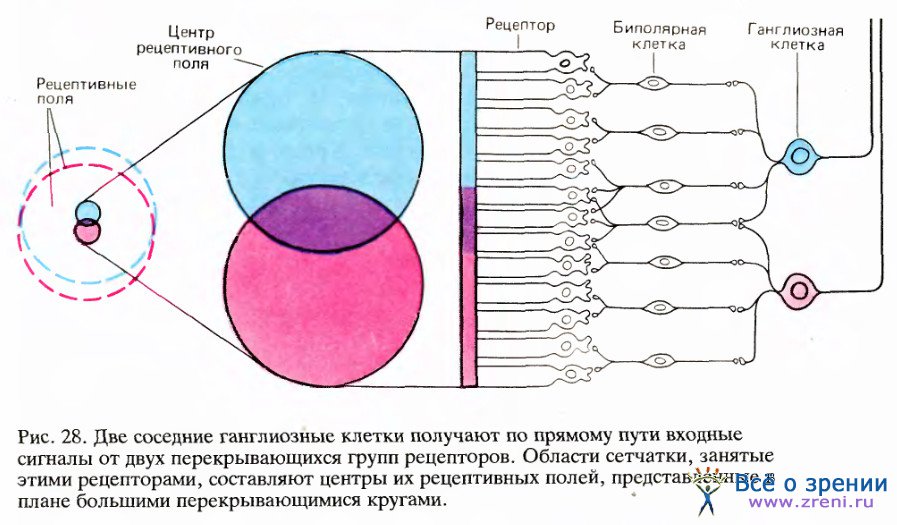
Он будет находиться в центрах рецептивных полей одних клеток и на периферии полей других клеток. Этот рецептор будет возбуждать некоторые нейроны через их центры, если это клетки с оп-центром, или через их периферию, если это клетки с off-центром; и он будет одновременно тормозить другие нейроны через их центры или периферию. Таким образом, маленькое световое пятнышко, появившееся на сетчатке, может вызвать разнообразную активность многих клеток.
---
Статья из книги: Глаз, мозг, зрение | Хьюбел Д.

Комментариев 0