Роговая оболочка │ Часть 1


Описание
Фиброзная (наружная) оболочка глазного яблока (tunica fibrosa bulbi) состоит из роговой оболочки (cornea) и склеры. Развивается этот слой из эктомезенхимы, окружающей глазной бокал во время эмбрионального развития.Роговица представляет собой прозрачную часть фиброзной оболочки, составляющую 1/6 площади поверхности глаза (1,3 см2) и имеющую больший радиус кривизны, чем склера (рис. 3.2.1).

Рис. 3.2.1. Вертикальный и горизонтальный размеры передней и задней поверхностей роговой оболочки (а), радиус кривизны роговой оболочки и склеры (б) и ее толщина в центральных участках и по периферии (в) (по Hogan et al. 1971)
Различают гистологическую и хирургическую границу роговицы. Гистологической границей является линия на внутренней поверхности роговицы, отделяющая прозрачную часть роговицы от непрозрачной склеры.
Хирургической границей считается линия, идущая от места прерывания передней пограничной пластинки (боуменовой оболочки) к месту прерывания задней пограничной пластинки (lamina limitans postrior sclererae; Decemett).
Спереди роговая оболочка овальной формы. Горизонтальный диаметр передней поверхности равен 11,7 мм, а вертикальный—10,6 мм (рис. 3.2.1). У мужчин диаметр роговицы приблизительно на 0,1 мм больше. У детей он меньше — 10 мм. Задняя поверхность роговицы имеет вид окружности (диаметром 11,7 мм).
Толщина роговицы в центре равна 0,52 мм, а по периферии — 0,67 мм. У новорожденных ее толщина больше, чем у детей первого года жизни, что связывают со становлением в этот период времени функции эндотелиальных клеток.
Несколько более подробно анатомические, физические и оптические свойства роговицы приведены в табл. 3.2.1.

Таблица 3.2.1. Размеры, оптические и физические свойства роговой оболочки
Центрально расположенная зона роговой оболочки, диаметром 4 мм, называется оптической зоной. Она почти сферичная. Радиус кривизны передней поверхности в оптической зоне равен 7,8 мм, а задней — 6,5 мм. Рефракционная сила в этой области равняется 43 дптр. У индивидуумов с астигматизмом оптическая зона может быть несколько эллипсоидной формы. К периферии роговица несколько уплощается, что придает ей форму гиперболоида Уплощение более выражено с назальной стороны и снизу.
Кривизна роговицы изменятся с возрастом. У новорожденных она более сферичная и уплощается к 5-летнему возрасту. При этом изменяются диаметр и площадь роговицы (табл. 3.2.2, 3.2.3).

Таблица 3.2.2. Диаметр, радиус кривизны и площадь роговицы в детском возрасте

Таблица 3.2.3. Сравнительные размеры роговой оболочки новорожденного и взрослого
В несколько более позднем возрасте развивается «правильный» астигматизм, заключающийся в том, что в вертикальном меридиане радиус кривизны роговицы меньше. В связи с этим роговая оболочка в вертикальном меридиане обладает более сильной рефракционной способностью. Роговица становится сферичной в среднем возрасте. При этом развивается «неправильный» астигматизм.
Сферичность и гладкость передней поверхности роговицы являются важными факторами, обеспечивающими ее прозрачность. При нарушении сферичности развивается астигматизм и существенно снижается зрение. Наиболее ярко это проявляется при кератоконусе. В тех случаях, когда формируется рубцовая ткань роговицы, но сохраняется ее кривизна, острота зрения страдает в меньшей степени.
Роговая оболочка постепенно переходит в непрозрачную склеру. Место перехода роговой оболочки в склеру называется лимбом. Именно в этой переходной зоне определяются довольно существенные структурные изменения роговицы (рис. 3.2.6).

Рис. 3.2.6. Область лимба. Переход переднего эпителия роговой оболочки в эпителий конъюнктивы глазного яблока: 1 — задний эпителий роговой оболочки (эндотелий); 2 — строма роговой оболочки; 3 — передний эпителий роговой оболочки; 4 эпителий конъюнктивы глазного яблока; 5 - субэпителиальная соединительная ткань конъюнктивы; 6—кровеносные сосуды; 7 —трабекулярная сеть; 8 — шлеммов канал. Отмечается изменение строения эпителиального пласта и появление обильной субэпителиальной ткани, содержащей большое количество кровеносных сосудов
Традиционно роговую оболочку разделяют на пять слоев —
- передний эпителий роговицы (epithelium anterius соrпеае),
- передняя пограничная (боуменова) пластинка (lamina limitans anerior; Bowman),
- собственное вещество роговицы (substantia propria соrпеае),
- задняя пограничная (десцеметова) пластинка (lamina limitans postrior соrпеае; Decemett)
- задний эпителий роговицы (эндотелий) (epithelium posterius соrпеае) (рис. 3.2.2, 3.2.3).
