Первичная зрительная кора

Содержание:
Описание
После того как в 1952 году была опубликована первая статья Куффлера о рецептивных полях ганглиозных клеток сетчатки с центром и периферией, открылись пути дальнейшей работы. Чтобы объяснить обнаруженные свойства этих клеток, нужны были дополнительные исследования на уровне сетчатки. Однако требовались также данные о следующих уровнях зрительной системы, чтобы понять, как мозг интерпретирует информацию, доставляемую глазами. Решение этих двух задач было сопряжено с огромными трудностями. При изучении центральных механизмов понадобилось несколько лет, чтобы разработать методику длительной (порядка нескольких часов) регистрации активности одиночной клетки. Еще труднее было подбирать зрительные стимулы, влияющие на эту активность.
↑ Топографическое отображение
До того как стало возможным продолжение работ в этом направлении, уже имелись кое-какие сведения об участках мозга, связанных со зрительной функцией: достаточно хорошо были прослежены начальные этапы передачи зрительной информации (рис. 35).
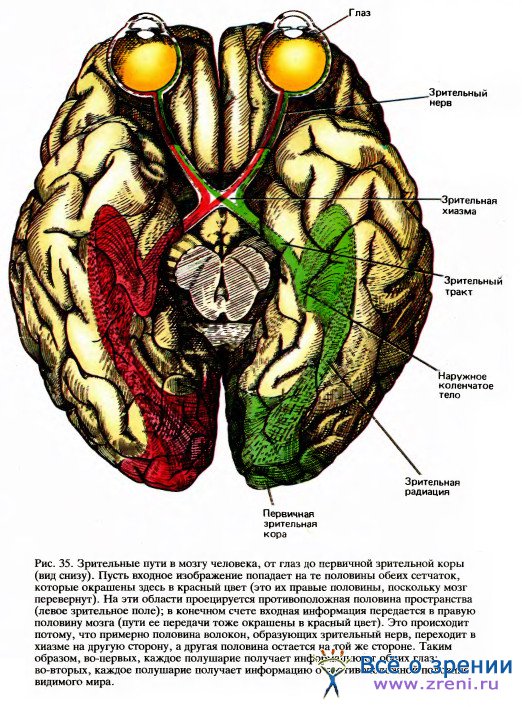
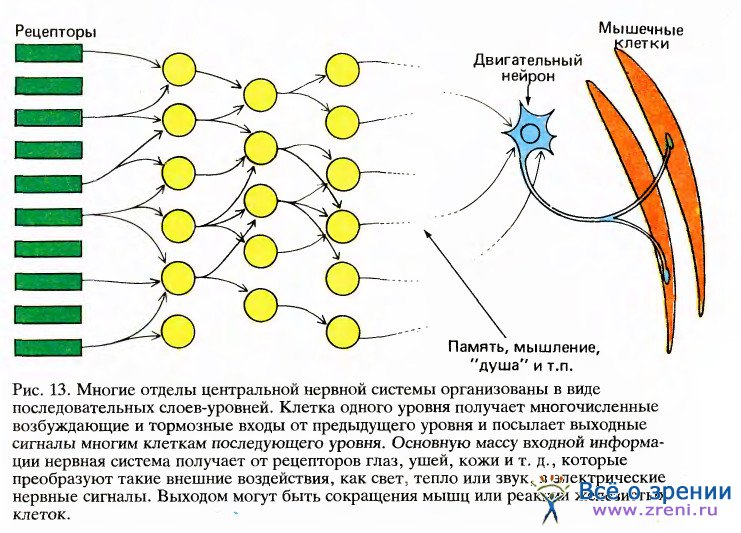
Мы знали, что волокна зрительного нерва образуют синапсы с клетками наружного коленчатого тела (НКТ) и что аксоны клеток НКТ оканчиваются в первичной зрительной коре. Было также ясно, что эти связи — от сетчатки к НКТ и от НКТ к коре — имеют топографическую организацию. Говоря о топографическом отображении, мы имеем в виду, что предшествующая структура проецируется на последующую упорядоченным образом: если идти вдоль какой-либо линии на сетчатке, то проекции последовательных точек этой линии в НКТ и в коре также образуют одну непрерывную линию. Таким образом, волокна зрительного нерва, выходящие из небольшого участка сетчатки, все будут направляться к какому-то небольшому участку НКТ, а все волокна от небольшой зоны НКТ придут в определенную зону зрительной коры. Такая организация связей не покажется удивительной, если мы вспомним упрощенную схему нервной системы на рис. 13: клетки группируются здесь в структуру, напоминающую стопку пластин, причем каждая клетка любой пластины получает входы от некоторой компактной группы клеток предыдущей пластины.
В сетчатке последовательные слои клеток расположены наподобие игральных карт в колоде, так что нервные волокна могут кратчайшим путем проходить с одного уровня на следующий. Клетки наружных коленчатых тел удалены на некоторое расстояние от клеток сетчатки, - точно так же как кора удалена от НКТ и находится в другой части мозга. Тем не менее характер соединений между клетками остается тем же — проекции отдельных зон на более высокие уровни организованы так, как если бы эти зоны точно накладывались друг на друга.
На выходе из глазного яблока волокна зрительного нерва просто собираются в пучок. Дойдя до НКТ, они расходятся и образуют своими окончаниями топографически упорядоченную проекцию. (Удивительно то, что в зрительном нерве на пути от сетчатки к НКТ эти волокна почти полностью перепутываются, но в НКТ снова «находят свои места».) Точно так же и волокна, выходящие из НКТ, расходятся в виде широкой полосы, которая идет через весь мозг к затылку и оканчивается в первичной зрительной коре, где проекция снова оказывается упорядоченной. После того как эти пути, пройдя через первичную зрительную кору и образовав синапсы в различных ее слоях, выходят из этой области и достигают других корковых зон, они опять образуют топографически упорядоченную проекцию. Поскольку на каждом уровне имеет место конвергенция связей, рецептивные поля в целом постепенно становятся все ближе, поэтому чем дальше от сетчатки, тем более размытым будет отображение зрительно воспринимаемой картины.
