Современные данные, обосновывающие и уточняющие теорию трехкомпонентности цветового зрения. Часть 2.


Содержание:
Описание
↑ Кривые основных цветовых возбуждений зрительного анализатора
Таким образом, за цвета, соответствующие трем основным цветовым возбуждениям зрительного анализатора, очевидно, должно признать цвета красный, зеленый и синий. Такой выбор, как мы старались показать, уже однозначно определяется и фактами цветовой слепоты и специальными опытами на лицах с нормальным зрением. В качестве основных цветовых раздражителей зрительного анализатора мы в достаточной мере произвольно могли выбирать различные тройки спектральных цветов и выражать через них любые другие цвета, как результат оптического смешения этих трех, принятых за основные, раздражителей. В отличие от этого, определяя цвета, соответствующие основным цветовым возбуждениям глаза, мы произвола проявлять уже не можем, не впадая в противоречие с фактами. Это обстоятельство и позволяет нам придавать найденным основным цветовым возбуждениям подлинное физиологическое значение.
Определение того, в какой мере возбуждается тот или иной цветоощущающий аппарат зрительного анализатора при воздействии на глаз различных лучей спектра, было произведено впервые Кенигом и Дитерици (1892). Ими построены кривые трех основных цветовых возбуждений глаза, которые должны были конкретизировать основные идеи теории трехкомпонентности цветового зрения. При построении кривых Кениг исходил из полученных им данных о смешении трех принятых за основные цветовых раздражителей, но он учитывал здесь также и цветовые уравнения, принимаемые за равенство цветнослепыми различного вида. Цветовая же слепота (типа девтеранопии и протанопии), в согласии с основными идеями трехкомпонентной теории цветового зрения, принималась им при этом как отсутствие у больного одного из трех цветоощущающих аппаратов.
Основными цветовыми раздражителями в своих опытах со смешением цветов Кениг брал спектральные лучи крайние красные, крайние фиолетовые и зеленые (505 mµ). В качестве же цветов, соответствующих основным цветовым физиологическим возбуждениям глаза, он вынужден был признать пурпурноватый красный, зеленый (505 mµ) и синий (близкий к 470 mµ). На рис. 17

Рис. 17. Кривые трех основных цветовых возбуждений (по Кенигу)
приведены построенные Кенигом кривые трех основных цветовых возбуждений глаза. По ординате отложена возбудимость, по абсциссе — длина волн спектра солнечного света (диффракционного).
В СССР работа по построению трех кривых основных цветовых возбуждений глаза была выполнена оригинальным методом Федоровыми (1935). Эти авторы получили кривые смешения, взяв в качестве основных раздражений спектральные цвета с длинами волн 680, 517 и 480 mµ; кроме того, они вызывали искусственную временную цветовую слепоту (типа протанопии и девтеранопии). Во время искусственной цветовой слепоты авторы могли промерять яркости в спектре, определявшиеся (на значительном протяжении спектра) в этом случае лишь одним («красным» или «зеленым») цветоощущающим аппаратом. Подобным образом оказалось возможным установить кривые «красно-» и «зеленоощущающих» аппаратов. Изменяя чисто расчетным, математическим путем кривые [«красного» и «зеленого», найденные в опытах со смешением цветов, так, чтобы эти кривые сделались возможно близкими к кривым у найденным из опытов с искусственной цветовой слепотой, Федоровы получили возможность установить и вероятный ход третьей «синей» кривой. Построенные Федоровыми на основании согласных данных описанной методики кривые основных цветовых возбуждений глаза приведены на рис. 18.
