Сосуды и сосудистая оболочка глазного яблока │ Часть 1


Содержание:
Описание
Увеальный тракт (tractus uvealis) глазного яблока состоит из радужной оболочки, ресничного тела (цилиарное тело) и сосудистой оболочки (хориоидея). Увеальный тракт легко отделяется от склеры. Сформирован он многочисленными сосудами — артериями и венами. В свою очередь артерии увеального тракта берут свое начало из ресничных артерий, подходящих к глазному яблоку. Вены увеального тракта впадают в вортикозные вены, отводящие кровь от глаза в вены глазницы. Перед тем как остановиться на строении увеального тракта, необходимо описать сосудистую систему, кровоснабжающую глазное яблоко.↑ Артерии и вены глазного яблока
Артериальная система глаза разделяется на две части: задние и передние ресничные артерии.
Различают следующие артерии (рис. 3.8.1, 3.8.2, 3.8.3, 3.8.4):
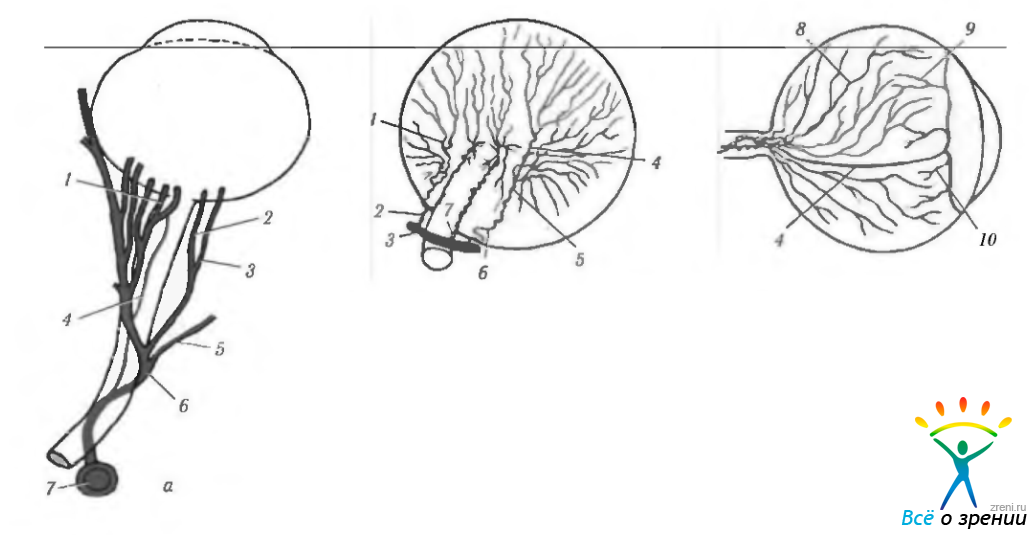
Рис. 3.8.1. Схема распределения сосудов, кровоснабжающих глазное яблоко: а—распределение сосудов в глазнице (1—задняя короткая ресничная артерия: 2—внутренняя длинная ресничная артерия; 3 — наружная длинная ресничная артерия: 4 — центральная артерия сетчатки; 5—слезная артерия; 6—глазная артерия; 7—внутренняя сонная артерия); б—распределение ресничных артерий (вид сзади) (1—дистальная назальная короткая ресничная артерия; 2— задняя ресничная назальная артерия; 3— глазная артерия; 4 — дистальная темпоральная длинная ресничная артерия; 5—дистальная темпоральная короткая ресничная артерия; 6—дистальная задняя ресничная артерия; 7 — параоптическая задняя ресничная артерия); в—распределение ресничных артерий (вид сбоку) (8— задняя короткая ресничная артерия; 9— передняя возвратная ресничная артерия; 10 — возвратные ветви задней длинной ресничной артерии)
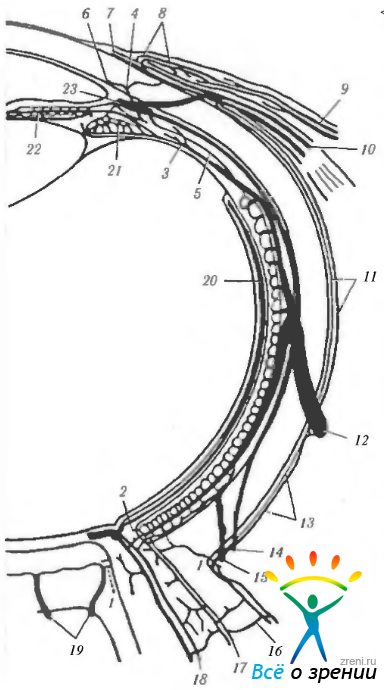
Рис. 3.8.2. Кровоснабжение глазного яблока (по Mever, 1989): 1 — ветвь короткой задней ресничной артерии к зрительному нерву: 2 — анастомозы между сосудами хориоидеи и центральной артерией сетчатки: 3 — вена, идущая от ресничной мышцы к вортикозной вене; 4 — ветвь передней ресничной вены, идущей от ресничной мышцы; 5 — возвратная артерия; 6 — шлеммов канал; 7 — лимбальное сосудистое сплетение; 8- передние сосуды конъюнктивы; 9 — задние сосуды