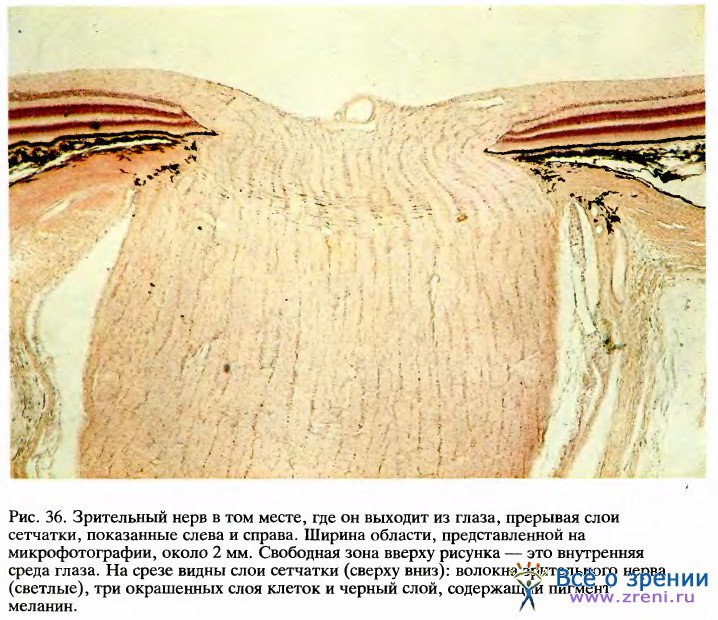
Еще одним важным и давним свидетельством в пользу топографической организации зрительных путей служат клинические наблюдения. Если поврежден определенный участок первичной зрительной коры, то развивается «локальная слепота», как если бы была разрушена соответствующая область сетчатки.
Итак, зрительный мир систематически отображен в структурах наружных коленчатых тел и коры. Однако в 50-х годах было неясно, что может означать такое отображение. В то время было еще не очевидно, что мозг обрабатывает получаемую информацию, преобразуя ее так, чтобы привести к более удобному для использования виду. Полагали, что зрительная сцена просто передается в мозг, а уж его задача — осмыслить ее (или эта задача, быть может, решается вообще не мозгом, а разумом). Из последующих глав мы узнаем, что такая нейронная структура, как первичная зрительная кора, производит глубокое преобразование поступающей информации. Нам почти ничего не известно о том, что происходит на дальнейших этапах ее переработки. Поэтому можно было бы сказать, что мы продвинулись не слишком далеко. Однако знание того, что определенная часть коры использует в своей работе вполне понятные принципы, дает основание для оптимизма — позволяет предполагать, что и остальные зоны коры работают так же. Возможно, настанет день, когда нам вообще не придется употреблять слово «разум».
↑ Ответы клеток наружного коленчатого тела
Волокна, идущие в мозг от каждого глаза, проходят через зрительную хиазму (от названия греческой буквы «хи» — х). В хиазме примерно половина волокон каждого зрительного нерва переходит на противоположную сторону мозга по отношению к данному глазу, а другая половина остается на той же стороне мозга. Пройдя хиазму, волокна направляются в несколько разных пунктов. Некоторые из них идут в нейронные структуры, имеющие отношение к таким специфическим реакциям, как движения глаз и зрачковый рефлекс.
Однако большая часть волокон оканчивается в двух наружных коленчатых телах (НКТ). По сравнению с корой и множеством других отделов мозга эти тела устроены сравнительно просто — все или почти все из примерно полутора миллионов клеток в каждом НКТ имеют непосредственные входы от волокон зрительного нерва, и большинство клеток (но не все) посылают свои аксоны в кору мозга. Отсюда следует, что пути, проходящие через НКТ в кору, имеют только одно синаптическое переключение.
Однако было бы ошибкой считать НКТ просто передаточной станцией. Сюда входят не только волокна из зрительного нерва, но и волокна, приходящие обратно из тех участков коры, на которые проецируются НКТ, а также из ретикулярной формации ствола мозга, имеющей отношение к процессам внимания и общей активации (arousal). Некоторые клетки НКТ имеют короткие аксоны (меньше миллиметра длиной), они не выходят за пределы НКТ, а образуют синаптические контакты с другими нейронами НКТ.
Несмотря на эти усложнения, одиночные клетки НКТ отвечают на световые стимулы примерно так же, как и ганглиозные клетки сетчатки, и у них сходная структура рецептивных полей с on- и off-центрами и сходные ответы на цветовые стимулы. Таким образом, если говорить о переработке зрительной информации, НКТ, по-видимому, не производит никаких значительных преобразований сигналов. Что же касается незрительных входов НКТ и локальных синаптических связей, то пока мы просто ничего не знаем об их роли.
↑ Представительство правой и левой сторон в зрительном пути
Волокна зрительного нерва распределяются между двумя НКТ не совсем обычным и на первый взгляд даже странным способом. Волокна от левой половины сетчатки левого глаза идут в НКТ той же стороны мозга, в то время как волокна от левой половины сетчатки правого глаза переходят в хиазме на другую сторону и, таким образом, попадают в то же левое НКТ (это показано на рис. 35). Аналогичным образом волокна от правых половин обеих сетчаток тоже оканчиваются в одном — правом — полушарии. Поскольку хрусталик создает на сетчатке перевернутое изображение, световые лучи, исходящие из правой половины зрительной сцены, проецируются на левые половины обеих сетчаток и информация передается в левое полушарие.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Термином зрительные поля обозначаются те участки окружающего мира или зрительной сцены, которые видны обоими глазами. Правое зрительное поле включает все точки, лежащие справа от вертикали, проходящей через ту точку, которую мы фиксируем взглядом (рис. 37).

Важно отличать зрительные поля, т. е. то, что мы видим во внешнем мире, от рецептивного поля, которое соответствует тому участку внешнего мира, который «видит» одиночная клетка. Перефразируя то, что говорилось в начале этого раздела, можно сказать, что информация от правой половины поля зрения передается в левое полушарие.
Аналогичным образом устроены и многие другие отделы мозга. Например, осязательные и болевые сигналы от правой половины тела приходят в левое полушарие, а двигательное управление мышцами правой стороны тела осуществляется левым полушарием. Обширный инсульт левой половины мозга приводит к параличу и потере чувствительности правой половины лица, правой ноги и правой руки, а также к потере речи. Но, что менее известно, такой инсульт обычно сопровождается утратой зрения в правой половине зрительного поля, причем для обоих глаз. Чтобы установить наличие такой слепоты, невропатолог просит больного встать перед ним, закрыть один глаз и смотреть другим на кончик носа врача. Затем он начинает обследовать зрительные поля больного, перемещая в разные точки пространства свою руку или специальный зонд.