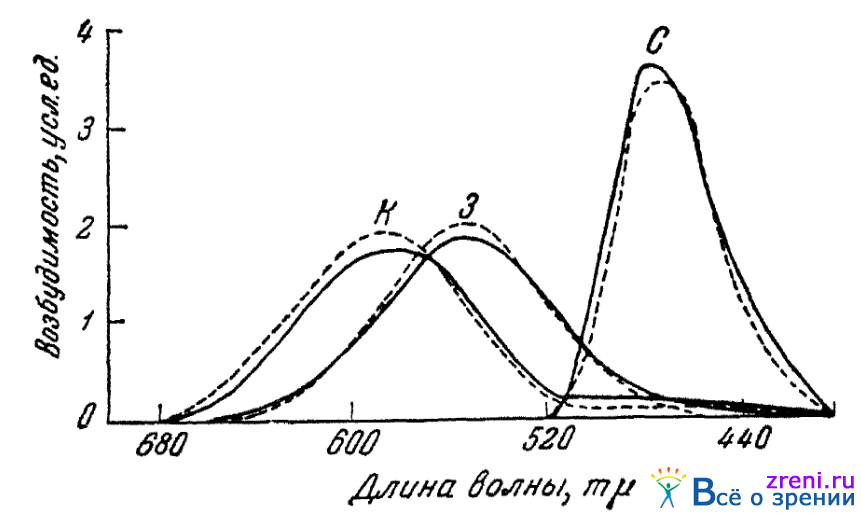
Рис. 18. Кривые трех основных цветовых возбуждений (по Федоровым)
Кривые построены для двух наблюдателей в условиях световой адаптации и фовеального зрения.
В табл. 2 приводятся данные опытов Федоровых.
Кривые основных цветовых возбуждений глаза, найденные Федоровыми (рис. 18), как увидим ниже, достаточно хорошо объясняют многие факты цветового зрения и, повидимому, должны быть признаны в настоящее время наиболее надежными.
Делались попытки определять црета, соответствующие основным цветовым возбуждениям глаза, и иначе: как такие цвета, которые не меняют своего цветового тона при различных условиях адаптации глаза. Райт и Уолтерс (см. Wright, 1946) на специальном приборе установили колориметрическое равенство между цветом, видимым одним глазом, и цветом, видимым другим глазом. По установлении такого равенства они подвергали один глаз цветовой адаптации к тому или другому цветному раздражителю и затем вновь предлагали другим (темноадаптированным) глазом восстановить равенство. Таким образом, они могли наблюдать, изменяется или не изменяется от цветовой адаптации цветовой тон того или другого спектрального излучения. Приближенным образом по этому методу авторы определяли цвета, наименее меняющиеся от адаптации, наносили их в качестве основных на треугольник смешения спектральных цветов и пересчитывали кривые смешения на эти новые основные раздражители, соответствующие, по мнению цитируемых авторов, уже цветам, которые возникали бы при возбуждении отдельных цветоощущающих аппаратов нашего зрительного анализатора. Кривые, построенные Райтом, соответствуют «красному», «зеленому» и «фиолетовому» основным возбуждениям, причем «зеленая» кривая в области синих лучей спектра имеет отрицательное значение. Помимо того, что здесь в качестве основного принимается фиолетовоощущающий аппарат и реальному физиологическому возбуждению зеленоощущающего аппарата в некоторых условиях приписывается отрицательная яркость, сомнения вызывает и применявшаяся методика, поскольку едва ли можно считать, что цветовая адаптация одного глаза никак не сказывается на, цветовой чувствительности другого.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Мало вероятными также представляются и кривые трех основных цветовых возбуждений глаза, предложенные Гехтом (1932). Эти кривые изображены на рис. 19,
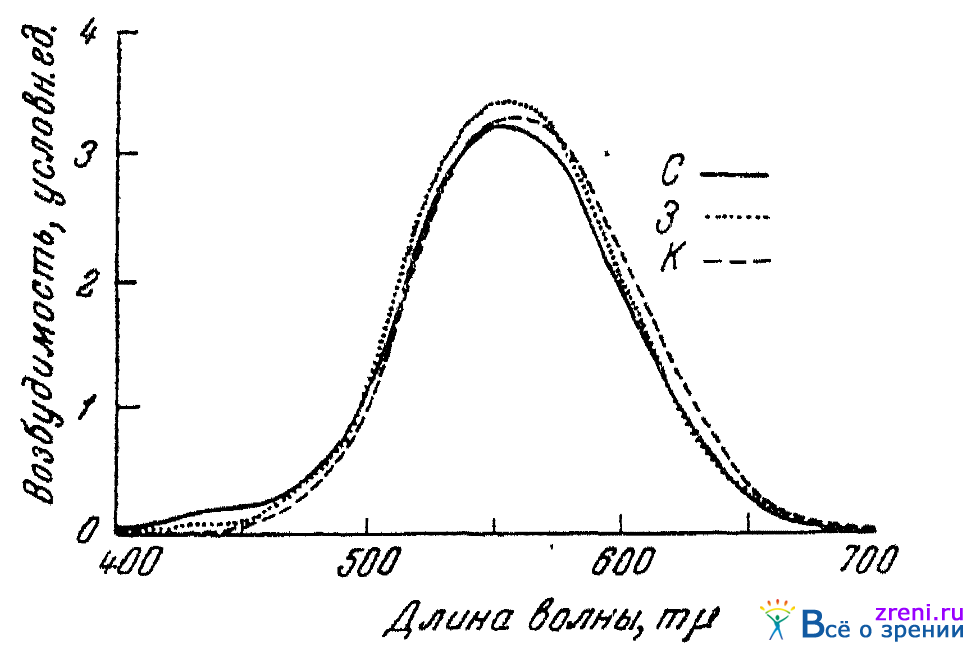
Рис. 19. Кривые трех основных цветовых возбуждений (по Гехту)