конъюнктивы; 10—передние ресничные артерии; 11 — эписклеральные артерии; 12—вортикозная вена; 13—эписклеральные сосуды; 14— задняя длинная ресничная артерия; 15 — задние короткие ресничные артерии; 16 сосуды твердой оболочки; 1~ сосуды паутинной оболочки; 18—центральная артерия и вена; 19— задние ресничные артерии; 20 — хориокапилляры; 21 — ресничные отростки; 22 — сосуды радужки; 23 — большой круг кровообращения радужки

Рис. 3.8.3. Схема кровоснабжения переднего отдела глаза (no Mover. 1989): артериальное кровообращение переднего отдела глаза обеспечивается поверхностным и глубоким артериальным кругом (эписклеральный артериальный круг, большой круг кровообращения радужки и внутримышечный артериальный круг ресничного тела). Они получают кровь из сагиттальных артериальных кругов (длинные задние ресничные артерии, мышечные и передние ресничные артерии и перфорирующих ветвей этих систем) (1 — глазная артерия; 2 — задняя длинная ресничная артерия; 3 — артерия прямой мышцы; 4 — эписклеральные капилляры; 5 — капилляры прямой мышцы; 6 — передняя ресничная артерия)
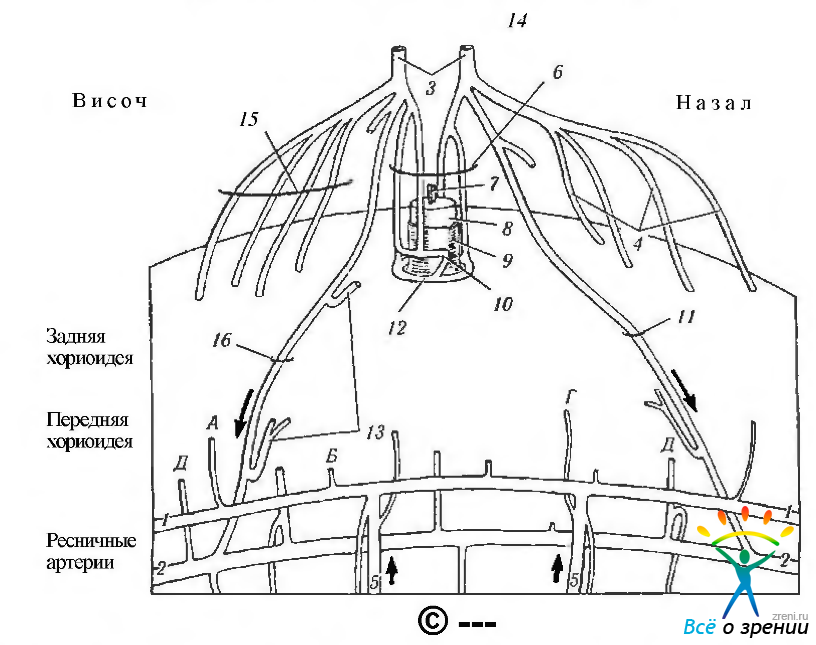
Рис. 3.8.4. Схематическое изображение распределения ресничных артерий (по Bron el al., 1997): 1 — внутримышечный ресничный артериальный круг; 2— большой артериальный круг радужки; 3— задняя ресничная артерия; 4 - короткие задние ресничные артерии; 5 -передние ресничные артерии; А—передняя хориоидальная ветвь ресничного внутримышечного круга; 6 — параоптические короткие задние ресничные артерии; 7 — сосуды сетчатки; 8 — мягкая оболочка; 9 — твердая оболочка; 10— проксимальный круг; 11 — задняя длинная ресничная артерия; 12— Kpyг Цинна; 13— возвратные ветви задней длинной ресничной артерии; 14 — глазная артерия; 15 — задние ресничные артерии; Б — ветвь внутримышечной ресничной артерии; В — артерия радужки, исходящая из ресничной ветви большого круга кровообращения радужки; Г—ветвь к хориоидее, исходящая из передней ресничной артерии; Д — ветвь к переднему отделу хориоидеи, исходящая из большого круга кровообращения
- Задние медиальная (назальная) и латеральная (темпоральная) ресничные артерии.
- Задние короткие ресничные артерии (аа. ciliaris brevis posterior).
- Задние длинные ресничные артерии (аа. ciliaris longi posterior).
