Горизонтальные клетки

Описание

Электронно-микроскопические наблюдения последних лет, а также электрофизиологические микроэлектродные исследования позволили окончательно установить, что горизонтальные клетки — нейроны и отвергнуть гипотезу о глиальной их природе [Максимова, 1970; Бызов, 1971, 1984; Gouras, 1972; Rodieck, 1973; Gallego, 1976; Naka, 1982].
Сходная дискуссия происходила еще в прошлом веке, когда ряд ученых считали горизонтальные клетки опорными или поддерживающими, a Ramon у Cajal [1972] не сомневался, что они типичные нейроны.
Роль горизонтальных клеток полностью не ясна, но совершенно очевидно, что они мультифункциональны. Топографическое положение синаптических окончаний отростков этих нейронов в триадах фоторецепторов обусловливает возможность влияния их на синаптическую передачу от рецептора к биполяру [Бызов, Голубцов, 1978; Byzov, 1979; Бызов, 1984]. Как интернейроны в других отделах нервной системы, горизонтальные клетки воспринимают сигналы от одних элементов — фоторецепторов и передают их различным структурам сетчатки — другим горизонтальным клеткам, обратно фоторецепторам, биполярным клеткам, опосредованно — ганглиозным клеткам.
Соотношение инвагинирующей в фоторецептор биполярной клетки с горизонтальными особенно ясно и детально представлено Sjostrand [1974, 1976]. По данным Sjostrand, от основного инвагинирующего отросточка биполяра отходит крошечное ответвление, входящее в «шейку» синаптического ленточного комплекса фоторецептора и являющееся постсинаптическим к отросткам горизонтальных клеток (см. рис. 3).

Автор считает, что процесс адаптации зависит от тормозного действия горизонтальных клеток на биполярную, осуществляющегося в «шейке» синаптического комплекса фоторецептора.
По Barlow, Levick [1965], горизонтальные клетки играют роль в организации деятельности детекторов направления движения. Из ряда физиологических работ следует, что горизонтальные клетки организуют периферию рецептивных потей ганглиозных клеток [Максимова, 1969; Naka, Nye, 1971; и др.]. Тела горизонтальных клеток ограничивают с внутренней (витреальной) стороны наружный плексиформный слой; отростки пронизывают его, направляясь к фоторецепторам. Продемонстрированы реакции горизонтальных клеток разных слоев: реакции L-тииа генерируются в ближнем к рецепторам слое, хроматические реакции R/G-типа — в более удаленном слое. Mitarai, Asa-no, Miyake [1974] подтвердили многообразие типов горизонтальных нейронов в сетчатке рыб. Авторы идентифицировали 5 типов потенциалов, возникающих в различных слоях клеток. Фотопические реакции L-типа генерируются в наружных горизонтальных клетках. Потенциалы С-типа — красно-зеленые, трехфазные и желто-синие — возникают в средних горизонтальных клетках. Идентификация нейронов проводилась при помощи внутриклеточных меток. Свойства рецептивных полей горизонтальных клеток, возможно, зависят от активности интерплексиформных клеток. Так, разрушение дофаминергических терминалей, свойственных интерплексиформным клеткам и оканчивающихся в наружном плексиформном слое, приводило к увеличению рецептивного поля горизонтальных клеток у карпа [Cohen, Dowling, 1983].
Морфология горизонтальных клеток весьма разнообразна в зависимости от вида исследуемого представителя позвоночных. У рыб наиболее крупные нейроны, образующие подслои или ряды.
