Сетчатка │ Часть 2


Описание
Сенсорная часть сетчатки
Сенсорная часть сетчатки представляет собой тонкую прозрачную оболочку, содержащую чувствительные к свету клетки, которые и превращают световую энергию в нервные импульсы. При диафаноскопии глазного яблока сетчатка выглядит пурпурно-красной из-за наличия в фоторецепторах зрительного пигмента (родопсин). Однако этот цвет быстро исчезает при освещении энуклеированного глаза на протяжении 5—10 минут. При этом сетчатка становится белой и полупрозрачной.
Толщина сетчатки в области зрительного нерва равняется 0,4 мм. Она истончается в области зубчатой линии с назальной стороны до 0,15 мм. Темпорально ее толщина 0,4 мм. В области центральной ямки (0,2 мм).
Основу сенсорной части сетчатки составляют нервные клетки — фоторецептор, биполярная и ганглиозная клетки, ассоциативные горизонтальные нейроны, амакриновые клетки, а также глиальные элементы — клетка Мюллера, фиброзные и протоплазматические астроциты, микроглия и олигодендроциты.
Фоторецепторы (палочки и колбочки). Слой палочек и колбочек является самым наружным слоем сенсорной сетчатки. Складывается он из цитоплазматических выростов палочек и колбочек фоторецепторных клеток. Фоторецепторы являются не чем иным, как высокоспециализированными нейроэпителиальными клетками. По структуре и направленности выполняемой функции они близки к рецепторным клеткам других тканей и органов (тельца Пачини, Краузе, Мейснера).
Тела фоторецепторных клеток располагаются в плоскости наружной пограничной мембраны, а их апикальные отростки (внутренние сегменты) лежат только снаружи этой мембраны.
Большое значение имеет знание распределения и пространственной ориентации фоторецепторных клеток, что в значительной мере способствует пониманию зрительных связей в сетчатке. Плотное расположение фоторецепторов и их точная ориентация вдоль зрительной оси обеспечивают детальный анализ поля зрения. Любое изменение расположения фоторецепторов приводит к нарушению зрения. Если между фоторецепторами появляются пространства (при центральной серозной ретинопатии) и они неравномерно распределены, развивается микропсия. Нарушение ориентации фоторецепторов вдоль зрительной оси приводит к метаморфопсии.
Фоторецепторы распределяются закономерным образом, в виде мозаики (рис. 3.6.13).
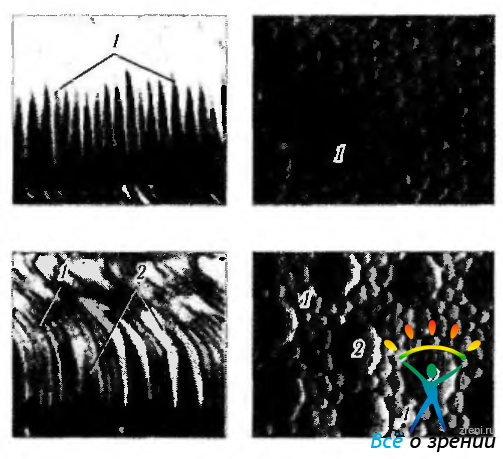
Рис. 3.6.13. Особенности «мозаичного» строения периферии сетчатки (а) и области центральной ямки (б): 1 — палочки; 2 — колбочки. Слева иллюстрируется срез сетчатки, а справа — плоскостной препарат (по Curcio et al., 1990)
В области желтого пятна лежат только колбочки. Вне желтого пятна колбочки кольцевидно окружены палочками.
В сетчатой оболочке обнаруживается от 77,9 до 107,3 млн (в среднем 92 млн) палочек и 4,08—5,29 млн (в среднем 4,6 млн) колбочек.
Существуют индивидуальные отличия плотности палочек и колбочек в зависимости от топографического отдела сетчатки. Наибольшее разнообразие плотности выявляется вблизи центральной ямки и у зубчатой линии, а наименьшее — в средней части сетчатки и по периферии.
Плотность колбочек максимальна в области центральной ямки (199 000 колбочек в мм2). При этом их число колеблется в широких пределах (от 100 000 до 324 000 колбочек в мм2). По мере удаления от центральной ямки плотность колбочек существенно уменьшается. Так, плотность колбочек уменьшается до 75 000 мм2 в 130 мкм от центра центральной ямки. Примерно в трех миллиметрах от центра центральной ямки отмечается наибольшая плотность палочек, а плотность колбочек уменьшается. Степень этого уменьшения различна в зависимости от направления. Так, плотность колбочек с назальной стороны на 40— 45% выше, чем с темпоральной. В периферических отделах сетчатки плотность колбочек опять возрастает (рис. 3.6.13—3.6.15).
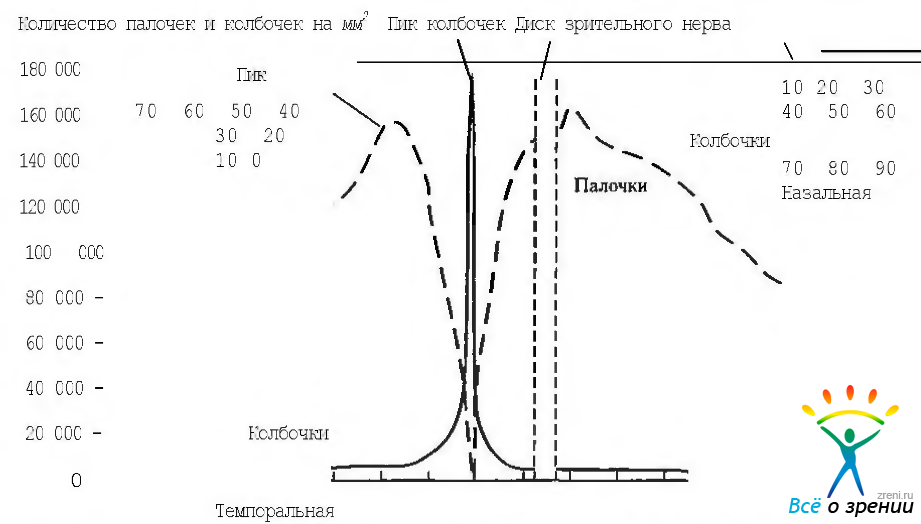
Рис. 3.6.14. Плотность палочек и колбочек вдоль горвонтального мерцщана (по Osterberg, 1935)
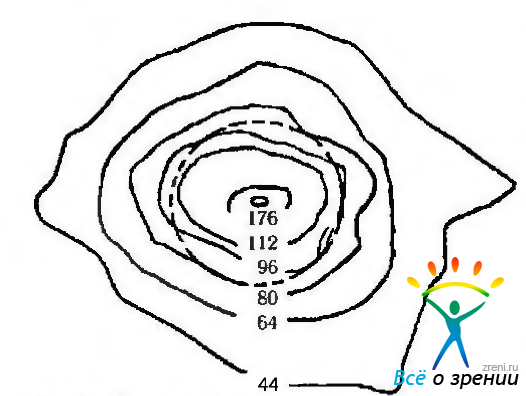
Рис. 3.6.15. Топографические особенности распределения плотности колбочек в области центральной ямки (no Curcio et al., 1987): контурные сплошные линии очерчивают области с количеством колбочек в одном квадратном миллиметре, равном цифре, приведенной на рисунке и умноженной на 1000. Окружность (пунктирная линия) очерчивает поле зрения, равное 1 градусу
Считают, что пространственное расположение колбочек в области желтого пятна является фактором, определяющим разрешающую способность глаза. Так, среднее расстояние между центрами колбочек колеблется от 2,53 ± 0,29 мкм до 6,16 ± 1.04 мкм. Наименьшее расстояние между клетками обнаружено в области центральной ямки. Это свидетельствует о наибольшей разрешающей способности сетчатки именно в этой области.
Необходимо отметить, что данные психофизиологических исследований относительно остроты зрения не полностью совпадают с приведенными выше анатомическими данными. По всей видимости, большое значение имеют другие факторы. Единственная область в сетчатке, где функциональная острота зрения совпадает с анатомической разрешающей способностью, располагается между 0,2 и 2,0°. Интересно, что острота зрения у новорожденных на два порядка ниже, чем у взрослых. В ближайшее время после рождения колбочки, палочки и клетки пигментного эпителия перемещаются к центру желтого пятна. При этом дифференциация фоторецепторов в центре сетчатки происходит медленнее, чем по периферии. Изучение сетчатки обезьян показало, что плотность колбочек, свойственная взрослым животным, появляется только к 15—18 месяцам после рождения. У человека плотность колбочек нарастает вплоть до 5—8-лет-него возраста. Наиболее важным фактором, определяющим низкую остроту зрения у новорожденных, является не плотность расположения колбочек, а неполная дифференциация желтого пятна. Косвенным подтверждением этому является альбинизм. У этих больных острота зрения низкая, а желтое пятно в структурном отношении напоминает желтое пятно новорожденного.