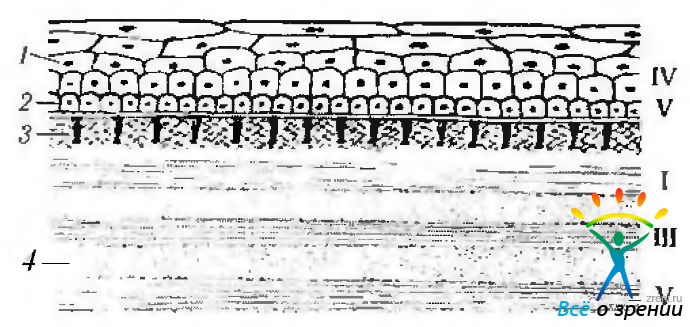
Рис. 3.2.2. Схематическое изображение строения роговой оболочки и распределения в ней различных типов коллагена (по Bron et al., 1997): 1 — эпителий; 2 — базальная мембрана; 3 — боуменов слой; 4 — строма; 5 — десцеметова мембрана; 6 - эндотелий

Рис. 3.2.3. Микроскопическое строение центральных участков роговой оболочки в полную ее толщину (а), а также при большем увеличении ее передних (б) и задних слоев (в): 1 — передний эпителий; 2 — боуменова оболочка; 3— строма; 4— десцеметова оболочка; 5 — задний эпителий (эндотелий)
Ряд авторов приводят и еще один слой — слезную пленку, имеющую большое физиологическое значение, но в гистологическом смысле не являющуюся структурным компонентом роговицы. В связи с важностью этого образования мы начнем изложение строения роговой оболочки именно с нее.
Слезная пленка. Для роговицы, выполняющей функцию линзы, граница между воздухом и передней поверхностью роговицы, на у ровне которой и реализуется преломляющая сила глаза, должна быть высококачественной оптической поверхностью. Качественную оптическую поверхность и обеспечивает слезная пленка. Другими функциями слезной пленки является смачивание конъюнктивы век во время мигания и антибактериальное действие, благодаря наличию в слезе лизоцима и бета-лизина.
Коэффициент преломления слезной пленки равняется 1,357, а объем — 7 мкл. Скорость обмена равна 0,5—2,2 мм3 мин.. Толщина слезной пленки колеблется от 6 до 20 мкм (в среднем 7 мкм).
Состоит она из трех слоев:
- наружный липидный, толщиной 0,1 мкм,
- средний водянистый слой, толщиной 7 мкм,
- внутренний слизистый слой, толщиной 0,02—0,05 мкм.
В состав липидного слоя входят стеариновые и холестериновые эфиры, находящиеся при температуре тела в жидком состоянии. Основной функцией липидного слоя является уменьшение испарения слезы. Главным источником липидов являются мейбомиевы железы и, в меньшей степени, железы Цейса и Молля.
Водянистый слой имеет наибольшую толщину и состоит из водных растворов неорганических солей, глюкозы, мочевины, ферментов, белков и протеогликанов. Компоненты водянистого слоя секретируются главной и добавочными слезными железами. Добавочные слезные железы в количестве 4—35 расположены в нижнем своде конъюнктивальной полости. Весят они от 0,3 до 7,0 мг, что составляет 10% от массы основной части слезной железы. Вследствие того, что между миганиями слезная пленка становится гиперосмотической, некоторые компоненты водянистого слоя могут путем осмоса поступать в водянистую влагу через роговицу.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Слизистый слой, лежащий под водянистым слоем, является частью поверхностного эпителия роговицы. Его толщина всего несколько сотых микрона, и он покрывает микроворсины эпителиальных клеток. Слизь вырабатывается бокаловидными клетками конъюнктивы и распределяется по поверхности роговицы и конъюнктивы благодаря мигательным движениям век. Часть растворимого муцина выделяют главные железы.
Формирование слезной пленки и поддержание ее структуры обеспечивается функцией век. При каждом мигании равномерно распределяется по поверхности глазного яблока муцин, а также водянистая и липидная части секрета. Сразу после образования пленки начинается и ее испарение.
Высокое поверхностное натяжение обычно сохраняется на протяжении одной минуты. Затем слезная пленка дестабилизируется, разрушается, и на передней поверхности роговицы образуется так называемые сухие пятна. С каждым новым миганием поверхность роговицы снова покрывается пленкой.
Промежуток времени между миганием и появлением сухих пятен называется временем распада слезной пленки. В норме это время составляет 15—34 секунды. Время распада менее 10 секунд свидетельствует о наличии патологического процесса слезной железы, желез пальпебральной и бульбарной конъюнктивы.
Увеличение количества липидов в составе слезной пленки или загрязнение конъюнктивальной полости могут быть причиной укорочения времени распада, что, в свою очередь, приводит к развитию симптома сухо го глаза.
Передний эпителий (epithelium anteriiis). Передний эпителий роговой оболочки в соответствии с гистологической номенклатурой, относится к многослойным плоским неороговевающим эпителиям, т. е. аналогичен эпителиальной выстилке пищевода, слизистым полости рта, надгортанника, влагалища и др.
Прозрачность эпителия зависит от однородности коэффициента преломления светового луча клеточным слоем. При отсутствии патологических изменений роговицы передний эпителий не виден при использовании щелевой лампы. Возникновение межклеточного отека приводит к тому, что эпителиальный пласт утрачивает свою однородность и становится видимым.
Толщина переднего эпителия роговой оболочки равняется 50,7 мкм. Состоит он из 5—6 покрывающих друг друга клеточных слоев (рис. 3.2.2, 3.2.3).
Клетки наиболее поверхностного слоя имеют плоскую форму, в связи с чем эпителий и получил свое название. Длина плоских клеток равна 45 мкм, а толщина — 4 мкм. Эти клетки имеют самую большую площадь, увеличивающуюся по направлению к периферии роговицы (850 мкм2 на периферии и 560 мкм2 в центре) (рис. 3.2.3—3.2.5).