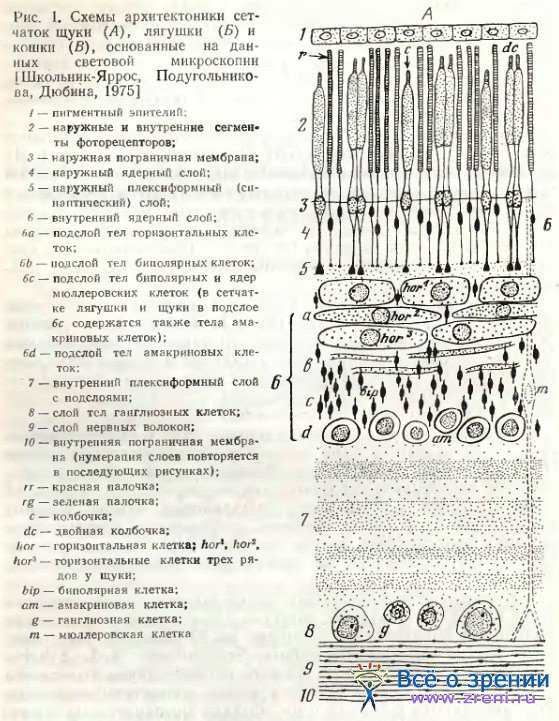
У щуки (см. рис. 1, А) три ряда занимают почти всю ширину внутреннего ядерного слоя. Parthe [1972] в превосходной работе весьма убедительно демонстрирует у Mugil brasiliensis, Eugerres plumieri и других рыб четыре типа горизонтальных клеток — три типа колбочковых и один тип палочковых. У Саrassius auratus, как следует из весьма детального исследования Stell [1975], Stell, Lightfoot [1975], несмотря па наличие лишь узкой прослойки сетчатки в 5—10 мкм толщиной, занятой горизонтальными нейронами, они также четко дифференцированы на три колбочковых и один палочковый типы. Различия в типах касаются размеров тел, обширности ветвлений дендритов и характера синаптических связей. Три типа колбочковых горизонтальных клеток у Carassius auratus обозначены Stell HI, H2, Н3. Каждому типу соответствует особая морфология, сочетающаяся со свойственной лишь этому типу функциональной характеристикой.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Столько же типов горизонтальных клеток в сетчатке черноморской ставриды [Подугольникова, Максимов, 1984]; в сетчатках очень большого количества разнообразных видов костистых рыб три их типа и соответственно ряда [Wagner, 1972; Подугольникова, 1981]. Но у некоторых рыб всего два ряда или даже лишь один ряд горизонтальных клеток, например у зеленушки [Подуголышкова, 1981]. Горизонтальные клетки в сетчатке рыб образуют правильные решетки, элементы которой расположены регулярно, с определенной периодичностью по отношению к периодичности расположения фоторецепторов [Wagner, 1972; Подуголышкова, Максимов, 1973, 1977]. Горизонтальные клетки одного и того же ряда соединяются друг с другом посредством тесных контактов [Yamada, Ishikawa, 19G5; Боровягин, 1966].
Применив Гольджи — электронную микроскопию, Stell, Lightfoot [1975] выяснили избирательность связей определенного типа горизонтальных клеток с колбочками. Так, клетки III-типа, отличающиеся наибольшей величиной тела и наименьшим размахом дендритов, функционально связаны с красно-чувствительными колбочками. Клетки Н2-типа никогда не контактируют с длинными членами двойных колбочек и с длинными одиночными колбочками. Функционально они связаны с зелено- и синечувствительными колбочками. Н3-клетки, имеющие небольшое тело и широкий размах дендритов, контактируют лишь с короткими и миниатюрными одиночными колбочками, т. с. с синечувствительными.
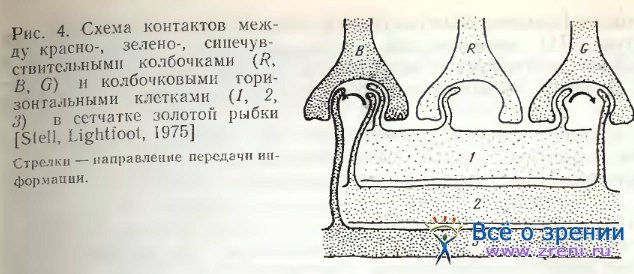
Stell [1975] также указывает на возможные пути передачи сигналов (рис. 4). Так, через Н1-клетки красночувствительные колбочки связываются с Н2-клетками, через Н2-клетки зелено-чувствительные колбочки и Н1-клетки связываются с Н3-клетками. В сетчатке ставриды в отличие от сетчатки золотой рыбки горизонтальные клетки среднего слоя контактируют только с красночувствительными двойными колбочками [Подугольникова, 1981]. У всех колбочковых горизонтальных клеток Carassius auratus Stell [1975] нашел аксоны, спускающиеся в глубь внутреннего ядерного слоя, где они, простираясь в горизонтальном направлении, оканчиваются длинными утолщениями. Эти терминальные утолщения аксона настолько велики, что Ramon у Cajal [1972] принимал их за самостоятельный ряд клеток. У палочковых горизонтальных клеток рыб аксоны не обнаружены.