В случае левостороннего инсульта можно показать, что больной ничего не видит справа от точки фиксации. Например, если врач поднимает в руке зонд и держит его между собой и больным немного выше своей головы, то при медленном движении руки справа налево (с точки зрения больного) больной ничего не видит до тех пор, пока белый рукав халата врача не пересечет среднюю линию и внезапно не появится в поле зрения больного. Если проверить другой глаз, то результат будет точно таким же. Полная правосторонняя гомонимная гемианопсия (так называют неврологи подобное выпадение половины поля зрения!) точно разделит и область центральной ямки (центр взора): если смотреть на слово was, фиксируя взглядом середину буквы а, то не будет видно буквы s, и даже от буквы а останется лишь ее левая половина — интересное, но весьма огорчительное переживание.
Такого рода тесты ясно показывают, что от каждого глаза сигналы передаются в оба полушария и что, наоборот, каждое полушарие мозга получает входные сигналы от обоих глаз. Это может показаться странным: после того что было сказано об осязательной и болевой чувствительности и о системе управления движениями, читатель может предположить, что левый глаз должен посылать сигналы в правое полушарие и наоборот. Однако в случае зрительной системы каждое полушарие мозга имеет дело не с противоположной стороной тела, а с противоположной половиной окружающего пространства. Но, впрочем, ситуация, когда сигналы от левого глаза передаются в основном в правое полушарие (и наоборот), имеет место у многих низших млекопитающих, таких как лошади и мыши, а у птиц и земноводных осуществляется даже полное перекрещивание зрительных нервов. У лошадей и мышег глаза расположены так, что они скорее направлены вбок, чем прямо вперед, в результате чего на большей части сетчатки правого глаза отображается правое зрительное поле, тогда как у приматов глаза направлены прямо вперед и на каждой сетчатке имеются отображения как правого, так и левого зрительного поля. Сделанное выше описание зрительных путей относится только к тем млекопитающим, у которых глаза смотрят более или менее прямо (как, например, у приматов) и поэтому видят почти один и тот же участок окружающего мира.
Сходным образом устроена и слуховая система. Понятно, что каждое ухо способно слышать звуки, исходящие как из левой, так и из правой половины окружающего пространства. Подобно глазам, каждое ухо передает информацию о звуке примерно в равной степени в обе половины мозга. При этом в слуховой системе, как и в зрительной, процесс передачи информации латерализо-ван: звук, доходящий до каждого уха от некоторого источника с правой стороны, анализируется в стволе мозга путем сравнения амплитуд и моментов прихода сигналов к тому и другому уху, и в результате реакция на этот звук формируется в основном в высших отделах левой половины мозга.
Здесь мы говорим о начальных этапах переработки информации. Если справа от меня стоит человек, который словами или жестами побуждает меня сделать что-нибудь левой рукой, то сообщаемая им информация рано или поздно должна попасть в правое полушарие моего мозга. Однако первоначально сигналы должны прийти в слуховую или зрительную кору левого полушария, и только после этого они передаются в моторную кору правого полушария.
Между прочим, никто не знает, почему правая половина окружающего пространства обычно проецируется в левое полушарие головного мозга. Из этого правила есть одно важное исключение — полушария мозжечка (отдел мозга, который в основном управляет движениями) получают входные сигналы главным образом от той же самой, а не противоположной стороны окружающего пространства. Это усложняет работу мозга, поскольку все волокна, соединяющие одно полушарие мозжечка с моторной корой в другом полушарии большого мозга, должны переходить с одной стороны мозга на другую. По поводу такой организации можно пока сказать лишь то, что она кажется загадочной.
↑ Слоистая структура наружного коленчатого тела (НКТ)
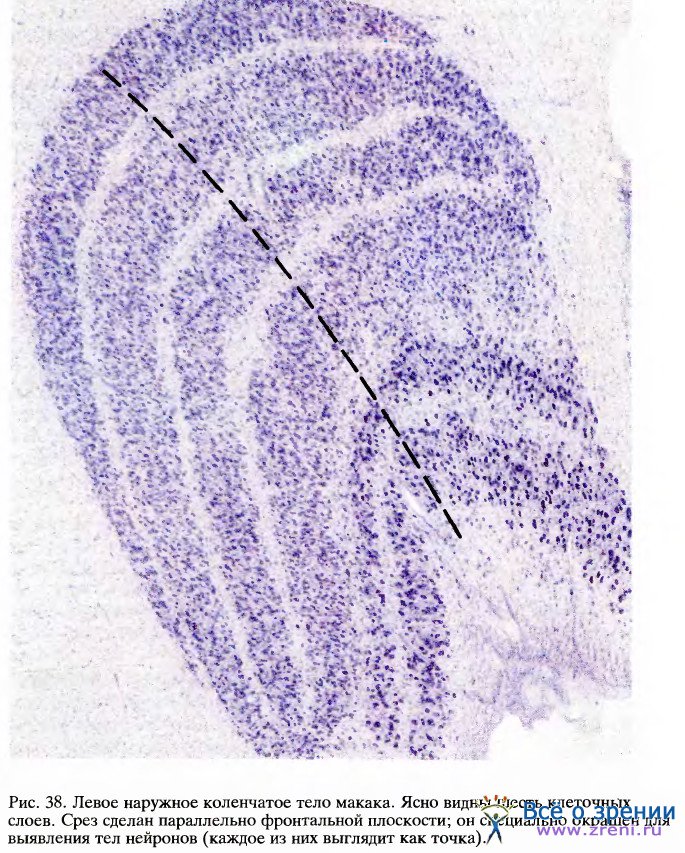
Каждое из НКТ содержит по шесть клеточных слоев. Отдельный слой имеет толщину в несколько клеток (от 4 до 10 и более). Весь этот шестислойный сэндвич согнут таким образом, что его поперечный срез имеет вид, показанный на рис. 38.