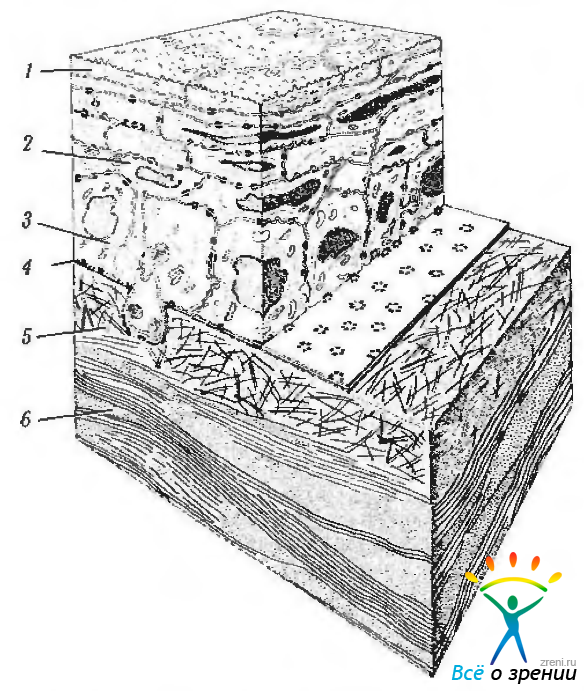
Рис. 3.2.4. Схематическое изображение светооптической и ультрастру ктурной организации передних отделов роговой оболочки (по Pouliquen, 1969): 1 — поверхностные эпителиальные клетки; 2 — эпителиальные клетки средних слоев; 3 — базальные клетки эпителия; 4 — базальная мембрана: 5 — боуменова оболочка; 6 — передние слои стромы
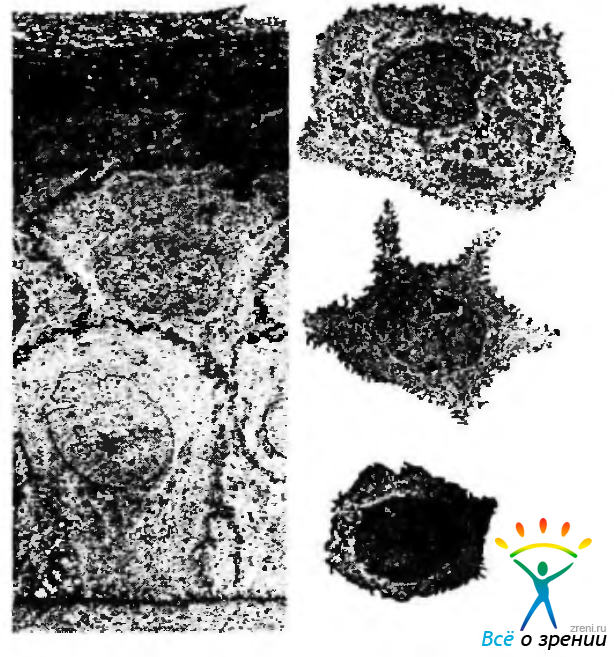
Рис. 3.2.5. Особенности ультраструктурной организации эпителиоцитов различных слоев переднего эпителия роговой оболочки (по Hogan et al., 1971): а — электроннограмма среза переднего эпителия роговой оболочки; б — электроннограммы изолированных клеток различных слоев роговицы
Между эпителиоцитами определяется большое количество десмосом. Запирательные пластинки расположены на апикальной поверхности клеток, т. е. поверхности, примыкающей к прекорнеальной слезной пленке. Эти органоиды рассматриваются большинством авторов как структуры, определяющие прозрачность роговой оболочки, обеспечивая мощное препятствие на пути распространения воды, электролитов и глюкозы в строме роговицы.
Обращенная кнаружи клеточная поверхность эпителиальных клеток образует большое количество микроворсин высотой 1—2 мкм и микроскладок, покрытых гликокаликсом. Слой гликокаликса, толщиной 300 нм, сохраняется после гистологической обработки. Состоит он из гликопротеидов и многочисленных микрофиламентов, длиной 150 нм. Микрофиламенты прикрепляются к цитоплазматической мембране клеток. Необходимо отметить, что в гликокаликсе, покрывающем конъюнктивальный эпителий, микрофиламенты значительно длиннее и достигают 300 нм.
Основной функцией микроворсин является стабилизация слезной пленки на поверхности роговицы. Среди поверхностно расположенных эпителиоцитов выявлены «светлые» и «темные» клетки, отличающиеся количеством микроворсинок. По мнению ряда авторов, «темные» клетки являются более старыми и в ближайшее время будут «слущены».
Цитоплазма эпителиоцитов поверхностных слоев насыщена органоидами (тонофилламенты, свободные рибосомы, шероховатый эндоплазматический ретикулум, аппарат Гольджи). Митохондрии, как правило, небольшого размера и встречаются нечасто. Это свидетельствует о низком уровне аэробного окисления и большей зависимости дыхания клеток от пентозного пути метаболизма. Часто встречаются центриоли. В цитоплазме можно также обнаружить включения гликогена в виде мелкодисперсных гранул, размерами 20—30 нм. Количество зерен гликогена заметно уменьшается при гипоксии эпителиоцитов (ношение контактных линз) и при регенерации клеток в посттравматическом периоде. В поверхностных клетках переднего эпителия видны многочисленные пузырьки. связанные с аппаратом Гольджи.
Средний (промежуточный, переходный) слой переднего эпителия складывается из 2 или 3 слоев клеток крыловидной и зонтикоподобной формы (рис. 3.2.3—3.2.5). Диаметр клеток — приблизительно 12—15 мкм. Ядра этих клеток, как и поверхностных своей длинной осью ориентированы параллельно поверхности роговицы. Их цитоплазматические отростки проникают между телами соседних клеток. Цитоплазма насыщена органоидами. Соединены клетки многочисленными десмосомами. Появляются в них тонофиламенты, длиной 8 нм.
Базальный слой представляет собой один слой высоких полигональных клеток, размерами 18x10 мкм. Ядра клеток базального слоя имеют диаметр 5,7 мкм и смещены в апикальную часть клеток. В этом клеточном слое определяются митотические деления. Именно по этой причине этот слой клеток называют еще герминативным. Один митоз встречается на 250 клеток. Значительно большее число митозов определяется среди клеток базального слоя, по периферии роговой оболочки.
