Как теория трехкомпонентности объясняет основные явления цветового зрения. Часть 4.


Содержание:
Описание
↑ Цветоаномалии
Случаи цветоаномалии (прот- и девтераномалии) с теоретической точки зрения можно объяснять как результаты частично измененной чувствительности того или иного из цветоощущающих аппаратов или сразу нескольких. Такая измененная чувствительность может выражаться либо в количественном изменении чувствительности цветоощущающего аппарата, либо в сдвиге или деформации соответствующей ему кривой основного цветового возбуждения. Следствием последнего являются неправильные, с точки зрения нормального глаза, цветовые уравнения.
Возможно, что в отдельных случаях цветовых аномалий мы имеем дело с деформацией кривой основного цветового возбуждения, захватывающей довольно узкий участок спектра. Поэтому при испытании цветового зрения спектральным аномалоскопом вскрыть какие-либо уклонения от нормы не всегда удается, тогда как при испытании при помощи таблиц уклонения от нормы в зрении этих же лиц обнаруживаются достаточно явно. Объясняется это именно тем, что в таблицах применяются цветные пятна с широкими спектрами отражения, в спектральном же аномалоскопе испытывается нормальность цветоощущающего аппарата обследуемого лица лишь по отношению к избранным трем монохроматическим участкам спектра.
↑ Полная цветовая слепота (ахромазия)
Полная цветовая слепота или ахромазия (ахроматопсия), когда распределение яркости в спектре соответствует сумеречному зрению, объясняется нефункционированием колбочкового аппарата, вызванным его врожденным отсутствием или его повреждением в нашем зрительном анализаторе.
Опыты Гуртового, Семеновской и Шеваревой, выполненные в нашей лаборатории на четырех ахроматах, заставляют признать, что это нарушение аппарата колбочкового зрения у ахроматов может быть различным по своему распространению. Есть основания предполагать, что у одних ахроматов не функционирует колбочковый аппарат на протяжении уже от рецепторов колбочек до выше лежащих центров, у других же поражение колбочкового аппарата коснулось, видимо, лишь высших, мозговых представительств в коре. Более же периферические части колбочкового аппарата (сетчаточные и подкорковые) у этого вида ахроматов оказываются в сохранности.
В нашей лаборатории Гуртовым (1950в) и Семеновской (1950) установлены следующие факты. Некоторые ахроматы различно реагируют по электрочувствительности их глаз на световые раздражители, объективно различающиеся по длине световой волны, но кажущиеся им совершенно одинаковыми, совершенно не отличимыми друг от друга. Так, электрочувствительность в условиях зеленого освещения была у них выше, в условиях красного освещения (при той же для них видимой яркости) — значительно ниже. Приписывать это различие не одинаковой энергии действовавших на глаз зеленых и красных лучей нельзя, ибо энергия красных лучей была больше энергии лучей зеленых, а известно, что большая энергия световых лучей при прочих равных условиях вызывает большую электрочувствительность. У нормальных трихроматов красный и зеленый свет сказывается на электрочувствительности глаз совершенно так же, как у описываемых ахроматов. Остается поэтому допустить, что цветоощущающие аппараты, воспринимающие красные и зеленые раздражители, у описываемых ахроматов отсутствуют лишь в коре, в более же низко лежащих участках зрительного анализатора они сохранены и доносят различные возбуждения до тех подкорковых центров цветового зрения, состояние которых влияет на уровень электрочувствительности глаз.