Сниженная острота зрения у этих больных связана также с недостаточностью развития межнейронных связей на уровне наружного коленчатого тела и зрительной коры головного мозга.
Плотность папочек и их распределение также являются объектом пристального внимания исследователей. Установлено, что диаметр свободной от палочек области желтого пятна равняется 0,35 мм. Это соответствует 1,25 градуса поля зрения (рис. 3.6.14, 3.6.15). Самая высокая концентрация палочек выявлена в области сетчатки, имеющей вид горизонтального эллипса. Этот эллипс несколько расширяется в носовом направлении и кверху. Именно от этого места плотность палочек медленно уменьшается по мере продвижения к периферии сетчатки.
С назальной стороны плотность палочек на 20—25% выше, чем с височной стороны. В верхней половине сетчатки палочек больше на 2%, чем в нижней половине. Равное соотношение палочек и колбочек обнаруживается на расстоянии 0,5 мм кнутри и на 0,4 мм выше центральной ямки.
Внутренние и наружные сегменты фоторецепторов. Внутренние и наружные сегменты фоторецепторов являются местом трансформации световой энергии в нервный импульс. Они имеют следующее строение. От тела фоторецепторной клетки отходит цитоплазматический вырост. Этот вырост подразделяется на две части — внутренний и наружный сегменты (рис. 3.6.16).
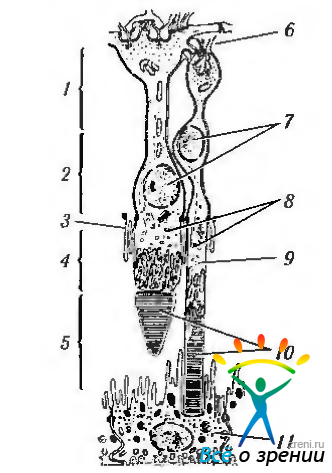
Рис. 3.6.16. Схематическое изображение особенностей и внутренних члеников палочек и колбочек (по Kolb et al., 1998): 1 — наружный плексиформный слой; 2-слой; 3 — наружная пограничная мембрана; 4 - наружный ядерный внутренний сегмент; 5 — наружный сегмент; 6 — синаптическое тело; 7 — ядра; 8 -миоидная часть; 9 — эллипсоидная часть; 10 — диски фоторецепторов; 11— пигментный эпителий
Наружный сегмент лежит в интерфоторецепторном матриксе и обращен к апикальной поверхности клеток пигментного эпителия. Основной функцией этого образования является преобразование световой энергии в электрические импульсы. В дальнейшем, нервные импульсы обрабатываются на уровне сетчатки и передаются по зрительному нерву коре головного мозга. Восприятие света и преобразование его в нервный импульс начинаются с активации последовательных реакций фотохимической стереоизомеризации зрительного пигмента, расположенного на дисках наружного сегмента фоторецепторов. К зрительным пигментам относятся родопсин и иодопсин. Родопсин, обнаруживаемый в палочках, обеспечивает фотопическое зрение.
Фотопическое зрение происходит в колбочках и обеспечивается трихроматическими пигментами. Колбочки содержат одну из трех молекул иодопсина, поглощающих свет в трех различных спектрах — 440 нм (синий), 540 нм (зеленый) и 577 нм (оранжевый). Обозначаются эти колбочки как S- (коротковолновые), М- (средневолновые) и L- (длинноволновые). Более подробные сведения о химических процессах, происходящих в фоторецепторных клетках в процессе формирования нервного импульса будут приведены в разделе «Зрительные пигменты и фоторецепция».
Для понимания механизмов цветового зрения большое значение имеет морфологическая дифференциация различных типов колбочек. Это необходимо для определения их связи с другими нейронами сетчатки. В настоящее время морфологическими методами дифференцируют средне- и длинноволновые колбочки у некоторых рыб, лягушек, птиц и рептилий. К сожалению, у приматов и человека возможна только дифференциация коротковолновых колбочек (S-колбочка) от остальных. У «синей» колбочки более длинный и больший диаметр внутреннего членика, который интенсивно окрашивается. Кроме того, «синие» колбочки не столь равномерно и закономерно распределены в сетчатой оболочке. Они составляют 3—5% от общего числа фоторецепторов в центре желтого пятна, и их число увеличивается до 15% на склоне области желтого пятна. Использование антител, обладающих аффинитетом к синему опсину, подтвердило то, что «синие» колбочки в области желтого пятна редки и лежат изолированно или полностью отсутствуют в зоне, расположенной недалеко от пика наибольшей плотности колбочек. Диаметр этой зоны равен 100 мкм (0,35 градуса). Самая высокая плотность «синих» колбочек (более чем 2000 клеток в мм2) выявлена в зоне шириной 0,1—0,3 мм.
Наружные членики колбочек и палочек являются результатом выпячивания плазматической мембраны фоторецептора. Наружный членик соединяется с внутренним сегментом посредством цитоплазматического перешейка. Общая длина обоих сегментов определяется локализацией и типом фоторецептора.
Наружный членик постоянно обновляется. Этот процесс иллюстрируется рис. 3.6.11, 3.6.12. При этом постоянно регенерируют и зрительные пигменты. Опсиновая часть молекулы родопсина синтезируется аппаратом Гольджи фоторецепторной клетки. Другая составная часть зрительного пигмента (ретинал — производное витамина А) поставляется дискам наружных члеников клетками пигментного эпителия сетчатки при помощи транспортной молекулы.
Внутренние и наружные членики палочек имеют длину 40—60 мкм на всем протяжении сетчатки. Длина сегмента колбочки максимальна в области желтого пятна (80 мкм) и постепенно уменьшается до 40 мкм к периферии сетчатки. В области зубчатой линии колбочки короче (4 мкм) и толще. Наружный сегмент палочки (длина 25—28 мкм и диаметр 1 — 1,5 мкм) не изменяется на протяжении всей сетчатки.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Необходимо отметить, что колбочки в области желтого пятна напоминают по форме и размеру палочки. Наружные сегменты колбочек вдали желтого пятна имеют диаметр 6 мкм в основании и 1,5 мкм на верхушке.
Наружные членики (сегменты) палочек имеют цилиндрическую форму и содержат плотно упакованные двойные дисковидные пластины, количество которых колеблется от 600 до 1000. Каждый диск имеет толщину 22,5—24,5 мкм. Расстояние между дисками равно 21 мкм. Никаких специализированных контактов между дисками, а также дисками и цитоплазматической мембраной не обнаруживается. Наружная поверхность каждого наружного сегмента покрыта слоем нейрокератана.
В дисках содержится до 90% молекул зрительного пигмента. Остальное количество его рассеяно по поверхности плазматической мембраны. Наружный и внутренний сегменты соединяет модифицированная ресничка. Именно в месте перехода наружного сегмента во внутренний сегмент цитоплазма суживается. Ширина этого перешейка равна 0,3 мкм, а длина 1 мкм. В основании внутреннего сегмента лежит базальное тельце, состоящее из одной пары центриолей. Ресничка состоит из девяти пар микротрубочек, расположенных кольцевидно. Пучки филаментов исходят из базального тельца и простираются поперек эллипсоидной части цитоплазмы, заканчиваясь в миоидной области цитоплазмы. В фибриллах ресничек высока активность АТФ-азы, свидетельствующая об интенсивном метаболизме этого образования. Правда, функции ресничек пока неизвестны.
Цитоплазма поверхности наружного сегмента формирует 9—12 микроворсинок, длиной 12,5 мкм. Функция микроворсинок пока неизвестна.