где по абсциссе показаны длины волн спектральных лучей равноэнергетического спектра, а по ординате — величина возбуждения красно-, зелено- и синеощущающего аппаратов глаза (в яркостных единицах). Гехт построил свои кривые произвольно с тем расчетом, чтобы из них можно было количественно вывести закономерности смешения цветов, распределения яркости в спектре, различения цветовых тонов и изменения насыщенности по спектру. Несмотря на то, что эти факты цветового зрения из этих кривых с известным приближением и выводятся, приписывать им реальное физиологическое значение все же нельзя. Как видно из рис. 19, каждый из трех цветоощущающих аппаратов, принимаемых Гехтом, возбуждается лучами всех длин волн спектра (от 400 до 700 mµ). Нами проводились многочисленные опыты по выяснению того, как цветовая чувствительность глаза к тем или иным лучам спектра изменяется под влиянием воздействия разного рода непрямых раздражителей (слуховых, обонятельных, электрических и др.) . Эти опыты показали, что колбочковая чувствительность глаза по отношению к концам спектра (лучам фиолетовым и крайним красным) никак не меняется под влиянием тех же непрямых раздражителей, которые явным образом изменяют цветовую чувствительность глаза» по отношению к средней области спектра — зеленым и оранжево-красным лучам. Согласовать этот, твердо установленный, факт с кривыми Гехта нам представляется невозможным, поскольку, по его кривым, и концы спектра возбуждают все три цветоощущающих аппарата глаза. Равным образом не вяжется с этими кривыми и то, что у цветное л епых (девтеранопов и протанопов), по данным наших же опытов, под влиянием непрямых (слуховых) раздражителей изменяется цветовая чувствительность лишь по отношению к синей области спектра, по отношению же ко всем прочим частям его чувствительность при тех же условиях остается неизменной. Как то отмечает и сам Гехт, его кривые не дают объяснения и тому сдвигу места наибольшей яркости в спектре, который наблюдается у девтеранопов и протанопов.
↑ Современные электрофизиологичеекие данные в пользу теории трехкомпонентности
Остановимся сейчас на новейших электрофизиологических опытах Гранита, поскольку именно в них можно видеть более прямое физиологическое доказательство правильности основных положении теории трехкомпонентности цветового зрения. Опыты состояли в регистрации «токов действия» (электрических потенциалов), возникающих в волокнах зрительного нерва при тех или иных раздражениях сетчатки светом. Опыты ставились на наркотизированных животных (лягушках, кошках и др.), у которых передняя часть глаза, до самой сетчатки, была удалена. Экспериментатор прикладывал к обнаженной сетчатке специальный микроэлектрод (диаметром в 25 µ и меньше), что позволяло отводить токи действия от отдельного нервного волокна зрительного нерва или от очень небольшой группы таких волокон. Посредством громкоговорителя или осциллографа определялась пороговая интенсивность того или иного светового раздражителя, впервые вызывавшая электрический ответ в испытываемых волокнах зрительного нерва. Это позволяло измерять чувствительность сетчаточных рецепторов, связанных с данным волокном.