- Передние ресничные артерии (аа. ciliaris anterior).
Задние медиальная и латеральная ресничные артерии. Одна медиальная и одна латеральная задние ресничные артерии отделяются от глазной артерии. Это происходит в месте прохождения глазной артерии над зрительным нервом. Иногда обнаруживается верхняя задняя ресничная артерия (рис. 3.8.1).
Медиальная и латеральная артерии делятся на 10—20 ветвей, а затем направляются вперед, окружая зрительный нерв. Проникают они в глазное яблоко, прободая склеру вокруг зрительного нерва. При этом они разделяются на короткие и длинные артерии (рис. 3.8.2). Больше всего образуется коротких ресничных артерий. Одна медиальная и одна латеральная ветви превращаются в длинные задние ресничные артерии.
Задние короткие ресничные артерии (аа. ciliaris brevis posterior). Большинство задних коротких ресничных артерий, после того как они отдают ветви склере, прободают склеру с височной стороны зрительного нерва в области проекции желтого пятна. Небольшое количество сосудов меньшего калибра прободают склеру по окружности зрительного нерва, но ближе к нему. Склеральные каналы, через которые проходят артерии, короткие и направлены кпереди. Располагающиеся в склеральном канале сосуды окружены тканью, аналогичной ткани супрахориоидеи.
Задние короткие ресничные артерии ди- и трихотомически делятся недалеко от диска зрительного нерва. При этом образуется большое количество дистальных ветвей и значительно меньше параоптических, располагающихся ближе к диску зрительного нерва. С височной стороны задних коротких ресничных артерий больше.
Маленькие параоптические артерии участвуют в формировании сосудов перипапиллярной части сосудистой оболочки, а также вертикальной трапециевидной полосы сосудистой оболочки, расположенной выше и ниже диска зрительного нерва. Нередко в формировании этой сети участвуют ветви круга Цинна—Халлера, (более подробно сведения о круге Цинна—Халлера приведены в разделе «Зрительный нерв»).
Дистальные задние короткие ресничные артерии снабжают большие области сосудистой оболочки треугольной формы, чьи вершины расположены приблизительно в месте входа каждого дистального пучка сосудов. Один из этих пучков располагается с назальной, а другой с темпоральной стороны.