Терминали аксонов горизонтальных клеток, а также связи по ходу аксонов у многих других видов рыб тщательно прослежены методом Гольджи Parthe [1982]. У наружных горизонтальных клеток (Н1) аксон тонкий, варикозный, затем утолщается и с изгибами спускается и переходит в тонкий отросток с отчетливой пуговкой на конце, примыкающей к телу малой биполярной клетки.
По ходу аксона прослеживаются тонкие нитеобразные отросточки с пуговками, оканчивающимися также вблизи тел биполярных клеток. Аксон горизонтальной клетки Н2 у карповых рыб отходит от одного из больших дендритов, в остальном сходен с аксоном у H1. У внутренних горизонтальных клеток Н3 Parthe видел, помимо основных контактов с фоторецепторами, тонкие отросточки дендритов, оканчивающиеся вблизи тел биполярных клеток. У палочковых горизонтальных клеток импрегнировались два типа отростков — многочисленные толстые восходящие, многократно делящиеся и оканчивающиеся у палочковых сферул; глубокие топкие отростки прослеживаются в горизонтальном направлении и оканчиваются крошечными пуговками вблизи тел крупных биполярных клеток.
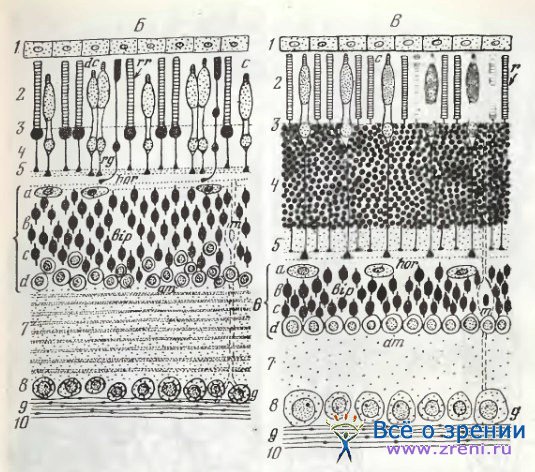
Горизонтальные клетки сетчатки лягушки весьма полно им-прегнированы по Гольджи еще в прошлом веке Ramon у Cajal [1972]. У малых наружных горизонтальных клеток тонкие длинные дендриты и один наиболее топкий очень далеко прослеживающийся отросток, короткие веточки которого оканчиваются небольшими пуговками у ножек фоторецепторов. Терминали этого аксона — 2-3 варикозные восходящие веточки. Внутренние горизонтальные клетки большей величины с короткими многочисленными дендритами, оканчивающимися у ножек фоторецепторов небольшими варикозными ветвлениями. Отчетливый аксон у внутренних горизонтальных клеток также был обнаружен Ramon у Cajal.
Современные исследователи Hassin, Witkovsky [1983], применив метод Гольджи и введение внутриклеточно люциферового желтого у шпорцевой лягушки (Xenopus), полностью подтвердили классические наблюдения о двух типах нейронов и наличии у обоих типов аксонов. По измерениям этих авторов, у малых клеток диаметр тела — 15-17 мкм, дендритные поля — до 50 мкм. У больших клеток соответственно — 20—25 мкм и до 65 мкм. Аксон прослеживался до 800 мкм без густого терминального ветвления. Электрофизиологически определено, что константа длины горизонтальных клеток сетчатки Xenopus превосходит во много раз величину дендритного ветвления (до 15 раз). Функционально горизонтальные клетки связаны с колбочками и палочками (но не с зелеными палочками).
Таким образом, архитектоника (см. рис. 1) и структурная организация горизонтальных клеток в сетчатке многих рыб и некоторых амфибий весьма различна. Можно думать, что структура слоя горизонтальных; клеток этих животных находится в определенной связи с приспособленностью их зрительного анализатора к среде обитания.