Этим методом было обнаружено, что, как бы экспериментатор ни перемещал микроэлектрод по исследуемой сетчатке у животных, обладающих цветовым зрением, всегда в условиях световой адаптации глаза получаются кривые чувствительности одного из следующих четырех типов. Если по абсциссе откладывать длины волн спектральных лучен, по отношению к которым испытывается чувствительность, а по ординате—относительное значение чувствительности, то первый из этих типов дает широкую, колоколообразную кривую, охватывающую весь видимый спектр и имеющую максимум в желтой области спектра (около 560 mµ). Рецепторы сетчатки, определяющие подобный вид кривой чувствительности. Гранит назвал доминатораыи. В отличие от них, рецепторы, дающие остальные три типа кривых, характеризуются тем, что эти кривые, будучи также колоколообразными, охватывают каждая лишь ограниченную область спектра, имея максимум соответственно или в области красного, или в области зеленого, или в области синего цвета спектра. Такие рецепторы Гранит назвал модуляторами (красно-, зелено- и синеощущающими). Нельзя не видеть здесь определенного фактического подтверждения основных идей трехкомпонентной теории цветового зрения, поскольку при всех своих различиях найденные модуляторы группируются все же именно в три группы с максимумами чувствительности в красном, зеленом и синем.
К установлению такого же факта пришел Гранит и посредством применявшегося им второго метода — метода цветовой адаптации. Сущность его заключается в сравнении кривой чувствительности данного сетчаточного рецептора к различным лучам спектра в условиях темповой адаптации (когда реагирующим аппаратом является зрительный пурпур) с кривой его чувствительности после адаптации к красному, зеленому или синему освещению. В последнем случае ординаты кривой чувствительности по всему спектру часто снижаются не в одинаковой мере. Это свидетельствует о том, что в сетчатке, кроме палочковых рецепторов со зрительным пурпуром, имеются еще и колбочковые рецепторы, снижающие свою чувствительность различно, в зависимости от того, к какому цвету адаптирован глаз. Подсчет вносимого здесь колбочками уклонения от пропорциональности в снижении ординат кривой и позволяет, по Граниту, вычислить кривую чувствительности этих колбочек или кривую чувствительности модуляторов.
На рис. 20
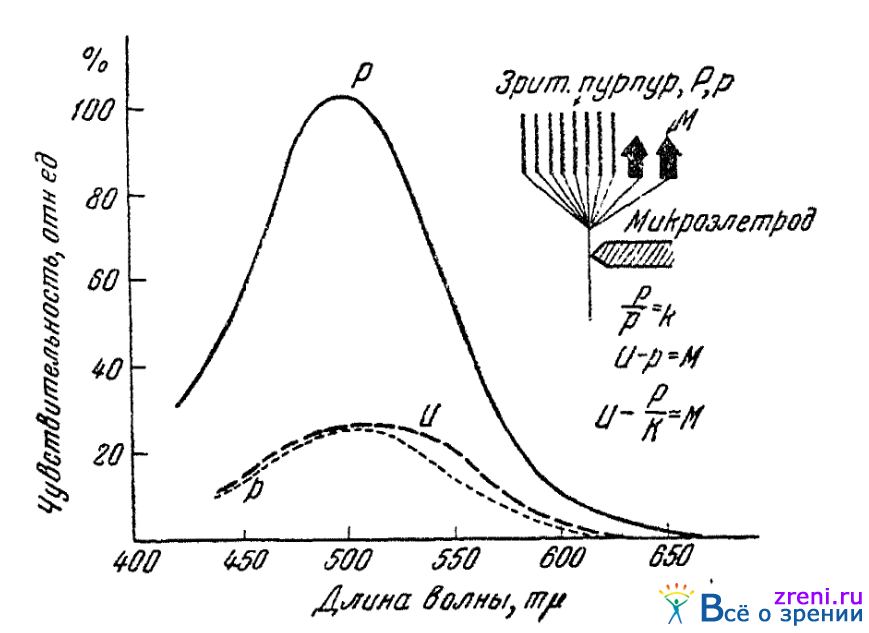
Рис. 20. Схема и результаты применения метода избирательной адаптации