При митотическом делении базальных клеток эпителия дочерние клетки перемещаются кпереди в слой крыловидных клеток. При этом клетки сохраняют свою полигональную формул, но становятся тоньше. Ядра уплощаются и ориентируются параллельно поверхности клетки. Число внутрицитоплазматичсских органоидов заметно уменьшается. При этом увеличивается количество межклеточных контактов (десмосом и запирающих пластинок). К базальной мембране эпителиальные клетки базального слоя присоединяются при помощи полудесмосом.
Дифференциация клеток переднего эпителия по слоям и пролиферативная активность клеток базального слоя довольно существенно изменяются с возрастом и под влиянием различных патологических факторов. Подтверждением тому являются как клинические наблюдения скорости регенерации переднего эпителия у пожилых людей, так и экспериментальные исследования при моделировании процессов старения организма в целом и эпителия роговицы в частности.
В базальном слое переднего эпителия можно обнаружить клетки неэпителиального происхождения. В первую очередь к таковым необходимо отнести дендритические клетки. Различают два типа клеток дендритической формы. Первый тип предположительно относится к меланоцитам, а второй — к так называемым клеткам Ларгенганса. Клетки Ларгенганса несут функцию иммунокомпетентных клеток. Именно они распознают чужеродный антиген и передают полученную информацию лимфоцитам. Эти клетки появляются в строме роговой оболочки довольно рано. С возрастом их количество уменьшается, и остаются они лишь по периферии роговицы! При воспалении роговицы клетки Ларгенганса появляются в центральных участках. В базальном слое довольно часто можно увидеть и лимфоциты и макрофаги.
Передний эпителий роговой оболочки к периферии в лимбальной области постепенно переходит в эпителий бульбарной конъюнктивы. Среди эпителиоцитов появляются бокаловидные клетки. изменяется характер подлежащей стромы. Базальная мембрана (рис. 3.2.4, 3.2.5). Базальная мембрана переднего эпителия окрашивается при проведении ШИК-реакции в розовый цвет (РAS-положительна). Толщина ее колеблется от 75 до 100 нм.
Базальная мембрана формируется благодаря синтетической деятельности базальных клеток эпителия. Эти клетки образуют и полудесмосомы. Через полудесмосомы вдоль мембран базальных клеток и через базальную мембрану проникают филаменты, обеспечивающие прочное сцепление эпителиальных клеток и мембраны. Часть фибрилл оканчивается на фибриллах коллагена I типа.
Базальная мембрана состоит из двух структурных компонентов — гранулярного и волокнистого. Глубокий слой осмиофилен и имеет толщину 30—60 нм. Называют этот слой lamina densa (темная пластинка). Толщина поверхностного слоя (lamina lucida) — 24 нм. Lamina lucida базальной мембраны представляет собой аморфную пластинку, спаянную с телом полудесмосомы. Эту зону пересекают «якорные» филаменты, которые затем проникают в lamina densa базальной мембраны и заканчиваются в боуменовой оболочке. Перечисленные структуры состоят из коллагена VII типа. Иммуноморфологически выявлены и особенности химической организации базальной мембраны. Так, lamina lucida состоит из гликопротеида ламинина и буллезного пемфикоидного антигена. Lamina densa состоит из коллагена IV типа. В базальной мембране обнаружен также фибронектин.
Плотный контакт между базальной мембраной и боуменовой оболочкой нарушается при обработке роговой оболочки детергентами, при воспалительных, дистрофических заболеваниях, отеке и диабете. При этом плотный контакт сохраняется между базальной мембраной и эпителиальными клетками.
При повреждении базальной мембраны развивается состояние, характеризующееся появлением рецидивирующих эрозий эпителия.
Базальная мембрана разрушается протеолитическими ферментами (трипсин, хемотрипсин). По мере старения организма она утолщается и становится многослойной.
Базальная мембрана толще по периферии роговой оболочки. Утолщается она при диабете и после травмы. Базальная мембрана сращена с боуменовой оболочкой.
Боуменова оболочка (передняя пограничная пластинка; lamina limitans anerior; Bowman) расположена под эпителием (рис. 3.2.2—3.2.4). Толщина ее составляет 8—14 мкм, и обнаруживается она при микроскопическом исследовании только у приматов, части птиц и рептилий, а также у рыб. Ее отсутствие у низших животных приводит к изменению эластичности роговой оболочки. По этой причине при определении внутриглазного давления у животных необходимо проводить калибровку инструментов.
При световой микроскопии боуменова оболочка выглядит гомогенной бесклеточной пластинкой, в связи с чем ее раньше называли мембраной. Тем не менее боуменова оболочка не имеет строения, характерного для мембранных образований. Поэтому более правильно назвать ее «слой» или «оболочка». Фактически, боуменова оболочка представляет собой так называемый модифицированный, т. е. видоизмененный, слой стромы роговицы.
При нормальном или повышенном внутриглазном давлении боуменова оболочка кажется гладкой. Тем не менее при падении внутриглазного давления, проведении аппланационной тонометрии, хирургических вмешательствах, а также при наложении давящей повязки на веки в боуменовой оболочке можно обнаружить многочисленные гребни (складки). Возникают они и при массаже роговицы через веко. Появление этих гребней связывают с изменением ориентации ремнеподобных «стромальных связок». Дегенеративные изменения гребней, что бывает при длительной гипотонии или атрофии глазного яблока, приводят к возникновению «шагреневой» поверхности роговицы.