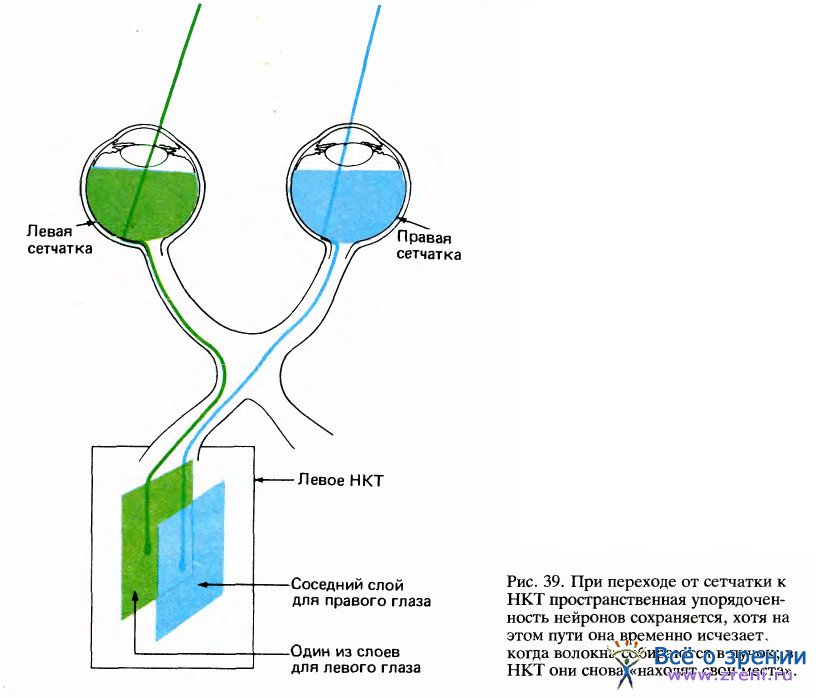
При переходе от сетчатки к НКТ простая схема, в которой каждый последующий слой клеток содержит проекцию предыдущего, становится более сложной. В НКТ проекции от сетчаток двух глаз объединяются, и два отдельных изображения, представленные на уровне ганглиозных клеток в сетчатках, проецируются на шесть слоев НКТ. Волокна от правого и левого глаза не конвергируют на одни и те же клетки НКТ — каждая из этих клеток получает сигналы только от какого-то одного глаза. Два множества клеток разнесены по отдельным слоям, так что в любом слое все клетки получают информацию только от одного глаза.
Эти слои расположены таким образом, что проекции от правого и левого глаза чередуются. Так, в левом НКТ проекции располагаются в следующем порядке (от поверхности в глубину): левая, правая, левая, правая, правая, левая. Не совсем ясно, почему последовательность 5-го и 6-го слоев «перевернута» (иногда мне кажется, что это сделано для того, чтобы порядок проекций было труднее запомнить). У нас пока нет никакого вразумительного объяснения самого факта чередования проекций.
В целом шестислойная нейронная структура имеет одну общую топографию для всех слоев. Левые половины обеих сетчаток проецируются в слои левого НКТ (рис. 39), а правые половины — в слои правого НКТ.
Любая точка в одном слое НКТ соответствует некоторой точке в поле зрения того или другого глаза. Если двигаться вдоль слоя НКТ, то соответственная точка в поле зрения будет перемещаться по некоторой траектории, определяемой характером отображения зрительного поля на НКТ. Если же двигаться перпендикулярно слоям НКТ (например, вдоль черной штриховой линии на рис. 38), как двигался бы микроэлектрод, проходя через разные слои, то соответствующие рецептивные поля клеток оставались бы в одном и том же участке поля зрения; при этом наблюдалось бы только чередование проекций от разных глаз, за исключением, конечно, того места, где идут подряд две проекции от одного глаза. Таким образом, каждая половина поля зрения шестикратно отображается на каждое из НКТ, трижды для каждого глаза, а проекции в слоях НКТ располагаются в точности друг под другом.
Наружное коленчатое тело выглядит как состоящее из двух частей. Его подразделяют на вентральные, или нижние, слои и четыре дорсальных, или верхних, слоя (вентральный — расположенный ближе к брюшной стороне тела, дорсальный — к спинной стороне). Вентральная часть НКТ образует особую структуру, так как клетки в соответствующих слоях отличаются от клеток в остальных четырех слоях — они крупнее и по-иному отвечают на зрительные стимулы. В то же время четыре слоя дорсальной части НКТ как гистологически, так и по своим электрофизиологическим свойствам сходны друг с другом. Поскольку величина клеток в этих двух отделах различна, вентральные слои стали называть крупноклеточными, а дорсальные — мелкоклеточными.
Волокна, выходящие из шести слоев НКТ, объединяются в один широкий пучок, называемый зрительной радиацией, который идет вверх до первичной зрительной коры (см. рис. 35). Здесь эти волокна равномерно расходятся и перераспределяются так, что образуется целостная проекция с топографической организацией (это аналогично распределению волокон зрительного нерва при входе его в НКТ). И вот, наконец, мы подходим к коре.
↑ Ответы клеток в коре
Главная тема этой главы — вопрос о том, как клетки в первичной зрительной коре отвечают на зрительные стимулы. Рецептивные поля нейронов НКТ имеют такую же организацию (разделение на центр и периферию), как и рецептивные поля ганглиозных клеток сетчатки, которые посылают свои аксоны к клеткам НКТ. Подобно ганглиозным клеткам сетчатки, нейроны НКТ различаются между собой главным образом свойствами рецептивного поля (on- или off-центр, местоположение в поле зрения) и особенностями ответов на цветовые стимулы. Возникает вопрос: а как обстоит дело с корковыми нейронами? Сходны ли они с клетками НКТ, посылающими в кору свои аксоны, или же у них появляются какие-то новые особенности? Ответ, как читатель уже должен догадаться, такой: корковые клетки действительно обладают новыми качествами, причем настолько необычными, что вплоть до 1958 года, когда их впервые стали изучать с помощью сложных световых стимулов, никто не мог даже приблизительно предсказать эти свойства.
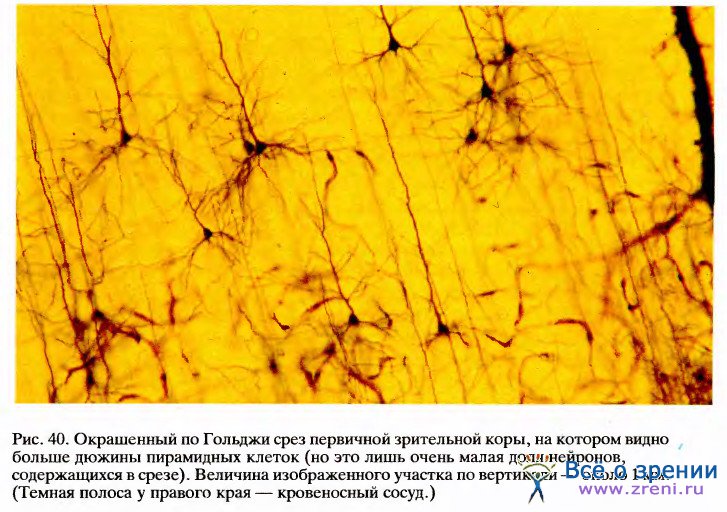
Первичная зрительная кора (стриарная кора) представляет собой слой клеток толщиной 2 мм и площадью в несколько квадратных дюймов1. Для того чтобы дать представление о размерах этой нейронной структуры, можно привести такие цифры: если НКТ содержит полтора миллиона клеток, то стриарная кора — около 200 миллионов клеток. Анатомическая структура стриарной коры удивительно сложна, однако нет необходимости знать ее детали, чтобы понять, каким образом преобразуется здесь поступающая зрительная информация. Более подробно строение этого отдела будет рассмотрено в следующей главе, где будет обсуждаться вопрос о его функциональной архитектуре.