приведена схема Гранита, поясняющая идею описываемого метода. Здесь кривая Р — распределение и величина чувствительности темноадаптированного глаза, когда чувствительность определяется зрительным пурпуром; р — кривая чувствительности, которая получилась бы после цветовой адаптации глаза, если бы с исследуемым волокном сетчатки были связаны рецепторы, содержащие лишь зрительный пурпур. Все ординаты кривой Р в этом случае были бы уменьшены в одинаковом отношении (P/p=k), поскольку уменьшение количества зрительного пурпура относительно одинаково сказалось бы на чувствительности ко всем лучам спектра, способным действовать на зрительный пурпур. Фактически же после цветовой адаптации устанавливается некоторая другая кривая чувствительности, например U. В ней не все ординаты по сравнению с Р снижены одинаково. Это означает, что с исследуемым волокном связаны рецепторы, чувствительность которых зависит не только от зрительного пурпура, но и от некоторых, избирательно чувствительных к различным цветам спектра рецепторам М. Под ними и имеются в виду цветоощущающие аппараты колбочек, называемые модуляторами. На рисунке справа показана схема соединения исследуемого волокна с букетом рецепторов, состоящих из далочек, содержащих зрительный пурпур, и из колбочек-модуляторов, содержащих в себе вещество уже с иным спектром поглощения. Справа жена рисунке показан и ход вычисления чувствительности этих модуляторов М.
Следует особо отметить, что посредством обоих электрофизиологических методов Гранит пришел к одним и тем же результатам. Все наблюдавшиеся им модуляторы, как мы уже отмечали, распределяются в три группы, с максимумами чувствительности, приходящимися на области красного (ок. 600 mµ), зеленого (ок. 530 mµ) и синего (ок. 460 mµ). На рис. 21 б
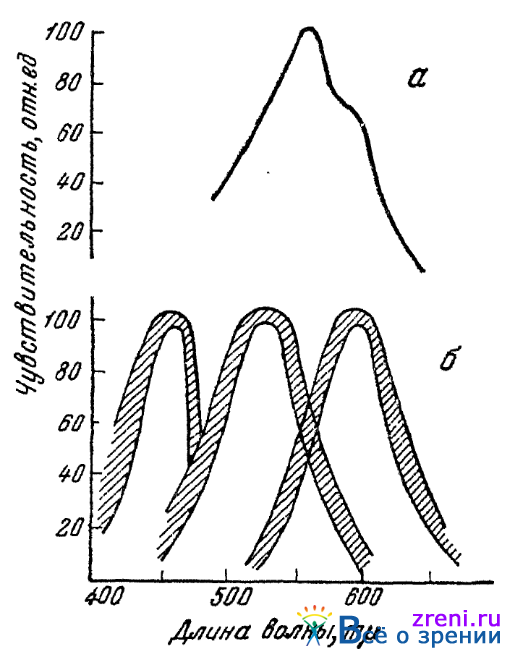
Рис. 21. Кривые доминаторов (а) и модуляторов (б) (по Граниту)
приведены кривые модуляторов для глаза кошки. Двойными кривыми обозначена область рассеяния отдельных опытных данных. По ординате отложена чувствительность, по абсциссе — длина волны лучей равноэнергетического спектра. Нельзя не отметить сходства этих кривых по положению максимумов с приведенными выше кривыми трех основных цветовых возбуждений, найденными для человеческого глаза совсем иными методами (см. рис. 17 и 18).
Итак, уже имеется ряд фактов, не только подтверждающих правильность основных положений теории трехкомпонентности цветового зрения, но и позволяющих с известной степенью вероятности высказать предположение, каким цветам соответствуют три основных цветовых возбуждения глаза и как эти возбуждения друг с другом сочетаются при воздействии на глаз тех или иных лучей спектра.