Ультраструктурно боуменова оболочка состоит из беспорядочно распределенных и плотно упакованных коллагеновых фибрилл диаметром 14—27 нм и длиной 240—270 нм. Периодичность поперечной исчерченности волокон равняется 64 нм. Основное вещество роговой оболочки имеет такой же состав, как и основное вещество стромы. Оболочка Боумена состоит из коллагена I типа, основного структурного компонента роговицы и склеры, а также коллагенов V, VI, III и VII типов. Ряд исследователей выявили коллаген IV типа. Передняя поверхность боуменовой оболочки, граничащая с lamina vitrea базальной мембраны эпителиальных клеток, гладкая, а задняя поверхность — неровная. При растровой микроскопии она выглядит волнистой и содержит поры диаметром 0,5—1,5 мкм. Через эти поры к эпителиальным клеткам проникают немиелинизированные нервные волокна.
Боуменова оболочка устойчива к повреждению и довольно длительно сохраняется при воспалении. Если же она разрушена, регенерации не наступает и это место замещается волокнистой тканью.
В норме боуменова оболочка не содержит клеток. Первым признаком развития патологического состояния роговой оболочки является появление в этой зоне клеток. Правда, необходимо отметить, что через поры в боуменовой оболочке и в норме возможна миграция к эпителиоцитам и клеток иного происхождения.
Собственное вещество (строма) роговицы (substantia propria соrпеае). Строма составляет 90% толщины роговой оболочки (450 мкм в центральных участках) и складывается из трех компонентов: коллагеновых пластин, клеток и основного вещества (рис. 3.2.3, 3.2.4). В соответствии с гистологической номенклатурой строма представляет собой плотную оформленную соединительную ткань.
Существует две теории, объясняющие прозрачность стромы роговицы. Первая предложена Maurice и сводится к предположению о том. что роговичные коллагеновые волокна формируют решетчатую структуру, уменьшающую светорассеивание в результате общей интерференции от каждой фибриллы. До тех пор, пока фибриллы расположены в решетке равномерно и промежуток между ними меньше длины волны видимого света (400—700 нм), роговица остается прозрачной. Когда же расстояние между фибриллами увеличивается, общая интерференция уже не имеет места и роговица мутнеет. Goldman, Benedek утверждают, что роговица прозрачная вследствие того, что фибриллы довольно малы по отношению к длине волны света и не преломляют свет при прохождении через них до тех пор, пока они не больше половины длины волны света.
В настоящее время прозрачность стромы роговой оболочки связывают с рядом структурных ее особенностей и химическим составом. Помимо вышеприведенных причин возможной прозрачности стромы роговицы, приводят и ряд других причин. Прежде всего, определенное значение имеет исключительно строгая ориентация коллагеновых пластин, что показано при помощи метода дифракции.
Имеет также значение определенное соотношение между коллагеном и матричными белками (протеогликанами). Нарушение этого взаимоотношения приводит к помутнению роговицы.
Необходимо отметить, что факт быстрого обратимого помутнения роговицы, которое имеет место при повышении, а затем снижении внутриглазного давления, очень сложно объяснить с позиций у казанных двух теорий. Поэтому вопрос о причинах прозрачности стромы до сих пор остается открытым.
Стромальные пластины. Каждая стромальная пластина состоит из пучка коллагеновых волокон, ориентированных параллельно друг другу (рис. 3.2.7).
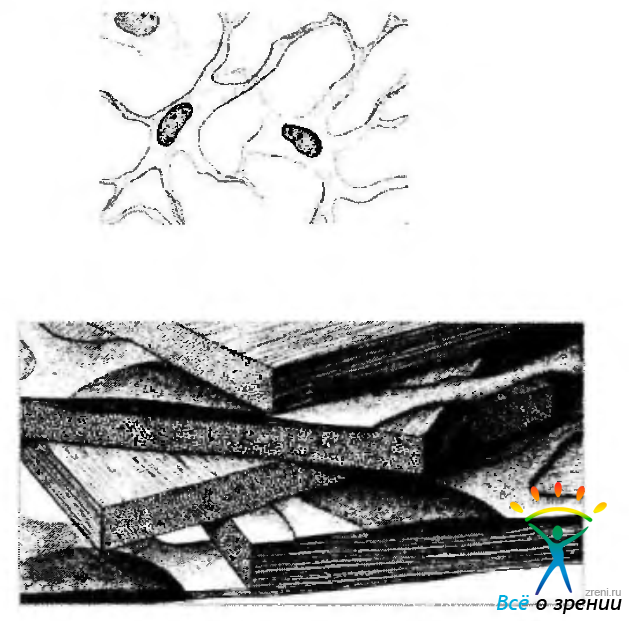
Рис. 3.2.7. Схема микроскопической организации стромы роговой оболочки (по Hogan et al., 1971): а — синтициальное расположение кератоцитов; б — расположение и структурная организация стромальных пластин
Фибриллы обладают типичной исчерченностью, равной 64 нм и характерной для коллагановых волокон других типов соединительной ткани. Коллагеновые волокна состоят, в основном, из коллагена I типа, хотя выявлен и коллаген III, VI и XII типов.
Отмечается уникальная однородность диаметра фибрилл, хотя и выявляется небольшое увеличение их диаметра в зависимости от глубины стромальной пластины. Фибриллы поверхностных слоев имеют диаметр 27 нм, а задних — 35 нм. Некоторые авторы не нашли подобных различий. Выявлено, что диаметр фибрилл передних и задних стромальных пластин одинаков и равен 22,0 ±1,0 нм. Расстояние между фибриллами также примерно одинаковое: 43,2+1,7 нм — в передних слоях и 45.6 нм — вблизи десцеметовой оболочки. Расстояние между фибриллами с возрастом уменьшается.