Задние короткие ресничные артерии проходят в наружном слое сосудистой оболочки (возвратные хориоидальные ветви), а от них отходят артериолы, формирующие промежуточный слой (слой Саттлера) (рис. 3.8.3). Перипапиллярно несколько ветвей хориоидальных артериол пересекают край диска и кровоснабжают преламинарную часть зрительного нерва. Кроме того, возвратные ветви кровоснабжают пиальную оболочку.
Задние длинные ресничные артерии (a. ciliares posteriores longae) (рис. 3.8.1—3.8.4). Медиальная и латеральная задние длинные ресничные артерии прободают склеру с обеих сторон зрительного нерва. Это происходит несколько кпереди от места проникновения коротких ресничных артерий. Затем они под косым углом проходят в склеральном канале (длина 4 мм). В последующем, изгибаясь кнутри под углом 45°, проникают внутрь глаза.
Отверстие склерального канала довольно широкое, так что в канале, помимо ресничных артерий, проходят нервы, а также вены. Свободное от перечисленных образований пространство каналов выполнено соединительной тканью.
Артерии достигают супрахориоидального пространства и направляются вперед в горизонтальной плоскости. Их ход можно наблюдать через конъюнктиву. Выглядят они в виде просвечивающихся синих линий.
В передней части сосудистой оболочки (или иногда в пределах ресничной мышцы) задние длинные ресничные артерии разделяются и формируют большой артериальный крут радужки (рис. 3.8.3, 3.8.4, 3.8.17).

Рис. 3.8.17. Меридианальный срез глазного яблока. Схематическое изображение кровоснабжения области лимба (красным цветом обозначены артерии): ПРА — передняя ресничная артерия, разделяющаяся на эписклеральные (Эп) и большие перфорирующие ветви (БПВ). Эписклеральные ветви дают начало эписклеральным, конъюнктивальным (Кв) и интрасклеральным (ИСк) ветвям. Эписклеральные ветви формируют поверхностное краевое сплетение роговицы (ПКСР). Две системы сосудов образуются из этого сплетения: первое (1) — распространяется вперед с образованием периферических роговичных дуг. а второе возвратные артерии (2), направляющиеся кзади и кровоснабжающие полоску конъюнктивы, шириной от 3 до 6 мм. Позже они образуют анастомозы в области свода конъюнктивы. Большие перфорирующие артерии (3) проходят сквозь склеру, соединяясь с большим кругом кровообращения радужки (БКрР). Ветви большой перфорирующей артерии проходят кпереди, формируя интрасклеральные сосуды в области лимба. Эта область часто кровоснабжается сосудами, исходящими из передней ресничной артерии в виде эписклеральных артерий (4). Вены окрашены в синий цвет. Венозный дренаж области лимба проходит в эписклеральные вены, которые затем впадают в глазные вены. Глубокое венозное склеральное сплетение (5) присоединяется к шлеммову каналу (ШК). Водяные вены (стрелки) начинаются в глубоком склеральном сплетении и соединяются с эписклеральными венами. Интрасклеральное венозное сплетение (6) образует выраженную сеть сосудов в строме лимбальной области. Важным путем дренажа венозной крови из ресничного сплетения (РСп) являются глубокое и интрасклеральное венозные сплетения. Один из подобных каналов обозначен цифрой 7.
Глубокие ветви передних ресничных артерий не вносят существенного вклада в формирование сосудистой сети большого круга кровообращения радужки. Они участвуют в образовании внутримышечного артериального круга, описанного еще Лебером в 1903 г. и расположенного в толще ресничной мышцы. Во всех плоскостях обнаруживаются анастомозы между ветвями передних ресничных и задних длинных ресничных артерий. Ответвления передних реcничных артерий кровоснабжают периферию сосудистой оболочки, образуя многочисленные «возвратные» артерии. Эти артерии кровоснабжают также шлеммов канал и лимбальную область.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Передние ресничные артерии (рис. 3.8.2— 3.8.4). Передние ресничные артерии отделяются от артерий четырех наружных прямых мышц глаза. Из каждой мышцы обычно выходит по две артерии. Исключением является наружная прямая мышца. От нее отделяется только одна артерия.