↑ О равных видах колбочек
Несмотря на только что сказанное, в природе цветового зрения остается еще очень много неясного. Пока нет анатомо-гистологических доказательств того, что колбочки сетчатки бывают троякого строения или имеют, каждая, троякого рода рецепторные аппараты. С применением более совершенной техники микроскопирования, с применением электронной микроскопии, возможно, удастся найти какую-либо «тройственность» в колбочках. Пока же, на основании данных физиологического, функционального изучения цветового зрения,можно лишь предположить, что в сетчатке имеются три сорта колбочек. Предположение, что каждая колбочка содержит в себе красно-, зелено- и синеощущающий аппараты, менее вероятно. Так, если раздражать сетчатку белым раздражителем точечного размера, то при передвиженм его по сетчатке цвет раздражителя будет казаться то красным, то оранжевым, то зеленым, то голубым, то синим, то белым. Такое явление объясняется тем, что белым светом раздражителя возбуждаются колбочки, не одинаково чувствительные к различным излучениям, входящим в белый свет. Светом возбуждаются то красно-ощущающие колбочки, то красно-ощущающие с зеленоощущающими, то только зеленоощущающие, то зеленоощущающие с синеощущающими, то только синеощущающие, то, наконец, колбочки всех трех сортов. В описанных выше электрофизиологических опытах Гранита при перемещении микроэлектрода на сетчатке с одного волокна на другое обычно находились рецепторы с различной чувствительностью по спектру: одни более всего чувствительные к оранжево-красным лучам, другие — к зеленым, третьи — к синим. Этот факт проще всего понимать как результат существования трех видов колбочек с различной чувствительностью.
Допущение трех видов колбочек в сетчатке встречало, однако, возражение, исходящее из фактов, наблюдаемых при сравнении остроты зрения при белом освещении, с остротой зрения при цветном освещении. В последнем случае, казалось, следовало бы ожидать меньшей остроты зрения. Ведь на одной и той Яче площади сетчатки колбочек одного какого-нибудь вида должно быть меньше, чем колбочек всех трех видов. Значит, эти колбочки должны быть расположены друг от друга дальше, чем колбочки всех трех видов. Если применяемое цветное освещение затрагивает лишь колбочки одного вида, то естественно ожидать, что острота зрения в этом случае будет хуже, чем при равноярком белом освещении, возбуждающем колбочки всех трех видов. Опыты не подтверждают, однако, этого вывода. При одинаковой яркости острота зрения оказывается почти одинаковой как в белом, так и в цветном свете (Koenig, 1897; Hartridge, 1947).
Однако можно объяснить этот факт и с точки зрения гипотезы о существовании трех видов колбочек (красно-, зелено - и синеощущающих). Для этого достаточно допустить, что различные виды цветоощущающих колбочек распределяются на сетчатке не в виде правильной мозаики отдельных единичных колбочек различного вида, а в виде неупорядоченной мозаики отдельных групп колбочек различного вида (гипотеза распределения колбочек по сетчатке «пучками»). В последнем случае отдельные колбочки одного вида могут отстоять друг от друга на сетчатке не дальше, чем колбочки разных видов, в силу чего острота зрения в монохроматическом свете может и не снижаться. Необходимо лишь фиксировать рассматриваемый объект «пучком» соответствующих колбочек.
Гипотеза распределения колбочек по сетчатке «пучками» находит себе подкрепление в том, что фиксация центральной ямкой сетчатки очень маленьких цветных пятен осуществляется различными местами центральной ямки соответственно цвету пятна. При фиксации, таким образом, под изображение цветной точки на сетчатке и подставляется «пучок» колбочек соответствующей цветовой чувствительности. Установить это удалось благодаря имевшейся в глазе исследователя микроскотоме на красный цвет. Наличие такой микроскотомы позволяло удерживать глаз в строго фиксированном положении и определять местоположение точек фиксации для разных цветов всегда по отношению к одному и тому же пункту (месту микроскотомы на красный цвет). В табл. 3

и приведены найденные таким путем Хартриджем (1947) удаленности точек фиксации красного , зеленого и синего пятнышек от имевшейся в его глазе микроскотомы на красный цвет. Расстояния указаны в условных единицах.