Коллагеновые фибриллы складываются в пластины, направление которых зависит от глубины слоя роговицы. Толщина одной пластины колеблется от 1,5 до 2,5 мкм, а ширина от 9 до 260 мкм. Число коллагеновых пластин равняется 300 в центральных участках роговицы и увеличивается до 500 по периферии.
Стромальные пластины задних отделов роговой оболочки, распространяются циркулярно вдоль лимба, формируй «циркулярную связку». В то же время стромальные пластины передних слоев располагаются параллельно друг другу и параллельно поверхности роговицы.
В центральных участках пластины перекрещиваются под различным утлом в горизонтальной плоскости. В поверхностных слоях роговицы пластины переплетаются примерно таким же образом, как в плетеных бамбуковых креслах. По периферии они раздваиваются, делятся на три части и перемешиваются с циркулярной коллагеновой пластинкой лимба (рис. 3.2.8).

Рис. 3.2.8. Особенности расположения и взаимоотношения коллагеновых пластин роговой оболочки и склеры. Обращает на себя внимание различный диаметр коллагеновых волокон, расположенных в склере (по Bron et аl., 1997)
Приведенное расположение стромальных пластин передних слоев роговицы приводит к формированию так называемой мозаики. Эту мозаику можно наблюдать, проведя следующие действия. Первоначально закапывают в конъюнктивальную полость флюоресцеин, нажимают на глазное яблоко пальцем. После открытия век четко видно распределение флюоресцеина в виде многоугольников. Подобное распределение флюоресцеина и отражает особенности архитектоники распределения коллагеновых пластин передних слоев стромы.
Параллельное расположение пластин передних отделов стромы роговицы и сохранение подобного расположения на границе с задними слоями позволяют производить межпластинчатое расслоение роговой оболочки при кератопластике.
Необходимо отметить, что передние и задние слои стромы отличаются как строением, так и физико-химическими свойствами. Так, задние слои стромы более упорядочены, более гидратированы, обладают более низким преломляющим индексом. Кроме того, коллагеновые пластины задних слоев стромы шире и толще (100—200 мкм— ширина и 1,0—2,5 мкм — толщина) передних слоев (0,5—30 мкм — ширина и 0,2—1,2 мкм — толщина). Имеются также и определенные различия строения кератоцитов.
Существование структурных различий передних и задних слоев стромы роговицы многие авторы рассматривают как основу большей устойчивости передних слоев к отеку. Именно это свойство обеспечивает сохранение кривизны роговицы и ее прозрачность при различных физиологических и патологических состояниях.
Стромальные пластины погружены в основное вещество, представленное различными типами протеогликанов. Гидрофильная часть основного вещества гликозаминогликанов, в которую погружены коллагеновые волокна, приобретает форму протеогликанов путем ковалентного соединения гликозаминогликанов с белками. Протеогликаны имеют довольно разнообразное химическое строение. В строме роговой оболочки из гликозаминогликанов обнаружены кератан сульфат, хондроитин-4-сульфат, ходроитии-6-сульфат, дерматан сульфат.
Молекулы гликозаминогликанов окутывают волокна и ориентируются перпендикулярно кол-лагеновому волокну. Именно связь между волокнами и протеогликанами опеспечивает прозрачность роговичной ткани.
Различные типы гликозаминогликанов в роговой оболочке распределены неравномерно. Некоторые из них преобладают в передних слоях стромы, другие — в задних слоях. С преобладанием того или иного типа гликозаминогликанов в различных слоях стромы связана различная степень гидратации стромы, с которой частично связана прозрачность стромы. Нарушение синтеза гликозаминогликанов (врожденное или приобретенное) приводит к помутнению роговицы, связанному с отложением продуктов патологического синтеза.
Клетки стромы (кератоциты). Основным клеточным элементом стромы роговой оболочки является кератоцит. Кератоциты составляют 2,4—5,0% объема стромы.
Наиболее близки кератоциты по происхождению и строению к фиброцитам. Обнаруживаются они во всех участках стромы, но с различной плотностью. Использование конфокальной микроскопии позволило установить, что плотность кератоцитов в центральных участках роговой оболочки равняется 20,5 ±2,9 кл/ мм3. Отмечено также, что в передних слоях стромы их плотность меньше на 10%. Плотность кератоцитов уменьшается с возрастом примерно на 0,45% в год.
Кератоциты обладают длинными отростками, ориентированными параллельно коллагеновым пластинам. Контактируют отростки с отростками рядом расположенных клеток этого же уровня, а также и клетками других уровней стромы (рис. 3.2.3, 3.2.7). При этом между ними формируются межклеточные контакты типа щелевых контактов. Предполагают, что эти контакты служат взаимодействию между кератоцитами, расположенными в виде сети во всей строме роговицы.
Толщина кератоцитов равна примерно 2 мкм. При этом ядро выглядит непропорционально большим.
Иммуноморфологически в цитоплазме клеток выявлены коллагены III, V и VI типов.
Цитоплазма кератоцитов бедна органоидами. В прямом контакте с цитоплазматической мембраной можно обнаружить пятна базальноподобного волокнистого материала, особенно по периферии роговицы. Плотный контакт этого материала с коллагеновыми фибриллами стромы приводит к образованию периодической структуры. Вокруг многих кератоцитов отмечается скопление фибриллярного и зернистого материала, являющегося структурным компонентом будущих коллагеновых волокон и основного вещества. Кератоциты обладают большой степенью подвижности.
Основная функция кератоцитов — синтез межклеточного вещества и коллагеновых фибрилл в период эмбриогенеза, после повреждения роговицы, а также поддержание метаболизма стромы на протяжении всей жизни.
Birk и Trelstad установили, что поверхность фибробластов отвечает за пространственную ориентацию коллагеновых фибрилл. Именно благодаря этому свойству формируются пучки.