На расстоянии приблизительно в 1,5 мм от лимба эти артерии разделяются на глубокие (склеральные) и поверхностные (передние эписклеральные) ветви. Артерии проникают внутрь глаза через короткие склеральные каналы и распространяются в ресничной мышце, соединяясь с «внутримышечным артериальным крутом». При этом они отдают ветви к радужке и «возвратным» артериям хориоидеи. Места вхождения артерий в склеру нередко пигментированы.
«Передние эписклеральные артерии» направляются вперед и формируют «эписклеральный артериальный круг», анастомозирущий с глубокими артериями.
«Эписклеральный артериальный крут» поставляет кровь склере, лимбу и перилимбальной конъюнктиве. Посредством глубоких ветвей. направляющихся к большому кругу кровообращения радужки, они кровоснабжают и радужку.
Необходимо отметить, что, поскольку передние ресничные артерии кровоснабжают ресничную мышцу, радужку и эписклеру, становится понятным, почему при воспалении радужки или ресничного тела (передний увеит) эписклеральные сосуды лимбальной области резко расширяются и переполняются кровью.
О важном значении передних ресничных артерий в кровообращении переднего отдела глаза указывают и клинические наблюдения. Так, повреждение сухожилий прямых мышц глаза при проведении хирургического лечения косоглазия приводит к нарушению кровообращения ресничной мышцы (и радужки), получающей кровь из бассейна передней ресничной артерии. Это вызывает ишемию переднего отдела глаза. Аналогичная ситуация возникает при проведении циркляжа по Аруга в процессе лечения отслойки сетчатки. При этом сдавление артерий и вен является причиной возникновения «переднего ишемического синдрома».
Вены. Вортикозные вены отводят венозную кровь практически от всего увеального тракта. Передние ресничные вены дренируют часть ресничной мышцы. Эти две венозные сети связаны между собой, что имеет большое компенсаторное значение при различных заболеваниях, сопровождающихся нарушением кровообращения. Если отток венозной крови через вортикозные вены нарушается, передние ресничные вены берут на себя их функцию.
Вены вортикозные (v. vorticosae: v. chorioideae oculi) (рис. 3.8.2, 3.8.4). Обычно обнаруживаются четыре вены (две верхние и две нижние). Они выходят из глаза, прободая склеру под косым углом вблизи верхней и нижней прямых мышц в 6 мм позади экватора. Верхние вены покидают глазное яблоко несколько ближе к зрительному нерву, чем нижние. В то же время темпоральные вены выходят ближе к экваториальной плоскости, чем медиальные. Верхняя темпоральная вена выходит несколько кзади (8 мм позади экватора) и прилежит к сухожилию верхней косой мышцы. Нижняя темпоральная вена расположена несколько кпереди (5,5 мм позади экватора). Иногда, особенно при близорукости, вены покидают глазное яблоко далеко позади экватора, иногда вблизи зрительного нерва. Нередко обнаруживается более четырех вен.
Вортикозные вены проходят сквозь склеру в каналах, длина которых равняется приблизительно 4 мм. Расположение их в канале можно наблюдать невооруженным глазом (темные полосы).
В каналах вены нередко разделяются на ряд стволов. При этом на поверхность глазного яблока выходит 6 или больше сосудов. Стволы вортикозных вен перед проникновением в склеру ампулоподобно расширяются.
Вены хориоидеи объединяются и образуют вортикозные вены. Задние венозные ветви отводят кровь от заднего отдела сосудистой оболочки, диска зрительного нерва, а иногда от перипапиллярной сетчатки. Ветви, расположенные вокруг диска зрительного нерва, непосредственно впадают в вортикозные вены.
Передние вены, отводят кровь в вортикозные вены от радужной оболочки, ресничных отростков, ресничной мышцы и переднего отдела сосудистой оболочки. Вены в области плоской части ресничного тела располагаются параллельно друг другу. В области зубчатой линии они идут в направлении вортикозных вен.