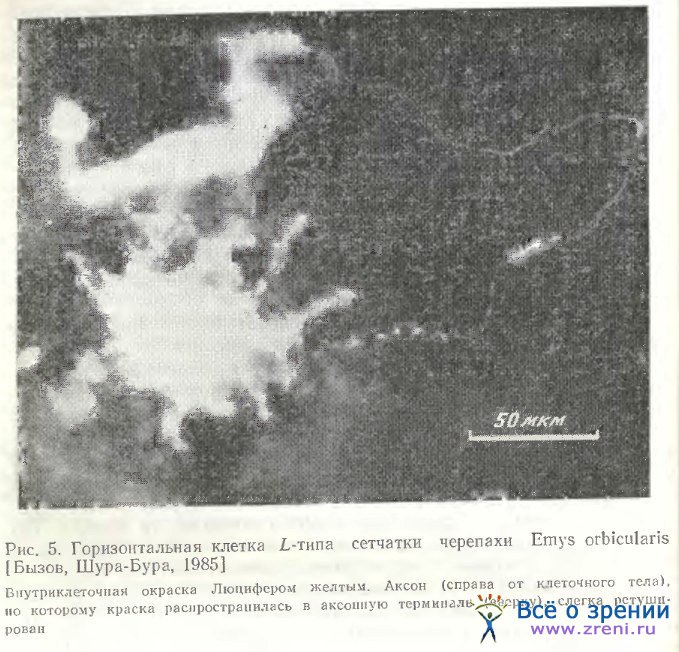
У некоторых видов черепах также обнаруживается 4 типа горизонтальных клеток, но лишь у одного из них (L-типа) найден аксон [Leeper, 1978а]. На рис. 5 представлена горизонтальная клетка сетчатки черепахи Emys orbicularis, окрашенная с помощью внутриклеточного введения люциферового желтого А. Л. Бызовым. Клетка расположена в наружной зоне внутреннего ядерного слоя и относится к типу L. От тела прослеживается тонкий извилистый аксон (справа) и утолщенная терминаль (вверху слева).
Ramon у Cajal различал у всех млекопитающих два типа горизонтальных клеток: наружные, плоские с дендритами, распространяющимися по наружному плексиформному слою, и внутренние, значительно больших размеров. Внутренние горизонтальные клетки подразделяются им далее на имеющие и не имеющие крупные дендритические отростки.

В сетчатке ежа, по нашим предварительным данным,— две разновидности горизонтальных клеток (рис. 6). У более крупных клеток тело плавно переходит в очень широкие густоветвящиеся дендриты, покрытые отчетливыми варикозностями. По ходу тонких отросточков, отходящих от основных дендритов, и на их концах прослеживаются нежные синаптические образования. У горизонтальных клеток меньшей величины тело округлой формы, отростки тоньше и более равномерны по диаметру, чем у крупных клеток. Шипообразные отросточки короче и многочисленнее с типичными синаптическими пуговками на концах. По форме и расположению клеточных тел выявленные нейроны в сетчатке ежа сходны с наружными горизонтальными клетками у млекопитающих по классификации Ramon у Cajal [1972]. В одном случае удалось проследить длинный аксон горизонтальной клетки, распространяющийся по наружному плексиформному слою (рис. 7).

Несколько аксонов у одного из двух типов горизонтальных клеток обнаружил методом Гольджи в сетчатке тупайи (Tupaia glis) Mariani [1985]. Впервые в сетчатке позвоночных автор увидел от 4 до 8 тонких аксонов, начинающихся от концов дендритов. У второго типа горизонтальной клетки морфология отростков не отличается от обычной.
В сетчатке кролика также наблюдаются более крупные и более мелкие горизонтальные клетки. От дендритов горизонтальных клеток в сетчатке ежа и кролика синаптические образования отходят не только по направлению к наружному плексиформному слою, но также в направлении внутреннего ядерного слоя (см. рис. 6). В этих случаях есть основание предполагать существование синаптических связей с отростками других горизонтальных клеток и с отростками биполярных нейронов. Наконец, нельзя исключить возможность связей с телами как горизонтальных, так и биполярных клеток. Действительно электронно-микроскопически синапсы обнаружены между отростками горизонтальных клеток и отростками и телами биполярных клеток в сетчатке кролика и кошки [Dowling, Brown, Major, 1966; Fisher, Boycott, 1974].