Как я уже говорил, процесс переработки информации в коре состоит из нескольких этапов. На первом этапе большинство клеток даег такие же ответы, как клетки НКТ. Рецептивные поля этих клеток обладают круговой симметрией. Это означает, что линия или граница (перепад освещенности) вызывает один и тот же ответ независимости от ее ориентации. Регистрировать электрическую активность корковых клеток этого уровня непросто, так как они очень малы и расположены близко друг к другу. Пока еще не ясно, отличаются ли вообще ответы этих корковых клеток от ответов клеток НКТ (точно так же как не ясно, отличаются ли реакции клеток НКТ от ответов ганглиозных клеток сетчатки). Сложность гистологического строения НКТ и коры позволяет думать, что между ними должны быть какие-то различия и что их можно будет выявить, если знать, в чем их следует искать; однако узнать это может оказаться трудным делом.
Положение еще больше усложняется, когда мы переходим к ответам клеток на следующем корковом уровне. Вероятно, эти клетки должны получать входные сигналы от нейронов предыдущего уровня, обладающих рецептивными полями с центром и периферией. Вначале нам было совсем не просто выяснить, на какие зрительные стимулы отвечают эти клетки второго коркового уровня. В то время (конец 50-х годов) лишь очень немногие исследователи пытались регистрировать ответы одиночных нейронов зрительной коры.
Те, кто это делал, получали противоречивые результаты. Они нашли, что клетки в зрительное коре работают, видимо, почти так же, как и в сетчатке, — были найдены как on-, так и off-клетки. Кроме того, был обнаружен еще один класс клеток, которые, казалось, вообще не отвечали на световые стимулы. Простота выявленных физиологических свойств корковых клеток на фоне просто дьявольской сложности морфологии коры ставила исследователей в тупик.
Сегодня это очень просто объяснить. Во-первых, применялись неадекватные стимулы — для активации клеток коры по привычке просто освещали всю сетчатку рассеянным светом, хотя Куффлер еще за десять лет до того показал, что такая стимуляция даже для нейронов сетчатки далеко не оптимальна. Для большинства же корковых нейронов диффузный засвет сетчатки — не только не оптимальный, но вообще полностью неэффективный стимул. Если многие клетки наружных коленчатых тел еще реагируют, хотя и слабо, на диффузный засвет, то корковые клетки, даже относящиеся к первому корковому уровню и сходные с клетками НКТ, практически не отвечают на такую стимуляцию. Таким образом, первая приходящая в голову мысль, что для активации зрительных нейронов лучше всего стимулировать все рецепторы сетчатки, оказалась совершенно неверной. Во-вторых (и это еще парадоксальнее), оказалось, что те корковые клетки, которые давали on- или off-ответы, на самом деле были не клетками, а просто аксонами клеток НКТ. Настоящие корковые нейроны в этом случае вообще не отвечали на стимуляцию! Они считали ниже своего достоинства обращать внимание на такой примитивный стимул, как диффузный свет.
Именно так обстояло дело в 1958 году, когда Торстену Визелу и мне удалось осуществить одно из первых успешных отведений электрической активности коры у кошки. В этих опытах положение кончика микроэлектрода в корковой ткани было настолько стабильным, что можно было прослушивать активность одной и той же клетки в течение примерно девяти часов. Чтобы вызывать разряды клетки, мы использовали все возможные и невозможные средства — разве что сами не вставали на голову. (Как большинство корковых клеток, время от времени найденная клетка давала спонтанный разряд, но мы пытались убедить друг друга, что разряд был вызван именно нашими стимулами, и в результате потратили впустую много времени.)

После нескольких часов безуспешных попыток у нас возникло смутное впечатление, что подача света на одну определенную область сетчатки вызывала некоторую реакцию, и мы сосредоточили свои усилия именно на этой области. В качестве стимулов мы в основном использовали круглые белые и черные пятнышки. Чтобы создать черное пятно, мы обычно брали предметное стекло размером 2,5x5 см, на которое был наклеен непрозрачный кружок; стекло вставлялось в специальное проекционное устройство, которое Сэмюэл Талбот сконструировал для подачи изображений на сетчатку. Для предъявления белых пятен мы брали медную пластинку такого же размера с небольшим отверстием (в те времена затраты на проведение исследований были гораздо меньшими).
По прошествии примерно пяти часов напряженной работы нам неожиданно показалось, что, когда время от времени клетка случайно давала реакцию, эта реакция скорее была связана не с черным пятнышком, нанесенным на стекло, а с самим этим стеклом. В конце концов нам удалось установить, что реакцию клетки вызывала слабая, но четкая тень от края стеклянной пластинки, когда ее задвигали в окошко проекционного устройства. Вскоре мы убедились, что этот край работал как стимул только тогда, когда тень от него пересекала определенный участок сетчатки, причем эта тень должна была иметь вполне определенную ориентацию. Самым удивительным было резкое различие результатов в двух случаях — когда ориентация стимула была оптимальной (тогда разряд напоминал пулеметную очередь) и когда мы изменяли ориентацию стимула или производили тотальную засветку глаза яркой вспышкой (в этом случае никакой реакции не было).
Этот феномен был обнаружен самым первым и направил нас по ложному пути. Дело в том, что, как нарочно, обнаруженный тип клеток был как раз тот, который мы впоследствии назвали классом сложных клеток, а они относятся к уровню на две ступени выше, чем первый корковый уровень клеток с рецептивными полями, разделенными на центр и периферию. Хотя сложные клетки — наиболее распространенный тип клеток в стриарной коре, в их свойствах трудно разобраться, не познакомившись сначала с клетками предыдущего, промежуточного уровня.