Внутренние членики (сегменты) палочек имеют цилиндрическую форму. Гистологически различают две части внутреннего членика: эозинофильную наружную, называемую эллипсоидной частью, и внутреннюю базофильную, называемую миоидной частью. Тинкториальные свойства этих двух областей изменяются в зависимости от метаболической активности фоторецептора. Эллипсоидная часть окрашивается эозинофильно в связи с наличием в ней большого количества митохондрий. В одной палочке можно найти до 600 митохондрий. Цитоплазма также содержит гладкий эндоплазматический ретикулум, нейротрубочки, свободные рибосомы и гранулы гликогена. Базофилия миоидной части зависит от большой концентрации в ней свободных рибосом. Миоидная область является центром белкового синтеза. По этой причине в ней располагаются шероховатая эндоплазматическая сеть, аппарат Гольджи, микротрубочки, микрофиламенты и гранулы гликогена. Таким образом, основной функцией этой части фоторецептора является обеспечение метаболических и синтетических функций клетки.
Наружные сегменты (членики) колбочек имеют различное строение в различных участках сетчатки. В области зубчатой линии и по периферии сетчатки они короткие и конические, а в fovea centralis продолговатые, напоминают наружные сегменты палочек.
Ультраструктурными исследованиями установлено, что наружный сегмент колбочки имеет больше дисков (1000—1200), чем наружный сегмент палочки. Междисковые пространства палочки более широкие (в колбочках — 3,5 мкм, в палочках— 16,5 мкм).
В отличие от дисков палочек диски колбочек соединены между собой и прикрепляются к плазматической мембране.
Внутренние сегменты (членики) колбочек. Наружные и внутренние членики колбочек связаны друг с другом посредством тонкого цитоплазматического перешейка, содержащего видоизмененную ресничку. Они изменяются в зависимости от их топографического расположения. В центральной ямке (fovea centralis) они более длинные и узкие. Ультраструктурная организация внутреннего сегмента палочек и колбочек одинаковая, за исключением того, что в колбочках значительно больше митохондрий (200—300 на срезе).
Наружная поверхность миоидной части палочек и колбочек покрыта волосоподобными цитоплазматическими отростками мюллеровских клеток, формирующих «корзины Шульца». Благодаря этому никакого контакта между смежными клетками нет. Отростки мюллеровских клеток участвуют также в регуляции состава внеклеточной среды фоторецепторов и служат для жесткой пространственной фиксации палочек и колбочек.
Наружная пограничная мембрана. При световой микроскопии видно, что наружная пограничная мембрана (рис. 3.6.1) отделяет слой палочек и колбочек от подлежащего наружного ядерного слоя сетчатки. Она простирается от диска зрительного нерва до зубчатой линии, где превращается в базальную пластинку, расположенную между пигментированными и бес-пигментными частями ресничного эпителия. Наружная пограничная мембрана представляет собой не что иное, как скопление в одной плоскости терминальных пластинок (zonulae adherentes), расположенных между мюллеровскими клетками и фоторецепторами, между смежными мюллеровскими клетками и, редко, между соседними фоторецепторами.
Наружная пограничная мембрана, таким образом, не является истинной мембраной. Через нее проходят небольшие молекулы. Главной функцией мембраны является обеспечение функционирования избирательного барьера на пути питательных веществ, которые проходят между рядом расположенными мюллеровскими клетками, а также стабилизация положения фоторецепторов.
Наружный ядерный слой. Наружный ядерный слой находится кнутри от наружной пограничной мембраны и содержит тела и ядра фоторецепторных клеток (рис. 3.6.1). В зависимости от участка сетчатки ширина этого слоя изменяется, прежде всего, из-за изменения числа рядов ядер.
С назальной стороны диска наружный ядерный слой имеет толщину 45 мкм и состоит из 8—9 рядов ядер. С височной стороны он состоит только из четырех рядов ядер, истончаясь до 22 мкм. В желтом пятне наличие 10 рядов ядер колбочек увеличивают ширину наружного ядерного слоя до 50 мкм. В области зубчатой линии наружный ядерный слой состоит только из одного слоя ядер колбочковых клеток, которые плотно прилежат к наружной пограничной мембране и четырем рядам ядер палочек, расположенным кнутри от них. Толщина ядерного слоя при этом приблизительно 27 мкм.
Ядра колбочек овальные и имеют диаметр 5—7 мкм. Расположены они на 3—4 мкм кнутри от наружной пограничной мембраны. Ядра палочек также овальные, диаметром 5,5 мкм.
Цитоплазма обоих типов клеток скудная. Тела палочек и колбочек окрашиваются по-разному. При использовании метода Унна тело палочек не окрашивается, а колбочки окрашиваются в интенсивно синий цвет. Используя трехцветный метод Маллори, после фиксации сетчатки жидкостью Ценкера можно четко дифференцировать центральную ямку. Центральная ямка окрашена в интенсивно красный цвет. Это связано с тем, что методом Маллори окрашиваются только колбочки.
Наружный сетчатый (плексиформный) слой (рис. 3.6.1) является местом соединения первого и второго нейронов, т. е. местом передачи информации от первого нейрона (фоторецептора) второму (биполярной клетке). Помимо указанных клеток в нем располагаются ассоциативные нейроны (горизонтальная клетка).
Две трети слоя состоит из внутренних волокон фоторецепторов, окруженных отростками мюллеровских клеток. Треть слоя состоит из дендритов биполярных и горизонтальных клеток, а также отростков мюллеровских клеток. Наружный плексиформный слой наиболее толстый в области желтого пятна (51 нм). Состоит он из косо идущих волокон, отклоняющихся от желтого пятна. Этот слой также известен как слой волокон Хенле.
Внутренние волокна в наружном плексиформном слое представляют собой аксоны палочек и колбочек. Диаметр аксона палочки приблизительно в четыре раза больше, чем у колбочки. Они содержат типичные органоиды — единичные митохондрии, немного свободных рибосом, гладкий эндоплазматический ретикулум, гранулы гликогена и плотно упакованные микротрубочки.
Синаптическая связь палочек со вторым нейроном происходит при помощи овальных расширений цитоплазмы диаметром 1 мкм. Называются они сферулами.
Синапсы колбочек отличаются. Эти отличия сводятся к тому, что колбочки образуют так называемую «ножку», т. е. ножкоподобное утолщение окончания цитоплазматического отростка колбочки. «Ножка» больше, чем сферула (7—8 мкм в парафовеолярной области и 5 мкм в области фовеа). Теперь мы более подробно остановимся на синаптических связях этого слоя.
Синапсы палочек. Синаптический комплекс палочек состоит из указанной выше пресинаптической сферулы, синаптической ленты и постсинаптических отростков, принадлежащих горизонтальным или биполярным клеткам (рис. 3.6.20—3.6.23).