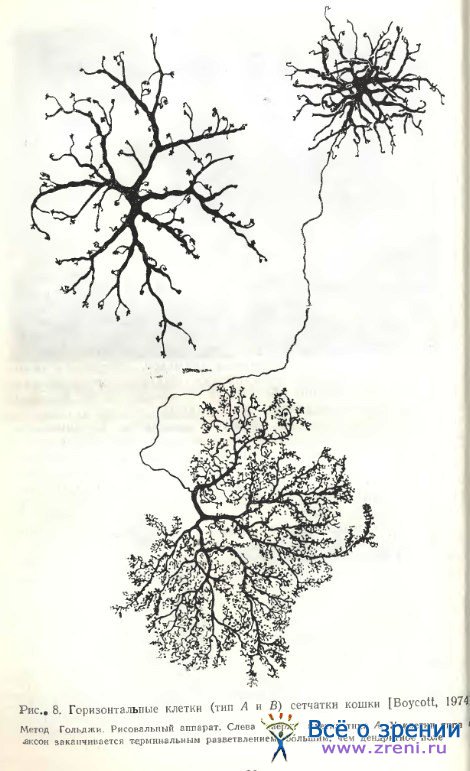
В сетчатке кошки методом Гольджи выделено два типа горизонтальных клеток (рис. 8). Один тип характеризуется отсутствием аксона; у другого аксон обнаружен [Gallego, 1971а; Fisher, Boycott, 1974; Kolb, 1974]. Наиболее детально морфология этих двух разновидностей представлена Fisher, Boycott [1974]. У горизонтальной клетки без аксона дендритное поле достигает на периферии сетчатки 250 мкм в диаметре, дендриты с отходящими от них топкими отросточками оканчиваются скоплениями терминалей. Авторы обозначили подобный нейрон А — типом. У клетки В — типа дендритное поле меньших размеров, отростки тоньше; аксон отходит от одного из дендритов. Терминальное ветвление аксона очень обширное и весьма густое, состоящее из бесчисленного множества мельчайших синаптических пуговок. Подобное терминальное ветвление аксона горизонтальной клетки ранее было обнаружено в сетчатке быка [Ramon у Cajal, 1972], теленка [Marenghi, 1900], собаки [Ramon у Cajal, 1972; Школьник-Яррос, 1958].
Связи дендритов горизонтальных клеток А- и В — типов с фоторецепторами в сетчатке кошки оказались одинаковыми: их окончания образуют латеральные компоненты триад в колбочковых синапсах [Kolb, 1974]. Аксонные окончания у В — типа образуют латеральные компоненты триад в палочковых синапсах. Таким образом, одна горизонтальная клетка В — типа функционально как бы разобщена на палочковую и колбочковую части. Kolb высказывает предположение о возможном смешении палочковых и колбочковых сигналов посредством щелевых контактов, обнаруженных Raviola, Gilula [1973] и другими авторами на уровне самих фоторецепторов, т. е. до их синапсов с горизонтальными клетками. Некоторые различия между двумя типами горизонтальных клеток обнаружены и на ультраструктурном уровне [Fisher, Boycott, 1974]. У клеток A-типа дендриты содержат нейрофиламенты, у клеток В — типа их нет.

На тангенциальных срезах сетчатки кошки нам удалось также наблюдать аксоны у некоторых горизонтальных клеток (рис. 9). Они отходят от одного из дендритов, как и в случаях, описанных Fisher, Boycott [1974]. По ходу аксонов наблюдаются типичные для аксонов в центральной нервной системе варикозности. У одной из импрегнированных клеток (рис. 9) аксон не виден. Однако известно, что метод Гольджи не дает возможности с полной уверенностью судить об отсутствии какого-либо нейрона или его части, в данном случае аксона. Терминальные ветвления аксонов у импрегнированных нами клеток не выявились. У клетки без аксона дендриты толще; возможно, что это A-тип по Fisher, Boycott [1974]. У всех зарисованных клеток заметно асимметричное расположение дендритов. Асимметрия в расположении дендритов горизонтальных клеток может соответствовать асимметрии рецептивных полей. По мнению Normann, Kolb [1980], определенная ориентация дендритных ветвлений горизонтальных клеток и их рецептивных полей может представлять начальные этапы формирования чувствительности к ориентации, полностью развитой в проксимальных отделах сетчатки и высших зрительных центрах.