На рис. 41
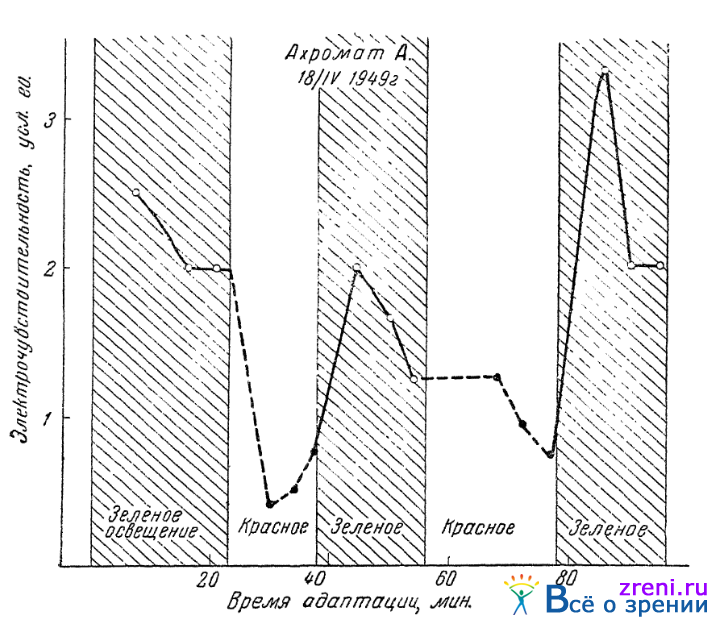
Рис. 41. Влияние цветности на электрочувствительность глаза у ахроматов (по Гуртовому)
показана различная реакция глаз одного из ахроматов на красный и зеленый свет, субъективно для него вполне одинаковый. По данным Гуртового, двое из четырех ахроматов такого явного различия в реакции на красный и зеленый свет не обнаружили. Эти опыты вынуждают допускать упомянутые выше различия, так сказать, в «глубине» ахромазии.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
В подтверждение этого допущения, обследуя тех же ахроматов, Шеварева (1950) нашла, что один из них, в зависимости лишь от цветности освещения, проявляет различную мышечную работоспособность при красном и зеленом свете. У других двух ахроматов Шеваревой не удалось установить какого-либо влияния цветности на мышечную работоспособность или электрочувствительность глаза. Очевидно, у них цветоощущающие приборы не функционируют и в периферической части зрительного анализатора.
Поражения колбочкового цветоощущающего прибора вообще могут иметь место на разных уровнях зрительного анализатора. Когда, например, цвет как таковой не ощущается (случай, описанный Шуманом), мы имеем, очевидно, дело с нарушением центральной, корковой части цветоощущающего аппарата, в то время как более низкие этажи его остаются в сохранности, благодаря чему могут осуществляться еще процессы цветового контраста.
↑ Вопрос об уровнях цветоощущающего прибора
На основании сказанного выше мы вправе заключить, что ряд важнейших фактов нормального и ненормального цветового зрения может быть достаточно удовлетворительно понят с точки зрения трехкомпонентной теории. Это не значит, конечно, что современной теорией уже разрешены все вопросы и что перед нами нет проблем, требующих еще разработки и разъяснения.
Некоторых из неразрешенных проблем мы коснемся. Прежде всего здесь следует указать на недостаточную выясненность, какие уровни зрительного анализатора являются анатомофизиологическим субстратом тех или иных процессов цветового зрения.
Известны факты бинокулярного контраста, когда изменение цвета, видимого одним глазом, вызывается цветом, видимым другим глазом, что свидетельствует о том, что субстрат одновременного цветового контраста может лежать и выше сетчатки. Говоря о явлениях индукции в зрительной области, мы указывали, в связи с данными опытов Селецкой, что, по-видимому, различные виды индуктивных влияний могут осуществляться преимущественно на том или ином уровне зрительного анализатора. Так, положительная индукция не наблюдалась при предъявлении световых раздражителей сетчаткам различных глаз и наблюдалась, когда раздражители действовали на сетчатку того же самого глаза (при равенстве прочих условий).
В только что рассмотренных случаях ахромазии мы пришли к выводу, что цветные раздражители могут влиять на электрочувствительность глаза и при отсутствии корковых представительств колбочковых возбуждений, т. е. в докорковой части зрительного анализатора.