В связи с тем, что метаболическая активность кератоцитов в норме снижена, эндоплаз-матическая сеть клеток развита слабо. Лишь после травмы и воспалительных изменений роговицы эндоплазматическая сеть становится хорошо заметной.
В строме роговицы встречаются лимфоциты, макрофаги и полиморфноядерные лейкоциты.
Задняя пограничная (десцеметова) пластинка (lamina limitans postrior соrпеае; Decemett). Десцеметова оболочка при световой микроскопии выглядит бесструктурной мембраной, покрывающей заднюю поверхность стромы роговицы (рис. 3.2.3, 3.2.9).
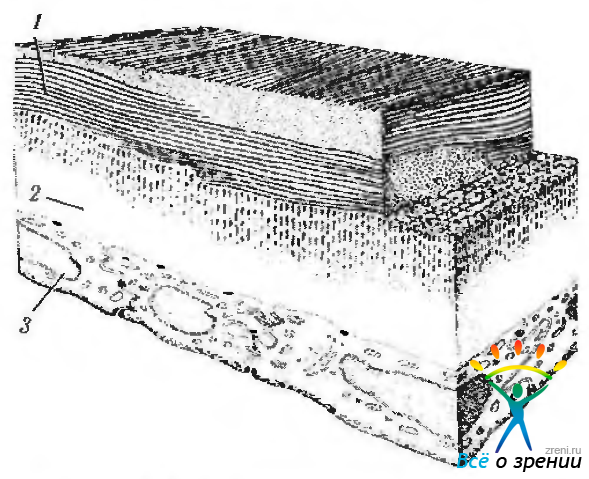
Рис. 3.2.9. Схема микроскопического строения задних слоев роговой оболочки (по Pouliquen, 1969): 1—строма роговой оболочки; 2—десцеметова оболочка; 3— задний эпителий (эндотелий)
В гистогенетическом и структурном смыслах она представляет собой базальную мембрану заднего эпителия роговицы (эндотелия), который ее и продуцирует. Эластичность является одной из наиболее важных ее характеристик. Волокна десцеметовой мембраны образуется на протяжении всей жизни человека. Толщина их при рождении равняется 3 мкм, а в старости — 8—12 мкм.
Как и другие базальные мембраны, десцеметова оболочка PAS-положительна и состоит из коротких и тонких фибрилл (10 нм). Фибриллы, в свою очередь, образованы коллагеном IV типа и погружены в гликопротеиновое основное вещество.
При ультраструктурном исследовании в мембране различают две области. Передняя ее треть имеет толщину 1—4 мкм и задние две трети — 5—15 мкм.
Передний слой десцеметовой оболочки, контактирующий со стромой, имеет многослойный пластинчатый вид, а задний — гранулированный. Именно передний слой возникает в эмбриональном периоде первым. На тангенциальных срезах этот слой состоит из однородных пластин коллагеновых волокон, образующих равносторонние треугольники. Треугольники связаны электронноплотными узлами. Эти соединения появляются на 5 месяце внутриутробной жизни, когда слой имеют толщину 3,1 мкм (2,2 — 4,5 мкм). Задние 2/3 мембраны образуются уже после рождения и состоят из гомогенного фиброгранулярного материала.
В мембране, помимо преобладающего коллагена IV типа, обнаружены коллагены III, V, VI и VIII типов.
С возрастом в десцеметовой мембране появляются, а затем увеличиваются в количестве коллагеновые волокна и слоистый материал. Этот процесс приводит к появлению на задней поверхности роговицы так называемых бородавок Хассал—Хенле (Hassal—Henle). При этом отмечается нарушение контактов между клетками эндотелия и нарушается барьерная функция последнего.
Несмотря на отсутствие в мембране Десцемета эластических волокон, она исключительно эластична. При травме нередко десцеметова оболочка скручивается в виде рулона, что обнаруживается при биомикроскопии. Десцеметова мембрана исключительно устойчива в отношении протеолитических ферментов.
Эндотелий (задний эпителий роговой оболочки). Эндотелий роговой оболочки представляет собой один слой плоских гексагональных клеток (плоский однослойный эпителий), расположенных на десцеметовой оболочке (рис. 3.2.3, 3.2.9—3.2.11).

Рис. 3.2.10. Плоскостной препарат эндотелия центральных участков роговой оболочки при исследовании его в фазово-контрастном микроскопе: отмечается полигональная форма клеток, их примерно одинаковые размеры и наличие плотных контактов между ними

Рис. 3.2.11. Схематическое изображение ультраструктурной организации клеток эндотелия роговой оболочки: 1 — микроворсинки; 2 — краевые выпячивания цитоплазмы в переднюю камеру глаза в местах межклеточных контактов; 3 — пиноцитозные пузырьки; 4 — центриоли; 5 — шероховатый эндоплазматический ретикулум; 6 — рибосомы; 7—ядерные поры; 8 — внутрицитоплазматические филаменты; 9 — аппарат Гольджи; 10 — межклеточные контакты различного типа
Наиболее распространено мнение о том, что они происходят из клеток нейрального гребня.
Эндотелий роговой оболочки рассматривают как один из наиболее важных структурных компонентов, обеспечивающих прозрачность роговицы. При этом показано, что обеспечение прозрачности роговицы связано со структурной организацией самой клетки, характера межклеточных контактов и расположением эндотелиальных клеток. Основной функцией эндотелиальных клеток при этом является поддержание постоянного гидростатического давления стромы роговой оболочки. Именно важная роль эндотелия в сохранении прозрачности роговицы явилась причиной многочисленных исследований, направенных на изучение строения и функции этой структуры глаза. Способствовало этому применение эндотелиальной прижизненной микроскопии.