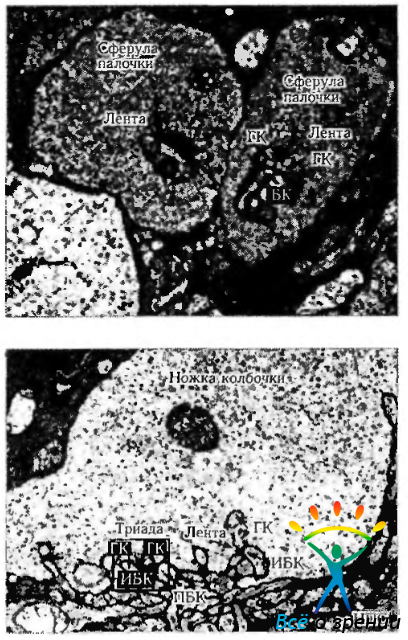
Рис. 3.6.20. Ультраструктурные особенности сферул палочек (а) и «ножек» колбочек (б) (по Kolb, 1998): НПС — наружный плексиформный слой; ГК—горизонтальная клетка; БК—биполярная клетка палочки; ИБК—инвагинирующая биполярная клетка; ПБК — плоская биполярная клетка
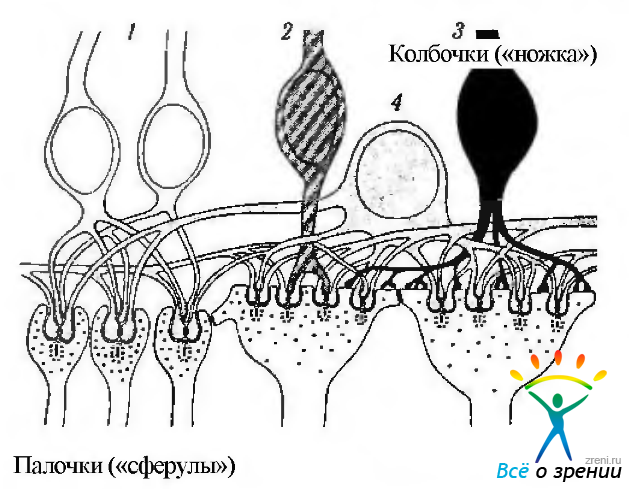
Рис. 3.6.21. Схематическое изображение синаптических тел палочки и колбочки: 1 — палочковые биполярные клетки; 2— карликовая биполярная клетка; 3 — плоская биполярная клетка; 4 — горизонтальная клетка. Отмечается наличие контакта между сферулой палочки и «ножкой» колбочки. Синаптические тела палочки и колбочки соединяются непосредственно, а также при помощи горизонтальной клетки
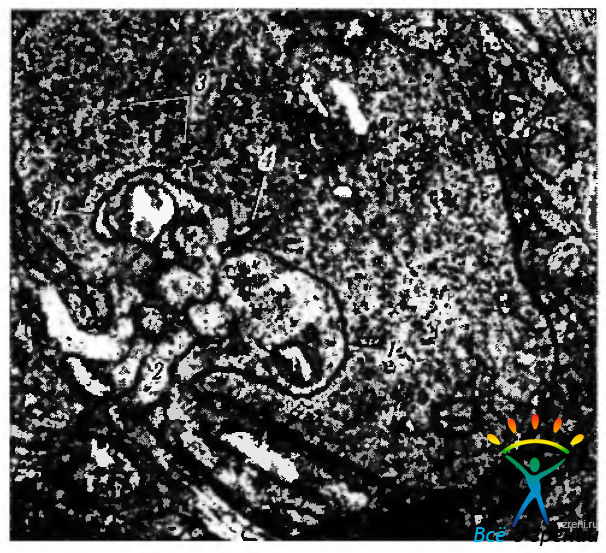
Рис. 3.6.22. Электроннограмма сферулы палочки: 1 — латеральные отростки горизонтальной клетки; 2 — отростки биполярной клетки; 3 — синаптические пузырьки; 4 — синаптическая лента

Рис. 3.6.23. Особенности синаптических связей палочек и колбочек с биполярными клетками и биполярных клеток с ганглиозными: видно, что одна биполярная клетка получает информацию от нескольких палочковых фоторецепторов и только от одной колбочки
Сферулы содержат многочисленные пресинаптические пузырьки, а также митохондрии и нейротрубочки. Плотность пре- и постсинаптической мембраны увеличивается вблизи синаптической щели (ширина синаптической щели 15 мкм). Перпендикуляр, проходящий через пресинаптическую мембрану, называется синаптической лентой, состоящей из трех электронноплотных слоев, каждый из которых имеет толщину 12 мкм. Отделяется она светлой зоной, имеющей толщину 40 мкм, и окружена ореолом пузырьков. Сферулы палочек содержат только две синаптические ленты, которые ассоциируются с двумя боковыми элементами, являющимися терминалами аксонов горизонтальных клеток, и двумя дендритами биполярных клеток палочек (рис. 3.6.22).
С одной сферулой палочки может входить в контакт несколько различных горизонтальных клеток (1—4 клетки). Различают два основных типа контактов — с телодендритами горизонтальной клетки и дендритом биполярной клетки. Каждая сферула входит в контакт с 4 биполярными клетками. В то же время каждая биполярная клетка контактирует с 50 палочками (вне фовеолы) и с несколькими сотнями палочек по периферии сетчатки.
Эти различия в характере межнейронных связей соответствуют различиям в разрешающей способности зрительной системы.
Синапсы колбочек. «Ножка» колбочки пирамидальной формы. Синаптические вдавления на «ножке» объединяют одновременно три нейрона, контактирующие в то же самое время и между собой. Подобная структура получила название «триада» (рис. 3.6.20, 3.6.21). Центральный аксон триады принадлежит биполярной клетке. Это аксон может входить в контакт с той же самой колбочкой в 10—25 различных точках. Два дендрита с обеих сторон триады исходят из различных горизонтальных клеток. Хотя только одна биполярная клетка входит в контакт с одной «ножкой» колбочки, контакт существует со многими горизонтальными клетками, число которых обычно 6—8. Такая «ножка» имеет также множество маленьких поверхностных вдавлений (так называемых базальных соединений), контактирующих с плоской диффузной биполярной клеткой. Подобный тип синапсов биполярных клеток формируется сразу с шестью колбочками. Базальные соединения представляют собой классические эксцитаторные (возбуждающие) синапсы и функционируют подобно щелевым контактам.
Присутствие многочисленных десмосом между отростками клеток наружного плексиформного слоя (десмосомы) препятствует свободному распространению в сетчатке метаболитов, жидкостей и экссудата.
Помимо биполярных и горизонтальных клеток, фоторецепторы контактируют и между собой. Палочки контактируют с палочками и колбочками. Происходит это благодаря так называемым щелевым контактам. От «ножки» колбочки отходят тонкие отростки, которые подходят к сферулам палочек и «ножкам» других колбочек. В тех местах, где эти отростки (называемые телодендритами) образуют щелевой контакт, формируется «электрический контакт», т. е. происходит передача информации без использования нейротрансмиттера. 3—5 подобных контактов определяется на одной сферуле палочки, образованной телодендритами колбочки. Одна «ножка» колбочки может иметь до 10 контактов с соседними палочками. «Ножки» S-колбочек («синих») не содержат такого большого количества контактов. По этой причине S-колбочки довольно изолированы.
Функциональное значение прямой электрической связи между различными типами фоторецепторов не совсем понятно. Первоначально многие исследователи предполагали, что такие связи разрушают пространственную интеграцию фоторецепторов и соответственно возможность анализа функционирования цветового зрения, «смешивая» информацию, получаемую от палочек и колбочек. Тем не менее на основании многочисленных физиологических экспериментов установлено, что колбочки благодаря этим связям могут нести информацию палочек. Это, при определенных условиях, может иметь большое физиологическое значение. При этом изучены интимные механизмы этого процесса, правда, с использованием экспериментальных животных.
Внутренний ядерный слой. Внутренний ядерный слой состоит из 8—12 рядов плотно упакованных ядер биполярных, горизонтальных, амакриновых, межплексиформных и мюллеровских клеток. При световой микроскопии можно различить четыре слоя, преимущественно содержащих тот или иной клеточный тип:
- Слой горизонтальных клеток (наиболее наружный).
- Слой биполярных клеток (наружный про межуточный слой).
- Слой мюллеровских клеток (внутренний промежуточный).
- Слой амакриновых и межплексиформных клеток (самый внутренний).
Горизонтальные клетки (рис. 3.6.24, 3.6.25, 3.6.26).

Рис. 3.6.24. Особенности строения тел и дендритного поля различных типов горизонтальных клеток человека. Световая микроскопия (импрегнация серебром) (по Kolb, 1998)
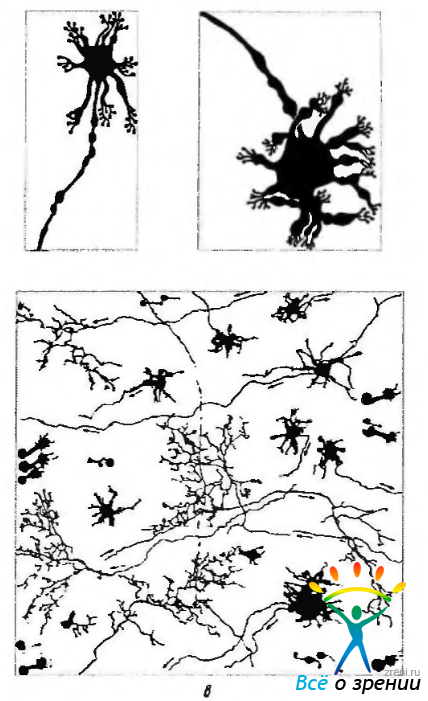
Рис. 3.6.25. Схематическое изображение различных типов горизонтальных клеток: а — горизонтальная клетка, контактирующая с колбочковым фоторепептором; б—горизонтальная клетка, контактирующая с палочковым фоторецептором; в — схематическое изображение характера контакта горизонтальных клеток различного типа в плоскости сетчатки
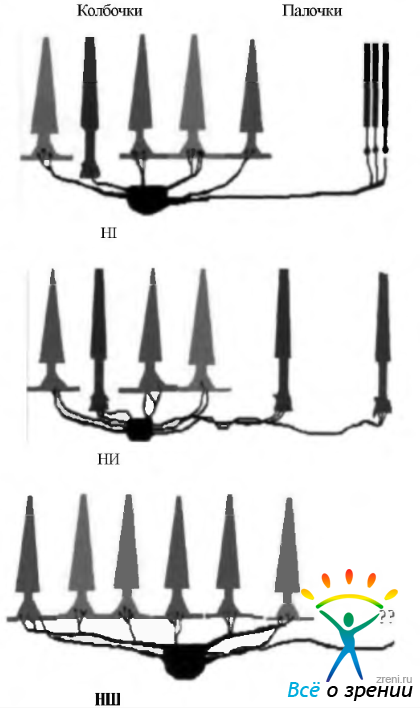
Рис. 3.6.26. Особенности соединения горизонтальных клеток с колбочками различных спектральных характеристик (пo Anhelt, Kolb, 1994)
Отростки горизонтальных клеток, в отличие от биполярных, образуют сеть, расположенную в горизонтальной плоскости и объединяющую фоторецепторы различных участков сетчатки.