Несомненными представляются в настоящее время и факты бинокулярного смешения цветов, т. е. получение некоторого нового, третьего, цвета в результате раздражения обоих глаз двумя различными цветовыми раздражителями. Опыты бинокулярного смешения цветов проводил еще Ходин (1876). Им были выяснены условия, способствующие наступлению бинокулярного смешения. Это — небольшая и одинаковая яркость смешиваемых цветов. Позже Тренделенбург (Trendelenburg, 1943) провел систематическое исследование бинокулярного смешения различных монохроматических лучей. Он смешивал то красные (671 m?) и зеленые (535 m?) лучи и получал из них оттенки желтого, то различные дополнительные цвета и получал в результате белый. Из этих опытов, как и из опытов других авторов, как будто следовало, что лучи короткой половины спектра при бинокулярном смешении являются относительно более влияющими, чем при смешении монокулярном.
Наблюдалось бинокулярное получение желтого цвета и путем смотрения одним глазом через красный (раттеновский № 29), а другим через зеленый (№ 58) светофильтры. При смотрении же через желтый (раттеновский № 16) и синий (№ 44 А) светофильтры в качестве результирующего удавалось получать ахроматический цвет (Гехт, 1928). Описаны опыты (Hernandez, 1940), в которых у испытуемых был выработан условный кожногальванический рефлекс в ответ на раздражение глаз желтым цветом как условным раздражителем. В качестве безусловного раздражителя применялся электрический ток. Предъявление других цветов электрическим раздражением не подкреплялось и не давало кожногальванического рефлекса. После выработки таким путем условного рефлекса на желтый цвет оказалось, что рефлекс возникал и при одновременном раздражении одного глаза красным цветом, а другого зеленым. Следовательно, такое бинокулярное раздражение вызывало в центрах то же возбуждение, которое вызывается желтым раздражителем.
Учитывая особо яркость смешиваемых лучей для того и другого глаза, Лившиц (1941) нашла, что результаты бинокулярного смешения цветов ничем не отличаются от результатов смешения монокулярного, выводы же Тренделенбурга на этот счет неправильны. Описанные факты, как мы уже говорили выше, свидетельствуют о том, что физиологический субстрат смешения цветов лежит в нашей нервной системе более центрально, чем сетчатка.
↑ О яркостном компоненте колбочковых ощущений
Всякое хроматическое, цветовое, ощущение имеет, как мы видели, три характеристики: цветовой тон, насыщенность и видимую яркость или светлоту. Выше были приведены данные, на основании которых можно признать, что способность получать, хроматические ощущения — цветовое зрение — есть функция колбочкового аппарата сетчатки и связанных с ним нервных центров. В настоящее время накопились факты, заставляющие предполагать, что и в пределах колбочкового зрения следует допускать различные анатомофизиологические субстраты для ощущения яркости цветов и их хроматичности — насыщенности. Если хроматичности без той или иной яркости мы испытывать не можем, то ощущение яркости без цветности, хроматичности, вполне возможно. Яркостная компонента оказывается, таким, образом, чем-то отделимым от компоненты хроматической.
В литературе [Кельнер (Koellner), 1929; Питт (Pitt),.1944] описаны случаи приобретенной полной цветовой слепоты, когда всякое ощущение хроматичности утрачено, а распределение яркости в спектре (кривая видности) соответствует дневному (колбочковому) зрению. Встречаются также случаи, когда в силу тех или иных болезненных процессов выпадает цветоощущение только в половинах сетчаток — так называемая гемиахроматопсия. Места сетчаток, утратившие цветоощущение, сохраняют, однако, колбочковое зрение, если о том судить по остроте зрения. Можно думать, что в этих случаях колбочки дают ощущение яркости, хроматического же ощущения они не способны вызвать.
То же самое, уже в обычных, а не в патологических условиях происходит и при смотрении боковыми местами сетчатки. Если интенсивность цветового раздражителя затрагивает колбочковое зрение, но при этом она не очень значительна, мы видим этот раздражитель периферическими местами сетчатки уже не хроматическим, а серым, большей или меньшей яркости. Относительная яркость различных цветов, видимых периферией, соответствует в этом случае кривой колбочковой, дневной видности.
Заслуживает внимания, что сужение зрачка (зрачковый рефлекс) определяется именно яркостью цветового раздражителя, а не его цветовым тоном или насыщенностью. Величина сужения зрачка для светлоадаптированного глаза под влиянием различных спектральных лучей как раз пропорциональна ординатам кривой дневной видности равноэнергетического спектра [Лауренс (Laurens), 1923].