Последние исследования показали, что у взрослых количество эндотелиальных клеток ограничено и довольно постоянно. Их количество порядка 500 000. С возрастом число клеток уменьшается. Наибольшее уменьшение плотности эндотелиальных клеток определяется в первые годы жизни и полностью коррелирует с увеличением площади роговой оболочки ребенка.
При использовании эндотелиальной микроскопии установлено, что плотность эндотелиальных клеток при рождении колеблется в довольно широких пределах (2627—5316 клеток в мм2). Плотность клеток падает примерно на 26% на первом году жизни. Дальнейшее падение плотности клеток на 26% отмечается на протяжении последующих 2 лет. Затем скорость уменьшения плотности клеток снижается и число клеток стабилизируется к среднему возрастут. Кривая, отражающая процесс уменьшения плотности клеток, имеет линейную или логарифмическую форму.
В процессе дифференциации уменьшается степень полиморфизма эндотелиального пласта, а также уменьшается количество клеток гексаганальной формы. Правда, необходимо отметить, что скорость уменьшения плотности и формы клеток колеблется в широких пределах и не дает исследователям возможности сделать окончательное заключение относительно значения этого процесса и факторов, влияющих на этот процесс.
У молодых людей размер клеток равен 18 — 20 мкм (высота — 5—6 мкм), а в более позднем возрасте — 40 мкм. Появляется бимодальность распределения клеток, как по размерам, таки по содержанию ДНК ядер.
Эндотелиальные клетки роговой оболочки присоединяются к десцеметовой оболочке при помощи полудесмосом. Рядом лежащие клетки плотно прилежат друг к другу и соединены десмосомами и запирательными пластинками. Запирательные пластинки распространяются по окружности апикальной поверхности клеток и закрывают межклеточные пространства, обеспечивая барьерные функции эндотелия. Рядом лежащие клетки соединяются также и при помощи «пальцевых вдавлений», представляющих собой цитоплазматические выросты, вдавливающиеся в тело соседней клетки. Несмотря на обилие межклеточных контактов, между клетками существуют щелевидные пространства, шириной 20 нм.
Наличие контактов между клетками предопределяет пропускную способность эндотелиального слоя. Они ограничивают пассивный транспорт в строму роговой оболочки. Любое проникновение жидкости в строму через межклеточные щели уравновешивается активным ионным транспортом, происходящим трансцеллюлярно. Процессы регуляции проникновения жидкости в строму могут быть нарушены при уменьшении плотности расположения эндотелиоцитов и расширении межклеточных пространств, что нередко наблюдается при патологических состояниях. Рядом исследователей установлены также критические пределы уменьшения количества эндотелиальных клеток. Резкое уменьшение плотности клеток приводит к необратимому нарушению гидратации стромы. Считается, что такой плотностью клеток является величина, равная 400—700 клеток в квадратном миллиметре (при норме 1400— 2500 клеток). Тем не менее клинические наблюдения показывают, что при ряде патологических состояний даже существенное снижение плотности клеток далеко не всегда сопровождается усилением гидратации стромы роговицы.
На апикальной поверхности каждой эндотелиальной клетки располагается от 20 до 30 микроворсинок высотой 0,5—0,6 мкм и шириной 0,1—0,2 мкм. Именно эти образования значительно увеличивают площадь контакта клеточной поверхности с влагой передней камеры глаза. Можно обнаружить и реснички. Они чаще видны по периферии роговицы. Обнаружение ресничек позволило Hogan, Alvarado, Weddell предположить, что эндотелиальные клетки имеют единое происхождение с клетками трабекулярной сети.
Цитоплазма эндотелиоцитов богата митохондриями, которые обеспечивают энергией активный транспорт, секрецию и высокий уровень синтеза протеинов. Эндотелиоциты содержат митохондрии в значительно большем количестве, чем любые другие клетки глаза за исключением рецепторных клеток. Обнаруживаются хорошо развитый гранулярный и агранулярный эндоплазматический ретикулум, многочисленные свободные рибосомы. Вблизи ядра четко виден аппарат Гольджи. Центриоли с ресничками располагаются в апикальной части клеток. В большом количестве определяются лизосомы. Отличительной чертой эндотелиальных клеток является наличие многочисленных пиноцитозных пузырьков, связанных с цитоплазматической мембраной (рис. 3.2.11). Экспериментальными исследованиями с использованием радиоактивной метки показано быстрое перемещение этих пузырьков через цитоплазму в сторону десцеметовой мембраны. Иммуногистохимически в цитоплазме эндотелиальных клеток выявлены основные гликозаминогликаны роговицы — хондроитин-6-сульфат, хондроитин-4-сульфат, гепаран-сульфат.
Необходимо остановиться и на основных физиологических функциях эндотелия роговицы. Одной из них является обеспечение клеток стромы питательными веществами. Процесс транспорта питательных веществ обеспечивается или диффузией между эндотелиоцитами, или активным переносом через содержимое клетки в направлении стромы.
Эндотелий играет главную роль в поддержании прозрачности роговицы путем активной регуляции содержания в строме воды. Эту функцию он выполняет, используя два механизма. Во-первых, он является активным барьером для солей и ряда метаболитов, проникновение которых в строму приводит к отеку последней. Во-вторых. он активно снижает осмотическое давление стромы наличием так называемого бикарбонатного насоса, возвращающего ионы из стромы назад в камерную влагу.
Кровоснабжение и лимфатическое дренирование роговицы обеспечиваются конъюнктивальными, эписклеральными и склеральными сосудами, являющимися ветвями передних ресничных артерий.
Продолжение в следующей статье: Роговая оболочка ? Часть 2
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0