Наибольшее количество горизонтальных клеток в области центральной ямки. Постепенно по мере продвижения к периферии сетчатки их число снижается. Горизонтальные клетки имеют короткие отростки, а аксон не ветвится вблизи тела клетки (на протяжении 200— 300 мкм). Длина аксона может достигать 2 мм.
В зависимости от размера клетки, особенностей строения синапсов между дендритами и аксонами, а также площади дендритного поля различают три типа горизонтальных клеток. Обозначаются они как клетки типов HI, НИ и НШ. Клетка HI отличается от остальных длинными мощными дендритами, входящими в контакт с колбочками при помощи «триад». Их аксон контактирует с палочкой. При этом образуются так называемые «точечные» синапсы. Аксон также образует синапсы на дендритах биполярных клеток палочек.
Приведенная выше схема синаптических связей является основой обработки информации, получаемой от многочисленных палочек. При этом информация собирается по площади сетчатки, превышающей площадь дендритного поля клетки.
Клеток типа НШ на 30% больше, чем типа НI, и они контактируют с колбочками.
Клетки типа НИ имеют тонкие дендриты и короткие (100—300 мкм) аксоны. Как дендриты, так и аксоны контактируют только с колбочками.
Недавние электронномикроскопические исследования сетчатки человека показывают, что существуют определенные закономерности в контакте между колбочками различных спектральных характеристик и различными типами горизонтальных клеток (рис. 3.6.26). Необходимо отметить, что клетки типа HI контактируют с колбочками всех спектральных типов. Наименее часто они контактируют с коротковолновыми («синими») колбочками. Клетки типа НТ, наоборот, чаще контактируют именно с «синими» колбочками, а клетки НШ с колбочками вообще не контактируют. На основании этих данных предполагают, что клетки типа HI можно рассматривать как клетки «яркости», а клетки типов НИ и НШ как клетки воспринимающие цвета.
Горизонтальные клетки млекопитающих характеризуются также наличием многочисленных «щелевых контактов» между дендритами соседних клеток. Благодаря этим контактам сигнал распространяется в плоскости синцития нейронов сетчатки. Помимо электрического сигнала через эти контакты могут проходить и низкомолекулярные вещества.
Строение тела горизонтальных клеток различных типов схоже. Тело клетки обычно уплощено и имеет диаметр 6—8 мкм. Ядро круглое и окружено аппаратом Гольджи. Цитоплазма содержит гладкую и шероховатую эндоплазматическую сеть, четкие митохондрии и многочисленные свободные рибосомы. Характерной особенностью горизонтальных клеток является наличие в цитоплазме включений, так называемых телец или «кристаллоида Колмера», описанного Колмером еще в 1918 г. Эти образования имеют длину 8—20 мкм и ширину 0,3—1,5 мкм и чаше обнаруживаются вблизи ядра, но видны и в цитоплазматических отростках. Состоят они из пакетов параллельно расположенных плотных трубочек в количестве от 5 до 30, отделенных промежутком шириной 2—6 мкм. Каждая трубочка складывается из 2—3 концентрических мембран, на внутренней и внешней поверхностях которых лежат рибосомоподобные частицы, чувствительные к рибонуклеазе. Предполагают, что эти образования представляют собой своеобразную форму шероховатой эндоплазматической сети. Функции горизонтальных клеток разнообразны. Более подробно о них будет изложено в 4-й главе. Здесь мы остановимся лишь на некоторых из них.
Во-первых, необходимо указать, что горизонтальные клетки интегрируют сигналы, поступающие от палочек и колбочек с выделением так называемых «каналов» передачи информации различного типа. При этом именно на уровне горизонтальных клеток уже четко определяется формирование структурно-функциональных нейронных единиц — «рецептивных полей», имеющих фундаментальное значение в обработке зрительной информации и передаче ее более высоко расположенным отделам центральной нервной системы. Именно благодаря «рецептивным полям» и формируются основные физиологические характеристики зрительного восприятия, такие как «контрастность», «цветовое зрение» и др.
Во-вторых, на основании выявления нейронных связей между горизонтальными клетками и фоторецепторами, а также физиологических исследований установлено, что горизонтальные клетки посылают зрительную информацию через синапсы обратной связи назад к фоторецепторам. Эти обратные связи способствуют функционированию «рецептивных полей».
В-третьих, благодаря наличию избирательных многоконтактных обратных связей горизонтальных клеток с палочками и колбочками различных спектральных характеристик, именно горизонтальные клетки объединяют и обрабатывают весь широкий спектр цветовой информации.
Биполярные клетки (рис. 3.6.27, 3.6.28).
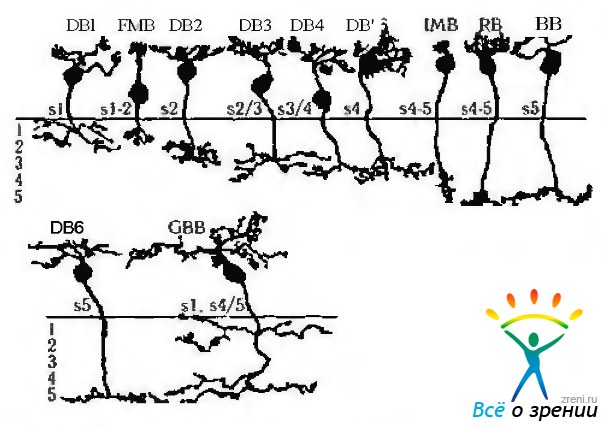
Рис. 3.6.27. Основные типы биполярных клеток сетчатки человека (по Kolb, 1998): DB— клетки диффузного типа; MB — карликовые клетки; ВВ — клетки «синих» колбочек; GBB — гигантские двухслойные; RB биполярные клетки палочек. Приведенные слева цифры указывают уровень распространения дендритов клеток во внутреннем плексиформном слое

Рис. 3.6.28. Ультраструктурные особенности синаптических контактов биполярных клеток на уровне внутреннего плексиформного слоя (по Kolb, 1998): 1 - амакриновая клетка; 2 — биполярная клетка; 3—ганглиозная клетка. Кружками указаны места формирования синапсов между различными типами клеток
Биполярные клетки являются вторым нейроном зрительного пути. В каждой сетчатке содержится приблизительно 35 676 000 подобных клеток.
Тела этих клеток располагаются во внутреннем ядерном слое, а их отростки распространяются на наружный и внутренний плексиформные слои.
Диаметр тела клетки в области желтого пятна равен 9 мкм, а в периферических отделах сетчатой оболочки — 5 мкм. В зависимости от типа синаптических отношений с другими клетками различают 9 основных типов биполярных клеток. Восемь типов клеток относятся к биполярным клеткам колбочек и один тип к биполярным клеткам палочек. Это следующие типы:
- Биполярные клетки палочек (щеткоподобные).
- Инвагинированные карликовые.
- Плоские карликовые.
- Плоские диффузные.
- Инвагинированые диффузные.
- Биполярные клетки «синих» колбочек, образующие ON-центр «рецептивные поля».
- Биполярные клетки «синих» колбочек, образующие OFF-центр «рецептивные поля».
- Гигантские двухслойные.
- Гигантские диффузные инвагинированные.
Биполярные клетки палочек (щеткоподобные) составляют 20% от общего числа биполярных клеток. Расположены они на расстоянии 1 мм от желтого пятна. Диаметр дендритного дерева клеток увеличивается по мере приближения клеток к периферии сетчатки.
В наружном плексиформном слое основной дендрит клетки делится на 2—3 ветви, которые после прохождения между «ножками» колбочек образуют в виде щеточек отростки, проникающие в сферулу палочки.