На радиальных срезах сетчатки кошки наблюдаются тела горизонтальных клеток различной формы (рис. 10) и разного размаха ветвления дендритов. Дендритные веточки оканчиваются множеством синаптических пуговок, в наружном плексиформном слое часть из них явно направляется к фоторецепторам, другие же находятся на уровне распространения дендритов биполярных клеток, а иногда и тел других горизонтальных и биполярных клеток. У 15-дневного котенка заметно незначительное еще распространение дендритов горизонтальной клетки (рис. 11) по сравнению со значительным их размахом у 2-месячного котенка.

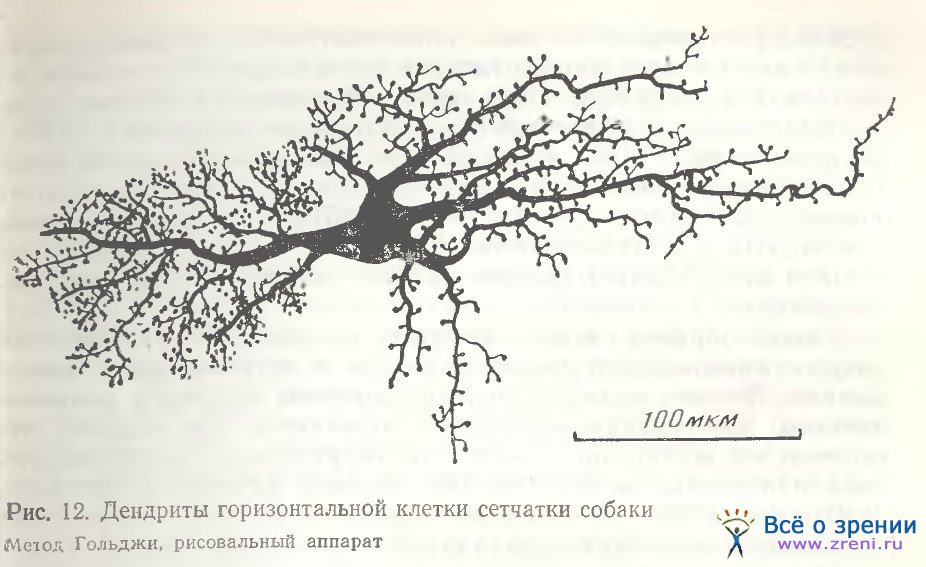
В сетчатке собаки удалось импрегнировать большое удлиненное веретенообразной формы тело горизонтальной клетки (рис. 12) с обилием дендритов довольно крупных, отходящих от тела почти во все стороны, снабженных большим количеством шиповидных отростков с крупными пуговчатыми синаптическими окончаниями. Следует заметить, что сходство представленной нами клетки из сетчатки собаки с внутренней горизонтальной клеткой сетчатки быка [Ramon у Cajal, 1972; рис. 13, табл. VI] очень велико. Аксоны горизонтальных клеток в сетчатке собаки на своем пути к фоторецепторам пересекают соседние аксоны, а конечные их образования, типичные телодендрин, переплетаясь, образуют сплошное, густое и затейливое кружево (рис. 13).
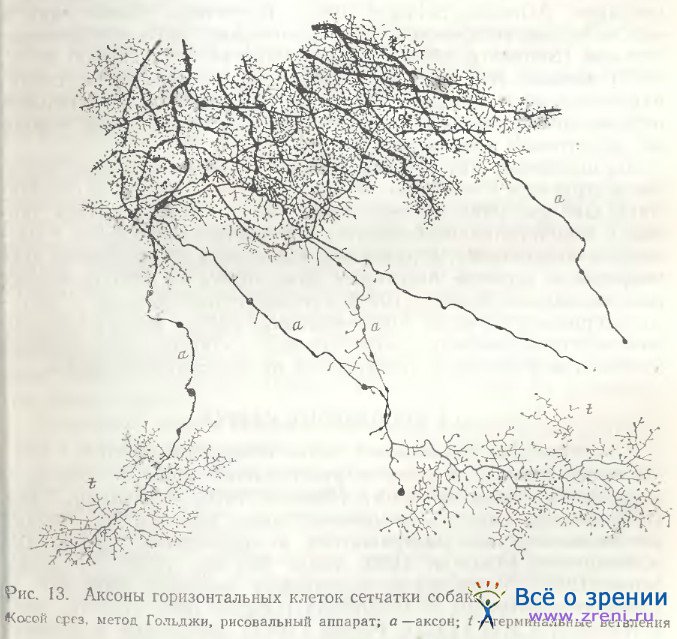
Мелкие синаптические пуговки расположены между крупными стволами аксона. Ячейки, образующиеся между основными ветвями, часто по форме соответствуют телам горизонтальных клеток (рис. 13) [Школьник-Яррос, 1958]. Образованное телодендриями густое кружево может способствовать взаимным связям между аксонами различных нейронов. При электронно-микроскопическом исследовании аксонных терминалей горизонтальных клеток сетчатки птиц действительно выявляется сеть с многочисленными щелевыми контактами. В этих щелевых контактах, однако, постоянно видны и синаптические пузырьки с обеих сторон [Gallego, 1982]. Типичные щелевые контакты наблюдались также между аксонами горизонтальных клеток в сетчатке шпорцевой лягушки [Witkovsky, Powell, 1981]. Проблема — передаются ли потенциалы от тела горизонтальной клетки по длинным весьма топким аксонам к их терминальным ветвлениям — дискутируется и еще окончательно не решена [Бызов, 1984].