Мкртычевой и Самсоновой (1944) в лаборатории академика Орбели изучалось влияние кислородного голодания на некоторые зрительные функции. Оказалось, что собственно хроматическая чувствительность колбочкового зрения в условиях кислородного голодания заметно изменяется (для различных цветов в различном направлении) — величина порогов цветоощущения заметно изменяется. Когда же авторы стали определять кривую видности для дневного, фовеального, зрения, то никакого отличия от кривой, получаемой в нормальных условиях (без кислородного голодания), им установить не удалось. При определении же кривой видности, как мы знаем, устанавливаются соотношения между цветами спектра по их яркости. Поэтому результаты работы Мкртычевой и Самсоновой показывают, что действие одного и того же фактора (кислородного голодания) может влиять на хроматическую слагаемую цветового ощущения и не сказываться на его яркостной слагаемой.
Об известной раздельности и самостоятельности яркостного и хроматического компонента в нашем колбочковом зрении свидетельствуют также и следующие факты (Pi?ron, 1942). При оптическом смешении дополнительных цветов яркости смешиваемых цветных раздражителей суммируются, хроматичности же, наоборот, взаимно уничтожаются. Кроме того, хроматическое действие раздражителя в таких случаях совсем не соответствует его яркости; например, синий раздражитель может нейтрализовать цвет гораздо более яркого, дополнительного к нему желтого раздражителя. Возьмем два дополнительных цвета и подравняем их яркость так, чтобы по методу мелькающего фотометра они не давали миганий. Адаптируем глаз к одному из взятых цветов. Оказывается, что это почти одинаково скажется на яркости обоих цветов и их установленное ранее методом мельканий фотометрическое равенство не нарушится. Что же касается хроматичности цветового тона цвета, возникающего от смешения взятых дополнительных цветов, то он, наоборот, резко изменится, после того как мы утомим глаз одним цветом. Отмечаются также различия в ходе нарастания и затухания яркости и хроматичности зрительных ощущении.
Различие в анатомо-физиологическом субстрате для яркостного и хроматического компонентов зрительных ощущении представляется в силу приведенных фактов весьма вероятным. Чего-либо более определенного относительно этого субстрата мы, однако, еще не знаем. Можно лишь высказывать на этот счет гипотезы. На основании данных электрофизиологических экспериментов, описанных выше, выясняется, что светлоадаптированные колбочковые сетчатки имеют в качестве четвертого типа своих рецепторов рецептор, реагирующий на всю область спектра и обнаруживающий максимум чувствительности в зеленовато-желтых лучах, т. е. как раз там, где лежит максимум яркости в спектре для светлоадаптированного глаза. Этот же рецептор (названный Гранитом «доминатором») и можно считать физиологическим аппаратом, соответствующим «яркостному компоненту» наших цветовых ощущений. В связи с этим высказывались предположения (Пьерон, 1945) о существовании в сетчатке четырех видов колбочек. Три и у них содержат в себе особые светореагирующие вещества, чувствительные преимущественно или к красным, или к зеленым, пли к синим лучам спектра: это колбочки красно-, зелено-или синеощущающие. Наряду с ними допускалось существование еще и колбочек «смешанных», содержащих в себе все три светочувствительных вещества и потому возбуждаемых всеми лучами спектра. Предполагается, что эта «смешанная» колбочка и является рецептором, соответствующим яркостном компоненте колбочковых ощущений; допускается, что возбуждение этой «яркостной» колбочки всегда в равной мере возбуждает и три «хроматические» колбочки, что также вызывает у нас ощущение яркости. Схема подобной сетчаточной колбочковой "четверки". по Пьерону, приводится на рис. 42.