В центральных участках сетчатки дендритное поле горизонтальных клеток маленькое (15 мкм) и дендриты контактируют с 15—20 палочками. По периферии сетчатки дендритное поле больше (до 30 мкм) и клетка входит в контакт с 40—50 палочками.
Аксоны биполярной клетки палочки во внутреннем плексиформном слое образуют синапсы с отростком амакриновой клетки, дендритами и телами клеток диффузных ганглиозных клеток (рис. 3.6.23, 3.6.28).
Плоские карликовые клетки самые маленькие. Дендриты клеток, имеющие вид пучка, проникают в «триаду» «ножек» колбочек. Апикальный дендрит экстрафовеолярных карликовых биполярных клеток делится на две части. При этом он образует синапсы с двумя различными колбочками. Аксоны переходят через внутренний плексиформный слой и образуют синапсы с отростками амакриновых клеток и дендритами «карликовых» ганглиозных клеток (рис. 3.6.28). В области центральной ямки одна карликовая биполярная клетка контактирует с одной колбочкой. Эти биполярные клетки участвуют в образовании OFF-центр «рецептивных полей» колбочковой системы.
Плоские диффузные и инвагинированные «карликовые» биполярные клетки обладают многочисленными дендритами, заканчивающимися на «ножках» многих колбочек. Апикальный дендрит этих клеток разветвляется в наружном плексиформном слое, распространяясь в горизонтальной плоскости. Кроме того, эти биполярные клетки формируют обширную сеть в перифовеолярной области. Инвагнированные «карликовые» биполярные клетки участвуют в формировании ON-центр «рецептивных полей» колбочковой системы.
Биполярные клетки «синих» колбочек образуют синапсы более чем с одной «ножкой» колбочек. Биполярные клетки «синих» колбочек чаще встречаются в 4 мм от желтого пятна, а их аксональные терминалы простираются до 30 мкм. Эти клетки имеют также два мощных дендрита, которые заканчиваются на той же самой колбочке или на другой колбочке или в нейропиле наружного плексиформного слоя.
Различают два типа гигантских биполярных клеток. Это деление определяется протяженностью дендритов клеток. В центральных участках сетчатки длина дендритов равна 50 мкм, а по периферии 100 мкм. Биполярная клетка подобного типа объединяет 15—20 колбочек.
Гигантская диффузная биполярная клетка имеет толстый дендрит, который делится на три длинные ветви, а также раздваивающийся аксон. Аксон заканчивается в 4-м слое внутреннего плексиформного слоя. Если бы не размер дендритного поля, гигантские диффузные биполярные клетки имели бы строение аналогичное строению плоской диффузной биполярной клетки.
Все типы биполярных клеток близки по ультраструктурной организации. Ядро их круглое или овальное с одним или двумя ядрышками. Аппарат Гольджи, как и центриоли, располагается на участке выхода большого дендрита. Выполняют цитоплазму также рибосомы, шероховатый эндоплазматический ретикулум, митохондрии. В дендритах (толщина 0,1—0,2 мкм) также обнаруживаются митохондрии и микротрубочки, пузырьки и микрофиламенты (диаметр 20 нм). Аксональный бугорок расположен напротив выхода дендрита. В аксонах биполярных клеток выявляются и нейротрубочки (диаметр 12,5 нм). До внутреннего плексиформного слоя аксоны окружены отростками мюллеровских клеток.
После потери глиальной оболочки аксон образует утолщение (телодендрон), содержащее большое количество синаптических пузырьков, особенно вокруг синаптической ленты. Эфферентный или постсинаптический телодендрон обладает обычными синапсами, в то время как эфферентные отростки, образующие пресинаптический контакт с амакриновыми и ганглиозными клетками, обладают типичными ленточными синапсами.
Необходимо указать и на то, что основная часть внутреннего промежуточного слоя внутреннего ядерного слоя занята телами мюллеровских клеток, хотя они могут быть обнаружены и в любом другом слое сетчатки. Более подробно строение мюллеровской клетки изложено ниже.
Амакриновые клетки (рис. 3.6.29—3.6.31).

Рис. 3.6.29. Особенности распределения отростков амакриновых клеток во внутреннем плексиформном слое (объяснение в тексте)
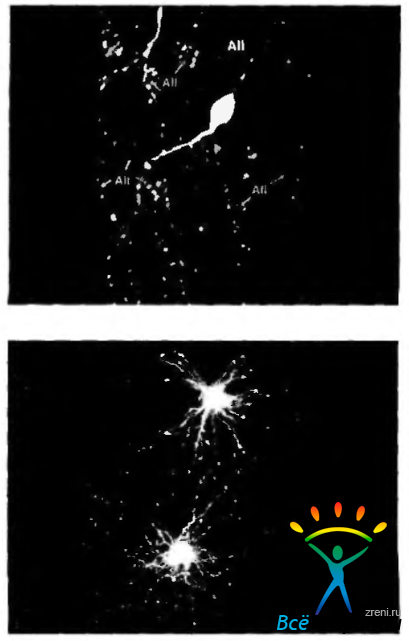
Рис. 3.6.30. Амакриновые клетки сетчатки, дающие положительную иммунногистохимичсскую реакцию, выявляющую серотонин (а) и допамин (б) (по Kolb, 1995)
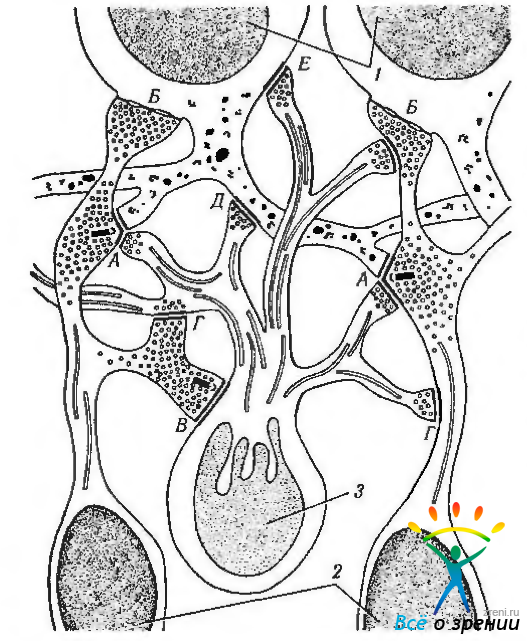
Рис. 3.6.31. Схематическое изображение синаптических контактов между биполярными, амакриновыми и ганглиозными клетками во внутреннем плексиформном слое (по Hogan et al., 1971): 1 - ганглиозные клетки; 2 - биполярная клетка; 3 - амакриновая клетка; А — аксодендритное окончание в диаде; Б — аксосоматическое окончание на ганглиозной клетке; В — контакт между амакриновой и биполярной клетками; Г—аксоаксонный контакт между отростками амакриновой и биполярной клетками; Д — аксодендритный контакт между амакриновой и ганглиозной клетками; Е—аксосоматический контакт между отростком амакриновой клетки и ганглиозной клеткой
Амакриновые клетки представляют собой нейроны, которые взаимодействует на втором уровне вертикального пути передачи зрительной информации, а именно в направлении: фоторецептор — биполярная клетка — ганглиозная клетка. Они формируют синапсы во внутреннем плексиформном слое. Эти клетки объединяют, а затем первично обрабатывают поступающую от биполярных клеток информацию и передают ее ганглиозным клеткам. Тела амакриновых клеток находятся несколько кнутри от ядер клеток Мюллера. Каждая амакриновая клетка имеет единственный отросток, обладающий свойствами дендрита и аксона. Отростки распространяются в обширной области во внутреннем плексиформном слое.
Тело амакриновой клетки имеет формул колбы диаметром 12 мкм. Располагаются они во внутреннем ядерном слое за исключением области желтого пятна. Цитоплазма содержит многочисленные митохондрии, шероховатую эндоплазматическую сеть (вещество Ниссля) и множество липидных включений. На внутренней поверхности клетки недалеко от ядра расположена ресничка.
В сетчатке человека амакриновые клетки отличаются разнообразным строением, и их описано 24 типа. При импрегнации сетчатки серебром по Гольджи выделяют два главных типа клеток: 1) диффузные и 2) стратифицированные.
Главный отросток клеток диффузного типа распространяется через все слои внутреннего плексиформного слоя. На его внутренней поверхности отросток разветвляется, формируя плотное горизонтально расположенное сплетение. В зависимости от протяженности отростков диффузные амакриновые клетки подразделяются на «узкопольные», охватывающие область шириной 10—50 мкм (составляет в среднем 25 мкм) и «широкопольные». Последние клетки во внутреннем плескиформном слое распространяются на 30—50 мкм, а в слое ганглиозных клеток до 600 мкм.