Действительно, в отличие от клеток первого уровня, имеющих рецептивные поля с центром и периферией, у обезьяны клетки более высоких уровней проявляют совершенно иные реакции. Обычно точечные световые стимулы вызывают лишь слабую реакцию этих клеток или вообще неэффективны. Для того чтобы вызвать ответ клетки, сначала нужно найти надлежащий участок зрительного поля для предъявления стимула, т. е. найти соответствующий участок экрана, перед которым находится животное. Иными словами, сначала мы должны выявить рецептивное поле данной клетки. После этого выясняется, что наиболее эффективным стимулом для данной клетки служит линия, движущаяся в рецептивном поле в направлении, перпендикулярном ориентации линии. Такой линией могла быть узкая полоса света на темном фоне (щель), темная полоса на светлом фоне или же прямолинейная граница между темной и светлой областями. Некоторые клетки отдавали предпочтение, часто очень сильно выраженное, какому-то одному из этих трех стимулов, другие отвечали с примерно одинаковой интенсивностью на стимулы всех трех типов.
Решающее значение имела ориентация линии — чаще всего клетка лучше всего реагировала на определенную оптимальную ориентацию, причем интенсивность ответа (число импульсов, возникавших при пересечении стимулом рецептивного поля) заметно снижалась при отклонении ориентации в любую сторону от оптимальной на 10—20 градусов; при еще большем отклонении реакция круто снижалась до нулевого уровня (см. рис. 41). Указанная цифра 10—20 градусов может показаться очень приблизительной. Вспомните, однако, что разница в положении часовой стрелки, показывающей «час» и «два», еще больше — 30 градусов. Когда ориентация стимула отличается от оптимальной на 90 градусов, типичная клетка, избирательно чувствительная к ориентации, перестает отвечать вообще.
В отличие от клеток на более низких уровнях зрительной системы нейроны, избирательно чувствительные к ориентации стимула, гораздо лучше отвечают на движущиеся, чем на неподвижные линии. Именно поэтому (см. рис. 41) при стимуляции таких нейронов мы использовали линии, движущиеся через рецептивное поле. Если использовать в качестве стимула неподвижную мелькающую (периодически вспыхивающую) линию, то зачастую клетка дает слабый ответ, и в этом случае предпочтительной оказывается такая же ориентация, как и при движущейся линии.
Многие клетки (вероятно, треть всей популяции) дают еще один характерный вид ответа на движущийся стимул. Вместо того чтобы давать один и тот же импульсный разряд независимо от направления движения, такие клетки отвечают более энергично при одном определенном направлении. Бывает даже так, что движение в одну сторону вызывает сильно выраженный ответ, а при движении в противоположную сторону нет вообще никакой реакции (это показано на рис. 41).
За один эксперимент можно оценить реакции 200—300 клеток, если после полного исследования одной клетки просто продвигать микроэлектрод дальше, до следующей клетки. Недостаток такой методики в том, что в одном прогоне можно исследовать только клетки, лежащие в коре на одной прямой линии: как только вы ввели тончайший микроэлектрод в корковую ткань, вы уже не можете сдвинуть его в поперечном направлении, не повредив при этом сам электрод или еще более нежную нервную ткань. С помощью такой методики регистрации самое большее, что мы можем сделать — это исследовать при одно проходке микроэлектрода примерно 50 клеток на миллиметр. Когда мы исследуем ориентационную избирательность нескольких сотен или тысяч клеток, оказывается, что все ориентации стимула встречаются примерно одинаково часто — вертикальная, горизонтальная и все промежуточные, наклонные ориентации. Если учесть характерные особенности окружающего нас мира, в который входят как деревья, так и линия горизонта, то возникнет вопрос: нет ли каких-нибудь выделенных ориентаций, таких как вертикаль и горизонталь, которые встречались бы чаще, чем другие? При попытках ответить на этот вопрос в разных лабораториях получали несколько разные результаты. Однако все исследователи соглашаются, что, если такие предпочтения действительно имеют место, они должны быть очень малы — настолько малы, что для выявления их требуется статистическая обработка данных. А в этом случае они вряд ли имеют какое-либо значение!
В стриарной коре обезьян примерно 70—80% клеток обладают свойствами ориентационном избирательности. Что касается кошек, то у них, по-видимому, все корковые клетки чувствительны к ориентации стимула, даже те, которые имеют прямые входы от наружных коленчатых тел.
Мы обнаружили заметные различия среди ориентационно-специфичных клеток, причем не столько в оптимальной ориентации стимула или в положении рецептивного поля на сетчатке, сколько в характере поведения клеток. Наиболее существенно различие между двумя классами нейронов — простыми и сложными клетками. Как можно догадаться по их названиям, клетки этих двух классов различаются по сложности своих ответных реакций. Поэтому мы сделали естественное предположение о том, что клетки с более простым поведением расположены в нейронной структуре коры ближе к ее входу.
↑ Простые клетки
В большинстве случаев по ответам простых клеток на стимул в виде маленького светового пятна можно предсказать их реакцию на стимул сложной формы. Кажддя из простых клеток, подобно ганглиозным клеткам сетчатки, клеткам НКТ и корковым клеткам с центрально-симметричными рецептивными полями, имеет небольшое четко очерченное рецептивное поле. Предъявление в пределах этого рецептивного поля стимула в виде светового пятнышка вызывает либо on-, либо off-реакцию в зависимости от того, в какой именно участок рецептивного поля подан стимул. Различие между простыми клетками и клетками предыдущих уровней заключается в конфигурации зон возбуждения и торможения. На предыдущих уровнях это центрально-симметричная конфигурация — имеется одна центральная on- или off-зона (возбуждающая или тормозная) и окружающая ее со всех сторон кольцевая зона с противоположными свойствами (тормозная или возбуждающая). Простые клетки коры более сложны. Зоны возбуждения и торможения в их рецептивных полях всегда разделены одной прямой линией или двумя параллельными линиями (рис. 42). Чаще всего встречается такая конфигурация, когда к длинной и узкой возбуждающей зоне с двух сторон примыкают более широкие тормозные зоны (рис. 42, А).