Найдены были особые места на сетчатке для точек фиксации и некоторых других цветов (оранжевого, желтого, голубого, фиолетового). С точки зрения трехкомпонентной теории это обстоятельство можно объяснить существованием «пучков» на сетчатке, содержащих в себе соединения колбочек двух видов, совместное возбуждение которых и позволяет видеть соответствующий, оранжевый, желтый, голубой или фиолетовый цвета.
В связи с основным ходом рассуждения нам важно здесь лишь отметить, что допущение трех видов колбочек и факты одинаковой остроты зрения при белом и цветном освещении могут быть согласованы друг с другом. Для этого надо лишь принять гипотезу распределения колбочек по сетчатке «пучками». Как мы только что видели, имеются факты, свидетельствующие в пользу такой гипотезы.?
↑ О природе начального процесса возбуждения цветоощущающего аппарата зрительного анализатора
Как же следует представлять себе процесс возбуждения цветоощущающих колбочек светом? Очевидно, световые лучи различных длин волн должны действовать на различные колбочки по-разному. «Красноощущающие» колбочки должны возбуждаться преимущественно длинноволновыми лучами спектра, «зеленоощущающие» — лучами средних длин волн и «синеощущающие» — лучами коротковолновыми.
В сумеречном, бесцветном, зрении начальным процессом зрительного возбуждения является фотохимический процесс, а именно выцветание зрительного пурпура в палочках сетчатки. Относительная яркость различных спектральных лучей для сумеречного зрения хорошо соответствует величинам поглощения этих лучей зрительным пурпуром и скорости его выцветания в этих лучах; нарастание световой чувствительности в темноте довольно хорошо соответствует ходу увеличения концентрации зрительного пурпура и т. д.
Колбочки сетчатки, являясь другой разновидностью концевых аппаратов зрительного нерва, весьма похожи на палочки. Естественно поэтому думать, что и в них возбуждение начинается с фотохимического процесса, возникающего в результате действия лучистой энергии. Чтобы оказать действие, свет должен быть поглощен веществом, реагирующим на освещение. При поглощении света молекулами вещества происходит увеличение их энергии. В результате такого поглощения энергии, происходящего по квантовым законам физики, молекула вещества, поглотившего свет, может перестать быть нейтральной и распасться на электрически заряженные частицы — ионы. При поглощении света зрительным пурпуром образующиеся при распаде его молекул ионы дают начало процессу возбуждения волокон зрительного нерва, связанных с палочковым аппаратом сетчатки (гипотеза П.П. Лазарева).