«Широкопольные» диффузные амакриновые клетки вступают в контакт с терминалами биполярных клеток палочек и ганглиозных клеток.
В зависимости от уровня расположения отростков во внутреннем плексиформном слое амакриновые клетки можно подразделить на следующие типы:
- нестратифицированные,
- мультистратифицированные
- и диффузные.
Отростки мультистратифицированных клеток разделяются на ветви, простирающиеся на расстояние до 400—600 мкм. При этом они занимают два или более уровней во внутреннем плексиформном слое. Ядра стратифицированных диффузных клеток меньше, чем ядра других амакриновых клеток, а их отростки охватывают область, шириной не более 50 мкм.
Амакриновые клетки также можно классифицировать по обнаруживаемому в них типу нейромедиаторов. Нейромедиаторами этих клеток являются нейроактивные вещества (ацетилхолин, гамма-аминомасляная кислота (ГАМК), глицин, допамин, серотонин) и нейропептиды (холецистокинин, энкефалин, глюкагон, нейротензин. соматостатин, вещество Р, нейропептид Y и вазоактивный кишечный пептид). В одной клетке могут присутствовать два или более перечисленных медиатора. Большинство амакриновых клеток содержат ГАМК, глицин, серотонин и допамин (рис. 3.6.30).
Физиологическое значение амакриновых клеток интенсивно изучается в последние годы. Именно благодаря одновременному использованию морфологических, иммуногистохимических и электрофизиологических методов исследования выявлен ряд функций этих клеток. Получены эти данные в экспериментах на животных, в частности на кошках.
У кошек различают несколько типов амакриновых клеток, функции которых достаточно хорошо изучены. Это амакриновая клетка А2, АН, А8, А13, А17, А19, А20, А22 и др. Об их роли будет рассказано в главе 4, посвященной зрительному пути.
Meжплексиформные клетки. Межплексиформные клетки описаны Gallego в 1971 г. Ядра межплексиформных клеток занимают самую внутреннюю часть внутреннего ядерного слоя. Поскольку тела клеток располагаются между амакриновыми клетками, некоторые авторы не выделяют этот тип клеток. Тем не менее отростки межплексиформных клеток, в отличие от амакриновых, простираются на плексиформные слои. Отростки к наружному плексиформному слою отходят непосредственно от тела межплексиформнои клетки или от их отростков. Сенсорный вход к межплексиформную клетку обнаруживается во внутреннем плексиформном слое, а большинство синапсов располагается в наружном плексиформном слое. Таким образом, информация передается между двумя указанными слоями. Межплексиформные клетки по своей природе относятся к центрифугальным нейронам. В сетчатке человека синапсы обнаруживаются, в основном, между межплексиформными клетками и горизонтальными клетками колбочек, получая при этом информацию от отростков амакриновой клетки. В зависимости от используемого нейромедиатора межплексиформные клетки позвоночных разделяют на три типа. Это клетки, использующие ГАМК, тирозин гидроксилазу и глицин. У человека выявлен только 1 тип клеток (ГАМК).
Внутренний сетчатый (плексиформный) слой (рис. 3.6.1, 3.6.31). Во внутреннем плексиформном слое контактируют второй (биполярная клетка) и третий (ганглиозная клетка) нейроны сетчатки. В пределах этого же слоя также взаимодействуют амакриновые и межплексиформные клетки. Кроме синаптических связей между биполярными, ганглиозными, амакриновыми и межплексиформными клетками этот слой содержит отростки мюллеровских клеток, а также обильную сосудистую сеть и ядра единичных ганглиозных и амакриновых клеток.
Внутренний плексиформный слой толще наружного плексиформного слоя. Отсутствует он только в области желтого пятна.
Во внутреннем плексиформном слое видны многочисленные синапсы, плотность которых достигает 2 млн в мм2.
Существуют определенные ультраструктурные особенности этого слоя сетчатки. Именно в этом слое биполярные клетки вступают в синаптический контакт с отростками амакриновых клеток и дендритами ганглиозных клеток, образуй так называемую «диаду». Наиболее часто один из элементов «диады» представляет собой дендрит ганглиозной клетки, а другой — отросток амакриновой клетки. Подобный тип синаптичекой организации соответствует наличию в этой области так называемых ганглиозных клеток «контрастности».
Реже в «диаде» обнаруживается два отростка амакриновой клетки или, что более редко, два дендрита ганглиозной клетки (рис. 3.6.31). Отростки амакриновых клеток связываются с аксонами биполярной клетки, телами и дендритами ганглиозных клеток посредством синапсов обычного строения. Межплексиформные клетки также образуют обычные синапсы, главным образом, с отростками амакриновых клеток.
В этом слое существует два уникальных типа синапсов, свойственных только амакриновым клеткам. Это «реципроктный» и «последовательный» синапсы. В «реципроктном» синапсе отросток амакриновой клетки в «диаде» образует синапс с терминалом биполярной клетки, обеспечивая, таким образом, обратную связь между амакриновой и биполярными клетками около синаптической ленты. «Последовательный синапс» состоит из двух последовательно расположенных синапсов между двумя отростками амакриновых клеток, а третий синапс образуется с дендритом ганглиозной клетки, аксоном биполярной клетки или другим отростком амакриновой клетки. Эта сеть обеспечивает взаимодействие соседних амакриновых клеток.
Синапсы амакриновых клеток располагаются слоями. Так, в области желтого пятна обнаруживается только два слоя синапсов, а по периферии число их слоев достигает пяти.
Слой ганглиозных клеток (рис. 3.6.1). Слой ганглиозных клеток состоит в основном из тел ганглиозных клеток. В этом слое обнаруживаются также отростки мюллеровских клеток, нейроглия и сосуды сетчатой оболочки. Ганглиозные клетки получают обработанные зрительные сигналы от предшествующих двух нейронов, обрабатывают их и передают в центральную нервную систему.
Ганглиозные клетки по периферии сетчатки образуют один слой клеток. С височной стороны диска зрительного нерва выявляется 2 слоя клеток, а по краям желтого пятна 6—8 слоев. В центре желтого пятна и диске зрительного нерва ганглиозные клетки отсутствуют.
Толщина слоя ганглиозных клеток колеблется от 10 до 20 мкм в назальной части сетчатки до 60—80 мкм в области желтого пятна.
В сетчатке взрослого определяется от 0,7 до 1,5 млн ганглиозных клеток. Соседние ганглиозные клетки плотно прилегают друг к другу за исключением периферии сетчатки. Здесь расстояние между ними достигает 400 мкм. Каждая клетка имеет один аксон. Собираясь на внутренней поверхности сетчатки, аксоны покидают глазное яблоко и формируют зрительный нерв.
В кольце, опоясывающем желтое пятно сетчатки, которое находится на расстоянии 0,4 — 2,0 мм от пятна, плотность ганглиозных клеток колеблется от 32 000 до 38 000 клеток в мм2. По периферии плотность ганглиозных клеток в назальном квадранте в три разе превышает таковую в темпоральном квадранте. Плотность клеток в верхнем квадранте превышает плотность клеток в нижнем квадранте на 60%. Отношение количества колбочек к количеству ганглиозных клеток колеблется от 2,9:1 до 7,5:1.
В слое ганглиозных клеток обнаруживается до 3% амакриновых клеток в центральных областях сетчатки, по периферии почти 80%. Ганглиозные клетки больших размеров (диаметр от 10 до 30 мкм), круглой или овальной формы. В области желтого пятна размер клеток несколько меньше.
В цитоплазме развита шероховатая эндоплазматическая сеть (вещество Ниссля) и аппарат Гольджи (рис. 3.6.32).

Рис. 3.6.32. Ультраструктурные особенности ганглиозной клетки (по Hogan, 1966): отмечается хорошее развитие шероховатой эндоплазматической сети (стрелки). Цитоплазма насыщена овальными пигментированными частицами, придающими сетчатке желтоватый цвет
Обнаруживаются также диффузно распределенные фрагменты гладкой эндоплазматической сети, митохондрии, капельки липидов и пигментные гранулы. С возрастом отмечается увеличение количества зерен липофусцина. С этим связывают усиление желтизны макулярной области.
Ганглиозные клетки обладают многочисленными нейрофиламентами, что позволяет легко отличать ганглиозные клетки от мюллеровских.