Вены ресничных отростков проходят кзади в виде параллельно расположенных сосудов, анастомозирующих в области плоской части ресничного тела с венами, идущими от внутренней стороны ресничной мышцы. В последующем они проходят сосудистую оболочку и впадают в вортикозные вены.
Вены ресничной мышцы направляются назад и впадают в вены ресничных отростков.
Вены радужки проходят подобно артериям, анастомозируя друг с другом. Затем они поступают в ресничное тело и объединяются с венам ресничных отростков, впадая в последующем в вортикозные вены.
Многие исследователи считают, что у человека водораздел между четырьмя областями венозного дренажа образует так называемый «Мальтийский крест», проходящий через диск зрительного нерва. Форма «креста» изменяется в соответствии с количеством вортикозных вен.
Две верхние вортикозные вены открываются в верхнюю глазную вену непосредственно либо через мышечные или слезные венозные ветви. Две нижние вены открываются в подглазничную вену или в анастомоз с верхней глазной веной.
Склеральные вены соответствуют склеральным ветвям коротких ресничных артерий. Они отводят кровь только от склеры. По этой причине их калибр меньше, чем калибр артерий.
Передние ресничные вены, подобно артериям, образуют ветви мышечных вен. Поскольку они дренируют только ресничную мышцу, они меньше, чем соответствующие артерии.
↑ Радужная оболочка
Радужная оболочка (Iris) представляет собой диафрагму, разделяющую пространство между роговой оболочкой и хрусталиком (рис. 3.8.5).

Рис. 3.8.5. Расположение радужной оболочки относительно роговой оболочки, ресничного тела и хрусталика: 1 — радужная оболочка; 2 — роговая оболочка; 3 — хрусталик; 4 — ресничное тело; 5 — дренажная система; 6 — угол передней камеры
Это пространство выполнено водянистой влагой и разделяется на две камеры — переднюю и заднюю. Расположенное в центре радужки отверстие — зрачок, — увеличиваясь или уменьшаясь в диаметре, контролирует количество света, поступающего в глаз, обеспечивая тем самым глубину резкости. Уменьшение диаметра зрачка устраняет также сферическую и хроматическую аберрацию. Процесс изменения диаметра зрачка происходит благодаря деятельности специализированных мышц —сфинктера и дилятатора.
Диаметр радужки равен приблизительно 12 мм, а ее периметр 38 мм. У молодых людей диаметр зрачка изменяется в пределах 1,5 — 8,0 мм. В пожилом возрасте диаметр зрачка нередко меньше, в связи с развитием фиброзных изменений сфинктера и атрофии дилятатора. Зрачок может быть расширен более чем на 9 мм при использовании мидриатиков. Степень расширения зрачка ограничена у больных диабетом. Радужка толще в области «воротничка» (0,6 мм) и зрачкового края (рис. 3.8.6).
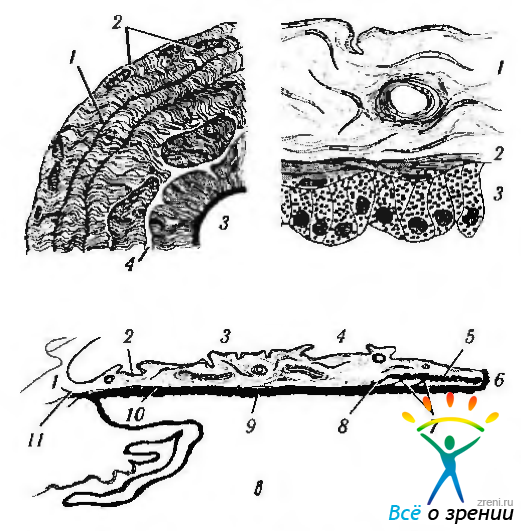
Рис. 3.8.6. Схематическое изображение особенностей анатомического строения радужной оболочки: а—передняя поверхность радужки (1 — цилиарные крипты; 2 - цилиарная зона; 3 - зрачковая зона; 4 крипта); б строение радужки в прикорневой области (большое увеличение) (1 — строма; 2 — дилятатор; 3 — пигментный эпителий); в — строение радужки и отношение ее к ресничному телу (1 — корень радужки; 2 — цилиарная крипта; 3 — борозда сокращения; 4 — крипта; 5 — сфинктер; б — зрачковый край; 7—шпора Фукса; 8—шпора Михельса; 9 — пигментый эпителий; 10 — дилятатор; 11 — шпора Грюнерта)
Истончается она к периферии (0,5 мм). Тонкая периферия радужки является наиболее частым местом травматических отрывов — иридодиализа. Наличие большого количества сосудов в этой области является причиной кровоизлияний в переднюю и заднюю камеры глаза в результате травмы.