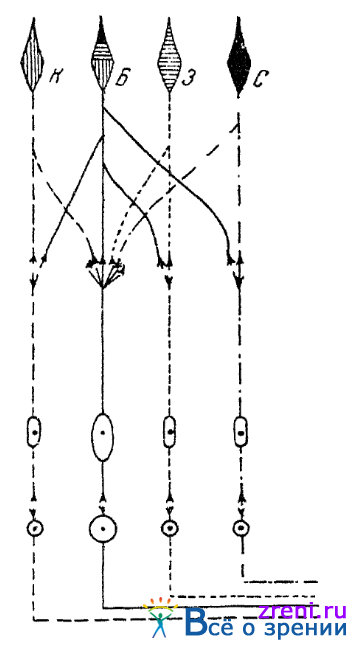
Рис. 42. Колбочковая «четверка» (по Пьерону)
↑ О физиологической природе ощущений от желтого цвета
Не вполне выясненным является и вопрос о физиологической природе ощущений от желтого цвета. С точки зрения трехкомпонентной теории физиологическим субстратом ощущения желтого является определенное соотношение возбуждений красно- и зелено-ощущающих аппаратов зрительного анализатора. Предполагается, что особого, желтоощущающего аппарата, который был бы четвертым рецептором, в нашем зрительном анализаторе не существует.
Приходится, однако, встречаться с фактами, которые, по мнению некоторых, говорят в пользу существования для желтого особого цветоощущающего аппарата. Так, наблюдалось, что чувствительность к синему возрастает после предварительного раздражения глаз желтым светом; если же предварительно раздражать глаз попеременно красным и зеленым, то после этого сенсибилизации к синему не наблюдается (Шварц, 1948). Хартриджем (Hartridge, 1947) описаны опыты, касающиеся видности цветового тона раздражителей весьма малого углового размера. Согласно этим опытам, желтая точка видна как желтая с большего расстояния, если вблизи нее имеется другая такая же желтая точка. Подобное же «содействующее» влияние, хотя и меньшее, наблюдается и тогда, когда эта вторая, «индуцирующая», точка зеленовато- или же красновато-желтая. Если же в качестве индуцирующих брались точки красные или зеленые, никакого «содействия» не наблюдалось, угловой порог видности «реагирующей» желтой точки оставался без изменения.
Эти и подобные им факты, как мы полагаем, говорят не о существовании особого рецептора для желтого цвета, а об особом действии того возбуждения, которое соответствует в коре головного мозга желтому цвету.
Что дело обстоит именно так, свидетельствуют и результаты недавних опытов, проведенных в нашей лаборатории Селецкой и Гуртовым (1950). Они наблюдали, что точечное раздражение желтого цвета индуктивного влияния не оказывает, в то время как входящие в это раздражение красные лучи, отдельно взятые, производят определенный индуктивный эффект.
↑ Особенности цветоощущения центра сетчатки
Ждет своего объяснения с эволюционно-биологической точки зрения и своеобразие цветовой чувствительности самой центральной части центральной ямки сетчатки. На особенности цветоощущения этой области нашего зрительного прибора обращено вновь внимание исследователей, хотя первые наблюдения на этот счет уже далеко не новы и восходят к Кенигу (1894), который на основании опытов над самим собой описал своеобразие цветовой чувствительности центральной ямки сетчатки. Согласно его наблюдениям, центральная ямка лишена цветовой чувствительности к синим раздражителям. Так, синяя точка слабой яркости для строго фовеального зрения не отличима от точки зеленого цвета. Если же посмотреть несколько боком, парафовеальными местами сетчатки, то синий цвет того же раздражителя становится хорошо видимым. Размер площади сетчатки, на которой не ощущается синий цвет, соответствует, по Кенигу, 55—70 угловым минутам. В этой области диск луны может делаться невидимым, если смотреть на него через синее стекло, в то время как другими местами сетчатки через то же стекло он видим. Все цвета спектра могут этой «синеслепой» частью сетчатки подравниваться смесям всего двух спектральных лучей (650 и 475 m?).