Ганглиозные клетки относятся к мультиполярным нейронам. Их дендриты распределяются в горизонтальной плоскости сетчатки, а также проникают во внутренний плексиформный слой. Их аксоны направляются к слою нервных волокон, где они ориентируются параллельно внутренней поверхности сетчатки (рис. 3.6.33).

Рис. 3.6.33. Особенности строения тела клеток и их дендритного поля в различных участках сетчатой оболочки (по Polak, 1940): а — область центральной ямки; б — область экватора; в — периферия
Ганглиозные клетки классифицируют в соответствии с размером тел клеток, степенью развития отростков и их протяженности. Классифицируют их также по типу синаптической связи с амакриновыми и биполярными клетками.
В последние годы описано приблизительно 18 различных морфологических типов ганглиозных клеток. Пока не совсем ясно, являются они только морфологической разновидностью основного типа клеток или различны и их функции.
В глазном яблоке человека идентифицировано два основных типа клеток, обозначенных как клетки M (зонтикоподобные) и клетки Р. В свою очередь клетки Р подразделяются на два подкласса: Р1, или карликовые нейроны, и Р2 (рис. 3.6.34).
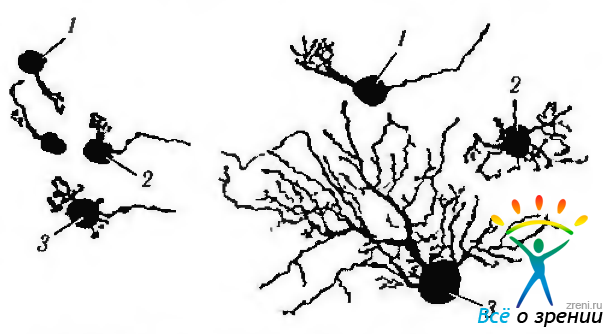
Рис. 3.6.34. Особенности строения ганглиозных клеток сетчатки человека, определяющих функционирование Р- и М-трактов зрительного анализатора: 1—карликовая Р1; 2— маленькая зонгикоподобная Р2; 3 — большая зонтикоподобная М
М-клетки проецируются на магноцеллюлярные (крупноклеточные) слои наружного коленчатого тела и определяют так называемый «не-оппонентный» ответ (см. главу 4). По своим физиологическим характеристикам, М-клетки человека напоминают ганглиозные клетки обезьяны, специализированные на восприятии цвета.
Р-клетки проецируются на парвоцеллюлярные (мелкоклеточные) слои наружного коленчатого тела. Клетки типа Р1 самые маленькие и обладают небольшим дендритным деревом.
Эти клетки определяют «оппонентный» ответ при стимуляции сетчатки средне- и коротковолновой частями спектра. Каждая Р-клетка получает информацию только от одной колбочки. Ганглиозные клетки типа Р1 подразделяют на так называемые широковетвистые (а-тип) и слабоветвистые (b-тип). Первые участвуют в формировании рецептивных полей с OFF-центрами, а вторые — с ON-центрами.
В области центральной ямки клетки типа Р1 составляют 90% общего количества ганглиозных клеток. В этой области сетчатки их удлиненные тела имеют размеры 8x12 мкм и достигают максимума (14x16 мкм) на расстоянии 8 мм от фовеа. Здесь они составляют 40—45% от общего количества ганглиозных клеток.
Единственный дендрит клетки формирует небольшое количество терминалов (5—7 нм в диаметре). Они переходят во внешнюю (а-тип) или внутреннюю (b-тип) треть внутреннего плексиформного слоя сетчатки. Синапсы а-типа образуются между аксонами плоских диффузных биполярных клеток, а синапсы b-типа — с инвагинирующими биполярными клетками.
В терминалах типа «диад» или «монад» присутствует до 55—81 лент. Количество отростков амакриновых клеток, которые образуют синапсы с дендритическим деревом этих ганглиозных клеток, приблизительно равно числу синаптических лент биполярной клетки. Распространяются дендриты ганглиозных клеток типа Р1 на 5—10 мкм в центральных областях сетчатки, а по периферии на 225 мкм.
Отличить ганглиозные клетки Р1 от Р2 в области центральной ямки практически невозможно. Однако в 1,5 мм от нее клетки типа Р2 значительно больших размеров. Дендритное поле клеток, расположенных на расстоянии 6—8 мм от центральной ямки простирается на 30—50 мкм, а лежащих по периферии сетчатки на 400 мкм.
Клетки типа Р2 проявляют выраженный ответ при стимуляции светом S-колбочек [229]. Они составляют 1 % общего количества ганглиозных клеток в области фовеа и 10% по периферии сетчатой оболочки.
Размер М-ганглиозных клеток больше, чем клеток типа Р. Больше и их дендритные поля (25—30 мкм). Причем дендритное поле увеличивается по мере продвижения к периферии сетчатки. Так, в 8 мм от центральной ямки дендритное поле равняется 160 мкм, а на расстоянии 14 мм — 270 мкм.
М-клетки составляет 5% общего количества ганглиозных клеток в области центральной ямки и 20% по периферии сетчатки.
В литературе сейчас активно обсуждается вопрос о гибели ганглиозных нейронов сетчатки при глаукоматозном процессе, именуемом как глаукоматозная нейропатия. Само это состояние рассматривается как многолетний хронический процесс с постепенной медленной потерей отдельных ганглиозных клеток или их небольших групп при сохранении морфологии и функции других. Предполагалось, что в этом процессе преимущественно погибают магноцеллюлярные М-нейроны. Однако Morgan et al. (2000) обнаружили, что в сетчатке обезьян с экспериментальной гипертензией в одинаковом соотношении погибают и магно- и парвоцеллюлярные нейроны. При этом клетки сморщиваются, так что объем М-нейронов достоверно уменьшается на 20%, а Р-нейронов — на 16%.
Слой нервных волокон (рис. 3.6.1). Слой нервных волокон образуется аксонами ганглиозных клеток (так называемые «центростремительные», или «приводящие» волокна), а также глиальными элементами, большим количеством капиллярных сосудов и центрифугальных (эфферентных) волокон.
Аксоны ганглиозных клеток образуют дуги, очерченные отростками мюллеровских и других глиальных клеток. Отдельные афферентные волокна имеют диаметр от 0,6 мкм до 2,0 мкм. Они содержат микротрубочки, митохондрии и гладкую эндоплазматическую сеть. В них происходит двухсторонний аксоплазматический поток двух типов — медленный и быстрый. Медленный поток (0,5—5 лш/день) несет высокомолекулярные белки, используемые для роста аксонов и их физиологической регенерации. Быстрый поток (10—2000 лш/день) обеспечивает функционирование синапсов путем поставки питательных веществ.
Механизмы, обеспечивающие аксонный транспорт, изучаются до сих пор. В соответствии с одной из теорий движение в направлении аксона обеспечивается движением цитоплазмы. Другие авторы считают, что в этом процессе основную роль играют микротрубочки. В подтверждение правильности последнего предположения приводится факт прекращения транспорта после обработки клеток колхицином, разрушающим микротрубочки.
Аксоны ганглиозных клеток сетчатки остаются немиелинизированными до момента достижения ими решетчатой пластинки.
Аффрентные волокна радиально продвигаются параллельно внутренней пограничной мембране и сходятся в области диска зрительного нерва. Исключением являются аксоны, исходящие из ганглиозных клеток, расположенных непосредственно с височной стороны диска зрительного нерва. Волокна папилло-макулярного пучка распространяются дугообразно. Верхние и нижние волокна отделены горизонтальным «швом», простирающимся от желтого пятна до крайней периферии сетчатки.
Наиболее толстым является слой нервных волокон у края диска зрительного нерва с назальной стороны (20—30 мкм). Толщина его уменьшается по мере приближения к зубчатой линии. Значительно точнее варианты изменения толщины слоя нервных волокон можно выявить при помощи лазерной офтальмоскопии. Эти данные важны при установлении диагноза ряда заболеваний глаза, в частности глаукомы.
Папилло-макулярный пучок является наиболее тонкой частью слоя нервных волокон, расположенного вокруг диска зрительного нерва. Поскольку наибольшее количество аксонов собирается с назальной стороны диска зрительного нерва, они образуют возвышенность (сосок), выстоящую в стекловидное тело.
Центрифугальные волокна, берущие свое начало в центральной нервной системе, заканчиваются во внутреннем плексиформном слое или самой внутренней части внутреннего ядерного слоя. Обычно они образуют синапс с амакриновыми клетками или стенками капиллярных сосудов. В последне

Комментариев 0