Но, если в палочковом аппарате наличие светореагирующего вещества в виде зрительного пурпура, или родопсина, уже давно с полной несомненностью установлено, спрашивается, найдено ли какое-либо светореагирующее вещество в сетчаточных колбочках? На вопрос этот в настоящее время следует ответить положительно. Штудниц (Studnitz, 1940) и затем Уолд (Wald, 1937) наблюдали, что сетчатки, почти или полностью лишенные палочек (у некоторых черепах, у кур), все же обнаруживают выцветание при освещении. Выцветание наблюдается даже и от действия крайних красных лучей, не способных вызывать выцветание зрительного пурпура. Блисс (Bliss, 1946) на куриных сетчатках установил более определенным образом и кривую поглощения света светочувствительным веществом (или веществами), содержащимся в сетчаточных колбочках. Поскольку максимум поглощения оказался в желто-зеленоватых лучах спектра (около 560mµ), поглощающее вещество должно иметь фиолетовый цвет. Поэтому его и назвали иодопсином (от греческого слова «ио» — фиолетовый и «эйдес»—вид; иодопсин—-фиолетовидный). На рис. 22
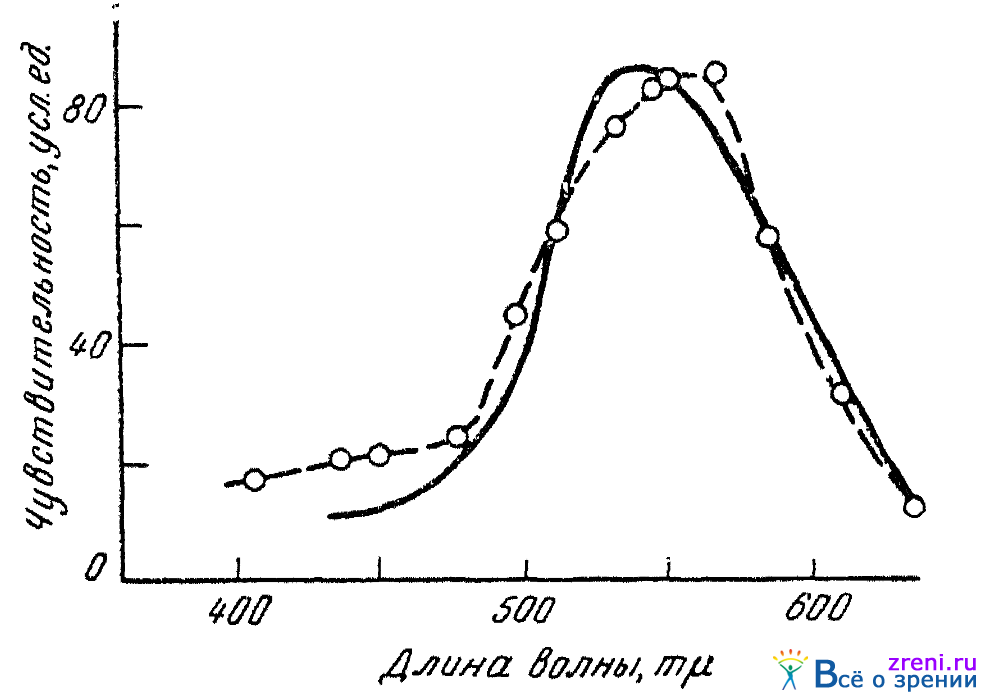
Рис. 22. Кривая поглощения иодопсина (по Блиссу)
штриховой линией показана кривая чувствительности иодопсина к различным монохроматическим лучам спектра, длина волны которых показана на абсциссе. Сплошной линией изображена кривая чувствительности колбочкового зрения человека. По абсциссе отложены длины волн равнокваитового спектра. Как видно, чувствительность (выцветание) имеющегося в колбочках (здесь, правда, в сетчатках кур) вещества обнаруживает хорошее соответствие с чувствительностью колбочкового зрения человека. Подобного рода факты говорят в пользу того, что и цветовое, колбочковое, зрение начинается с процессов фотохимических, процессов изменения молекул некоторого светочувствительного вещества.
Высказывались, однако, предположения, что начальный процесс зрительного возбуждения есть процесс не химический, а физический. Такова, например, гипотеза Вейгерта (Weigert, 1929). Этот автор отмечает, что при освещении очень слабых растворов некоторых светочувствительных веществ наблюдается уподобление цвета раствора цвету раздражающего света. Такое «цветовое уподобление» происходит благодаря пространственным перемещениям внутри взаимносвязанных частиц (так называемых элементарных бинарных абсорбирующих систем) и не является химическим процессом, изменяющим свойства молекул вещества. Слабый раствор зрительного пурпура обладает такой способностью «цветового уподобления». Вейгерт допускает наличие и в колбочках зрительного пурпура, яо лишь в значительно меньшей концентрации, чем в палочках (этим может объясняться и смещение максимума поглощения в более длинноволновую область спектра по сравнению с местом максимального поглощения у пурпура палочек). Однако эта теория едва ли может дать объяснение начальных процессов цветового возбуждения. Как подчеркивает и сам автор, если освещение не очень слабо, никакого явления «цветового уподобления» не наступает. Между тем наша способность различать цвета бывает особенно хорошей как раз при не очень слабом световом раздражении.
Наиболее вероятной поэтому следует считать фотохимическую теорию цветового возбуждения сетчатки.
Продолжение в следующей статье: Современные данные, обосновывающие и уточняющие теорию трехкомпонентности цветового зрения. Часть 3.
---
Статья из книги: Цветовое зрение | Кравков С. В.

Комментариев 0