Для того чтобы проверить предполагаемую карту рецептивного поля, составленную путем тестирования маленьким световым пятном, мы попробовали использовать в качестве стимулов другие конфигурации. Вскоре мы выяснили, что чем большую долю той или иной зоны рецептивного поля покрывает данный стимул, тем сильнее выражено возбуждение клетки или ее торможение. Иными словами, имеет место пространственная суммация локальных воздействий. Мы обнаружили также явление антагонизма — взаимного погашения локальных воздействий при одновременной стимуляции возбуждающей и тормозной зоны. Таким образом, для клетки с рецептивным полем, изображенным на рис. 42, А, наиболее подходящим будет стимул в виде узкой полосы, расположенной в рецептивном поле так, чтобы она точно совпадала с возбуждающей зоной и не заходила в тормозную зону (см. рис. 43). Даже незначительное изменение ориентации этой полосы приведет к уменьшению эффективно действующей площади зоны возбуждения и затронет также тормозную зону; в результате частота разряда в ответе клетки уменьшится.
На рис. 42, Б и В показаны рецептивные поля простых клеток двух других типов, которые отвечают наилучшим образом на темные линии и на прямолинейные границы светлого и темного; при этом чувствительность клеток к ориентации стимула остается примерно такой же, как у клеток первого типа. Клетки всех трех типов совсем не реагируют на стимул в виде диффузного освещения. Такое взаимное погашение процессов возбуждения и торможения напоминает реакцию нейтрализации кислоты основанием, которую выполняют студенты в лабораторных работах по химии. Таким образом, уже на этом корковом уровне отмечается большое многообразие нейронов. Если взять класс простых клеток, то у них встречаются три или четыре разных типа рецептивных полей, причем имеются клетки, настроенные на любую из возможных ориентаций стимула, и клетки с рецептивным полем в любом участке поля зрения.
Величина рецептивных полей простых клеток зависит от их расстояния от центральной ямки. Однако в одной и той же зоне сетчатки тоже есть некоторые различия в размерах рецептивных полей. Самые маленькие рецептивные поля, расположенные в центральной ямке и около нее, имеют величину примерно 0,25x0,25°. Что касается клеток типа, показанного на рис. 42, А и Б, то ширина центральной зоны составляет у них не более нескольких угловых минут. Эта величина совпадает с минимальным диаметром рецептивных полей ганглиозных клеток сетчатки или клеток НКТ. Если же взять область далекой периферии, то размеры рецептивных полей простых клеток здесь могут достигать 1x1°.
Даже теперь, спустя двадцать лет, мы все еще не знаем, как устроены входные для корковых клеток цепи, от которых зависят специфические реакции этих клеток. Предложен ряд правдоподобных схем, и вполне может оказаться, что одна из этих схем или какая-то их комбинация окажется верной. Свойства простых клеток могут определяться нейронами предшествующего уровня с круглыми рецептивными полями; проще всего предположить, что простые клетки имеют прямые возбуждающие входы от многих клеток предыдущего уровня — таких, у которых центры рецептивных полей лежат в зрительном поле на одной прямой линии (рис. 44).

Несколько труднее предложить гипотетическую схему для клеток, избирательно реагирующих на границы темного и светлого (см. рис. 42, В). Возможен такой вариант: данная простая клетка имеет входы от двух наборов клеток предыдущего уровня, у которых центры рецептивных полей расположены с двух сторон от одной линии — по одну сторону клетки с on-центрами, а по другую с off-центрами, причем все эти входы возбуждающие. Во всех таких гипотетических схемах возбуждающий вход от клетки с оп-центром по логике вещей эквивалентен тормозному входу от клетки с off-центром, при условии, что клетка с off-центром обладает спонтанной активностью.
Выяснение действительного механизма, определяющего реакции простых клеток, — задача непростая. Относительно данной простой клетки нужно будет узнать, от какого рода клеток она получает входные сигналы, например выяснить для каждой из предшествующих клеток структуру рецептивного поля, его местоположение и ориентацию (если она имеется), тип центра (опили off-), а также характер посылаемых сигналов — возбуждающие они или тормозные. Поскольку методов получения таких сведений пока нет, мы вынуждены пользоваться косвенными подходами, а это увеличивает вероятность ошибки. Схема, представленная на рис. 44, кажется мне наиболее правдоподобной, так как она самая простая.
↑ Сложные клетки
Сложные клетки соответствуют следующему уровню (или уровням) зрительного анализа. Они наиболее многочисленны в стриарной коре и составляют здесь, вероятно, около трех четвертей всей популяции нейронов. Первая из клеток, чувствительных к ориентации стимула, исследованная Визелом и мною, — та, которая отвечала на стимул в виде края стеклянного слайда, — почти наверное была сложной клеткой.
Общим свойством сложных и простых клеток является способность реагировать только на линии, ориентированные определенным образом. Сложные клетки, так же как и простые, отвечают на стимулы, предъявляемые в ограниченном участке поля зрения. От простых они отличаются тем, что реакции их нельзя объяснить формой и распределением возбуждающих и тормозных зон в рецептивном поле. Включение или выключение небольшого неподвижного пятна в пределах рецептивного поля редко вызывает ответ клетки. Даже на надлежащим образом ориентированную неподвижную полосу или границу клетка чаще всего не реагирует или дает лишь слабый, быстро затухающий ответ одного и того же типа как при включении, так и при выключении стимула. Однако, если должным образом ориентированная линия перемещается через рецептивное поле, возникает хорошо выраженный длительный разряд импульсов. Этот разряд начинается в момент, когда линия входит в рецептивное поле, и продолжается до тех пор, пока она не выйдет за его пределы (см. рис. 41, где показана схема ответа).