В сетчатке приматов наблюдаются две разновидности горизонтальных клеток [Kolb, Mariani, Gallego, 1980], отличающихся формой и расположением дендритных и аксонных ветвлений.
У клеток HI терминали дендритов образуют отчетливые грозди синапсов, у НII от тела клетки отходят 5—6 основных дендритов, распадающихся сразу же па кисточки тонких извитых переплетающихся терминалей. Аксон у горизонтальной клетки первого типа (HI) прямой, редко импрегнирующийся до конечных терминалей, у HII аксон очень топкий и извилистый. Размеры тела и обширность ветвления дендритов изменяются в зависимости от расположения нейрона в центре или па периферии сетчатки. Прослеживая окончания дендритов ДО ИХ синапсов — боковых компонентов триад колбочковой ножки — Boycott, Kolb [1973а] установили, что лишь один компонент триады происходит от исследуемой горизонтальной клетки, другой компонент — от соседней клетки. В fovea одна горизонтальная клетка контактирует с 6—9 колбочками, па периферии сетчатки — с 30 40 колбочками. Аксон горизонтальной клетки (HI) в сетчатке приматов оканчивается в палочках, образуя латеральный компонент палочковой триады [Kolb, 1970]. Терминали аксона клетки НИ в сетчатке приматов оканчиваются в колбочках, по данным Kolb, Mariani, Gallego [1980], именно в синечувствительных.
Таким образом, можно говорить о существенных отличиях морфологии горизонтальных нейронов в сетчатке ряда позвоночных. Значительные отличия обнаружены недавно и физиологически. Как упоминалось выше, в сетчатке рыб найдено несколько типов горизонтальных клеток, раздельно воспринимающих палочковые и колбочковые сигналы [Parthe, 1972; Stell, Witkovsky, 1973]. В сетчатке черепахи (Chelydra serpentina) утолщенные аксонные терминали горизонтальных клеток первого слоя имеют как колбочковые, так и палочковые входы [Copenhagen, Ashmore, Schnapf, 1983]. В сетчатке кошки один и тот же нейрон воспринимает как палочковые, так и колбочковые сигналы [Steinberg, 1969; Niemeyer, Gouras, 1973; Kelson et al., 1975]. Однако Nelson с соавторами считают, что аксон, соединяющий тело и дендриты с густой терминальной аксональной ветвью, лишь разъединяет палочковые и колбочковые компоненты сигналов в сетчатке кошки.
Существенно, что у различных позвоночных нет единообразия в структуре и числе типов горизонтальных клеток [Kolb. Mariani, Gallego, 1980; Gallego, 1982]. Kolb [1974], обобщая данные о структурных особенностях горизонтальных клеток у различных позвоночных, справедливо напомнила об отсутствии этих нейронов в сетчатке некоторых разновидностей глубоководных рыб, по данным Locket [1971]. Следовательно, наряду с другими нейронами сетчатки горизонтальные клетки весьма разнообразны, что, как можно думать, отражает особенности в развитии зрительной функции в зависимости от окружающей среды.
---
Статья из книги: Нейроны сетчатки | Е.Г. Школьник-Яррос, А.В. Калинина

Комментариев 0