Не так давно аналогичные явления были описаны Уилмером и Райтом (Wright, 1946). В объектах небольшой яркости, видимых центральной ямкой сетчатки под малыми углами зрения (около 20 угловых минут, чему соответствует приблизительно объект диаметром в 6 мм с расстояния в 1 м), глаз не отличает синего от зеленого и красного от пурпурного. Если же на подобные пары цветов посмотреть с того же расстояния другими местами сетчатки, сдвинув взор на 40—50 минут в сторону, то указанных смешений цветов уже не наблюдается, хотя изображение и находится все еще в области желтого пятна. Неразличение цветов в первом случае следует поэтому объяснять не пигментацией области желтого пятна, а физиологическими особенностями цветоощущения центральной ямки сетчатки.
На специальном колориметре для поля зрения, размером всего в 15—20 угловых минут, Райт нашел также возможным все цвета спектра подравнивать смесям всего двух лучей. Для этого брались лучи синие (460 m?) и красные (650 m?). Результаты определений (по данным опытов Ранта и Томсона; Wright, 1946) приведены на рис. 43.
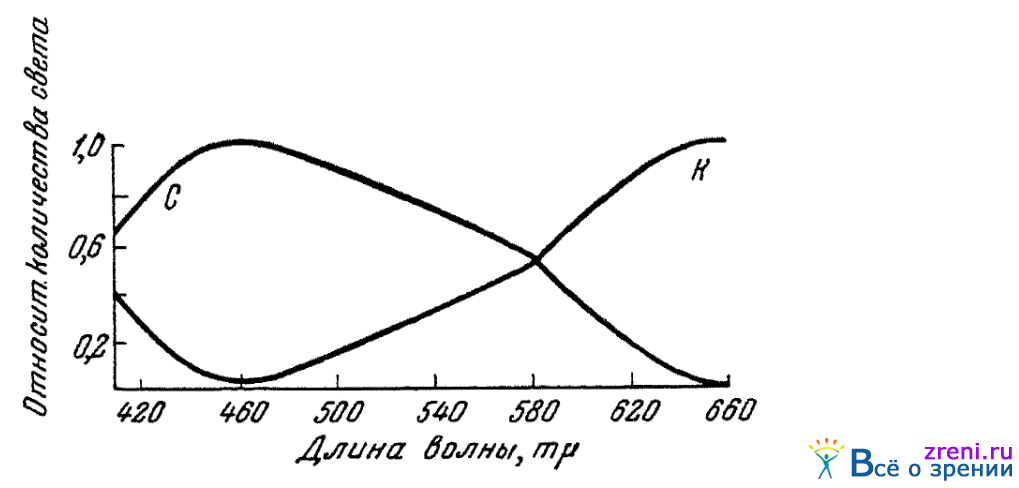
Рис. 43. Кривые смешения основных раздражителей для центра фовеальной части сетчатки (по Райту и Томсону)
По ординате показаны относительные количества света длиной волны в 460 m? (С) и 650 m? (К), которые нужно было брать, чтобы для строго фовеального зрения устанавливать цветовые равенства с цветами, длины волн которых отложены по абсциссе. Интенсивности излучения 460 и 650 m?, уравниваемые с цветом 582 m?, приняты условно за равные.
Указанные авторы на основании всех этих наблюдений находят возможным говорить о слепоте центральной ямки сетчатки на синий цвет, поскольку ощущение синего в ней не возникает и все многообразие цветов может быть получено оптическим смешением всего двух раздражителей. Правда, они не считают все же эту слепоту полной, по ним центральная ямка обладает лишь резко пониженной чувствительностью к синим лучам.
Вывод этот находит подтверждение и в опытах Раутина и Демкиной (1948), выполненных в Гос. Оптическом институте в Ленинграде. Авторы определяли зоны ошибок, допустимых при колориметрировании разных цветов. Они определяли, как можно изменять количества основных раздражителей, воспроизводящих данный цвет, без того чтобы цвет показался нам уже изменившимся. Оказалось, что при уменьшении угловой величины поля все большей делается ошибка для синего слагающего. Иными словами, когда площадь раздражителя приближается к размеру центральной ямки сетчатки, участие синего возбуждения делается все меньшим. Поэтому и оказывается возможным, не изменяя цветового ощущения, все более значительно изменять долю синего раздражителя.
---
Статья из книги: Цветовое зрение | Кравков С. В.

Комментариев 0