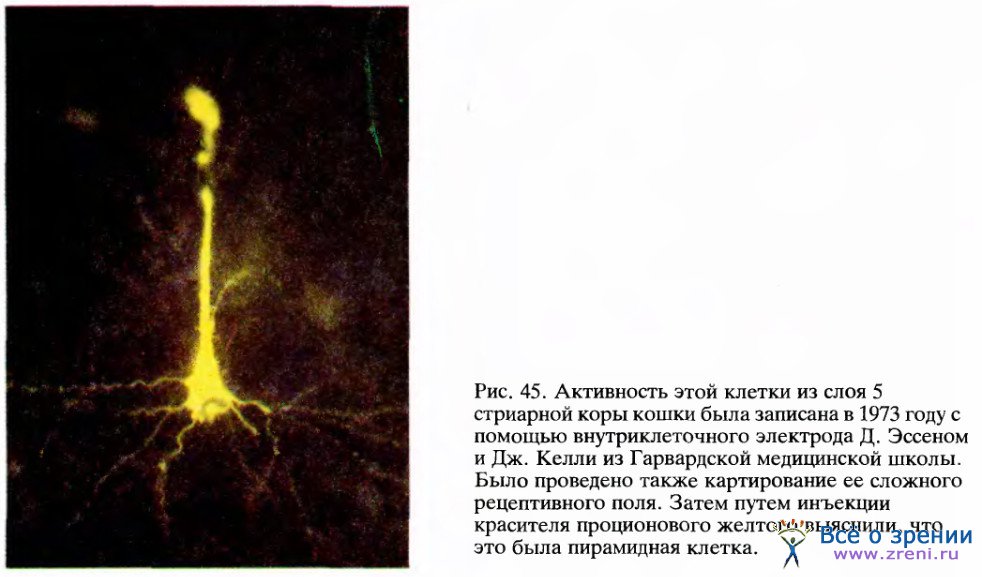
Напротив, для того, чтобы вызвать длительный разряд простой клетки, нужно предъявить надлежащим образом ориентированную неподвижную линию в определенном участке рецептивного поля. Если же использовать движущуюся линию, то возникает только кратковременная реакция в тот момент, когда линия пересекает границу тормозной и возбуждающей зон, или же в то время, когда линия проходит через возбуждающую зону рецептивного поля. Те сложные клетки, которые способны реагировать на неподвижные световые «щели», полоски или границы, дают импульсный разряд независимо от того, в каком месте рецептивного поля расположен стимул, лишь бы ориентация его была подходящей. Однако те же стимулы совершенно неэффективны, если их ориентация далека от оптимальной (рис. 46).

Схемы на рис. 46 (для сложной клетки) и на рис. 43 (для простой клетки) демонстрируют существенное различие между двумя типами клеток: у простой клетки реакцию вызывает оптимально ориентированная линия лишь в очень узком диапазоне положений, а у сложной такая линия вызывает ответ, в каком бы участке рецептивного поля она ни предъявлялась. Это различие связано с существованием четко очерченных возбуждающих и тормозных зон в рецептивном поле простой клетки и с отсутствием их в рецептивном поле сложной клетки. Сложная клетка дает пример генерализации (нелокальности) ответа на линию в пределах более обширной области.
В целом у сложных клеток рецептивные поля несколько больше, чем у простых, но не намного. У макака в области центральной ямки чаще всего встречаются рецептивные поля сложных клеток величиной примерно 0,5x0,5°. В этой области сетчатки оптимальные размеры стимула как для простых, так и для сложных клеток составляют около двух угловых минут. Таким образом, «разрешающая способность» сложных клеток такая же, как у простых клеток.
Как и в отношении простых клеток, мы как следует не знаем, как организована система связей, передающая сигналы сложным клеткам. Однако и здесь нетрудно предложить несколько возможных схем их организации. Согласно простейшей из таких схем, сложная клетка получает входные сигналы от множества простых клеток, рецептивные поля которых имеют одну и ту же ориентацию, но размещены, частично перекрывая друг друга, по всему полю сложной клетки, как показано на рис. 47. Если связи от простых клеток к сложным возбуждающие, то всякий раз, когда в рецептивное поле сложной клетки попадает стимул в виде линии, возбуждаются некоторые простые клетки. В результате будет возбуждаться и сложная клетка.
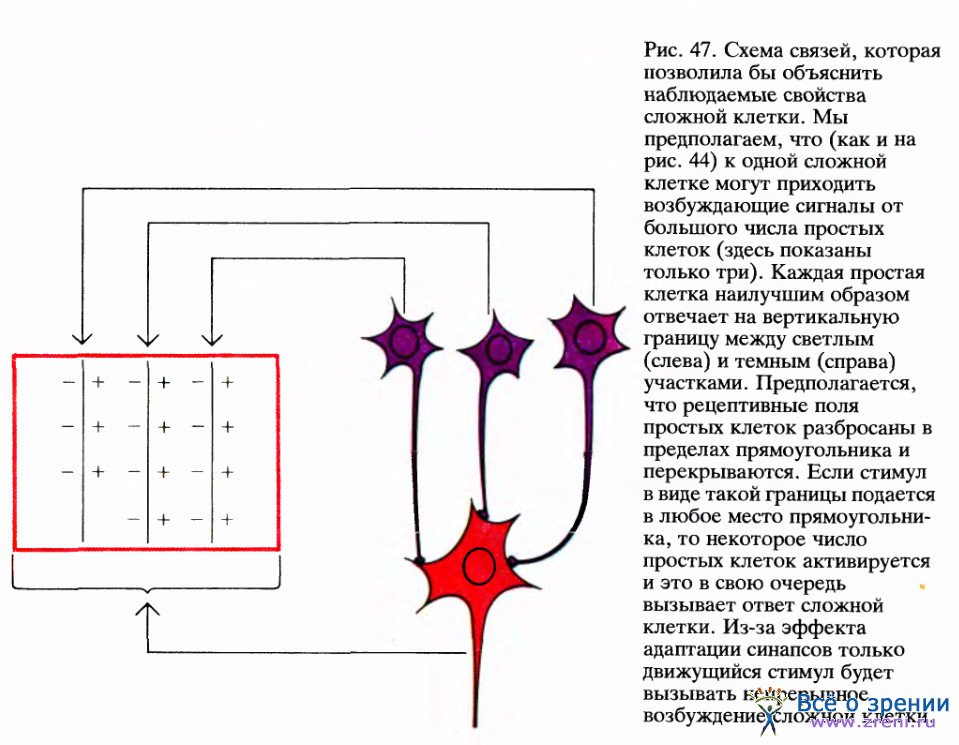
Обычно в ответ на неподвижную линию сложная клетка дает короткий импульсный разряд (даже если стимул остается включенным). В этом случае мы говорим, что происходит адаптация ответа. Если же перемещать линию в рецептивном поле сложной клетки, наблюдается непрерывный разряд: адаптация преодолевается в результате последовательного срабатывания новых простых клеток.
Читатель, должно быть, заметил, что обе приведенные схемы связей — от клеток с круглыми рецептивными полями к простым клеткам (рис. 44) и от простых клеток к сложным (рис. 47) —предполагают использование возбуждающих связей. Однако в этих двух случаях процессы возбуждения должны быть совершенно различными. В первой из этих схем требуется суммирование одновременных сигналов от клеток с круглыми рецептивными полями, лежащих на одной линии. Во второй же схеме для активации сложной клетки движущимся стимулом необходимо последовательное возбуждение множества простых клеток. Было бы интересно выяснить, с какими морфологическими различиями (если они есть) связано это различие в механизме суммации.
---
Статья из книги: Глаз, мозг, зрение | Хьюбел Д.

Комментариев 0