Своей задней поверхностью зрачковый край радужки лежит на передней поверхности хрусталика. При воспалительных заболеваниях этот контакт может привести к прилипанию пигментных клеток радужки к капсуле хрусталика и формированию задних синехий.
При нарушении положения хрусталика (сублюксация, дислокация) или его удалении радужка отклоняется назад в плоскость корня радужки. В этих случаях передняя камера становится более глубокой и радужка дрожит при движении глаза (иридодонез).
Основную толщу радужной оболочки составляет строма, которая имеет мезенхимное происхождение. В анатомическом смысле некоторые авторы выделяют поверхностные и глубокие слои стромы.
Поверхностный (пограничный) cлой стромы более короткий и распространяется от корня радужки до так называемого «воротничка» (рис. 3.8.7). «Воротничок» виден на передней поверхности радужки в виде зубчатой линии, расположенной на определенном расстоянии от зрачкового края. Считают, что передний мезенхимальный слой представляет собой остатки сосудистой зрачковой мембраны, хорошо развитой в эмбриональном периоде. В связи с характером расположения сосудов в эмбриональном периоде поверхностный слой стромы имеет радиальное трабекулярное строение. Именно этот слой определяет степень пигментации радужки (рис. 3.8.7, 3.8.8).
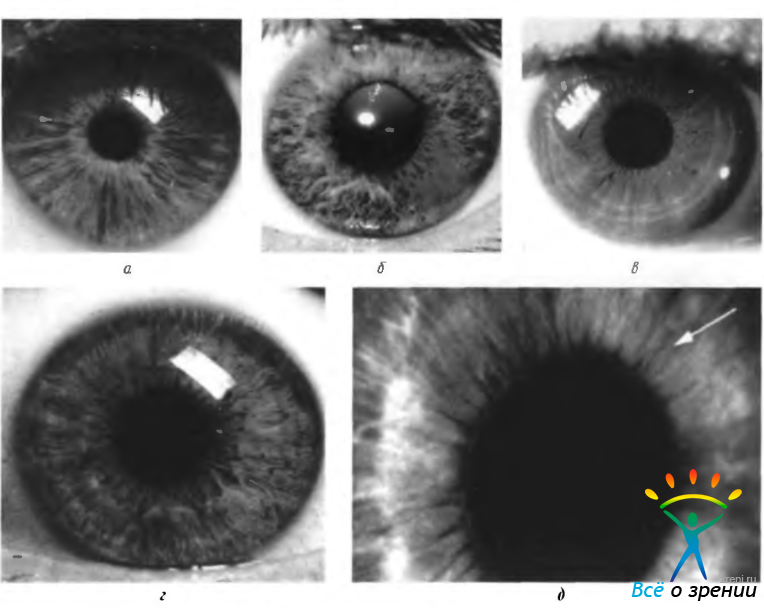
Рис. 3.8.7. Некоторые типы пигментации радужной оболочки: а — слабо пигментированная радужка. Радиальное расположение коллагеновых колонок; б- голубая радужная оболочка с сектором интенсивной пигментации; в — коричневая радужная оболочка в результате диффузной интенсивной пигментации стромы. По периферии видны сократительные борозды; г — пигментация переднего пограничного слоя, особенно интенсивная в области «воротничка»; д — слабо пигментированная радужка. Просматривается сфинктер (стрелка)
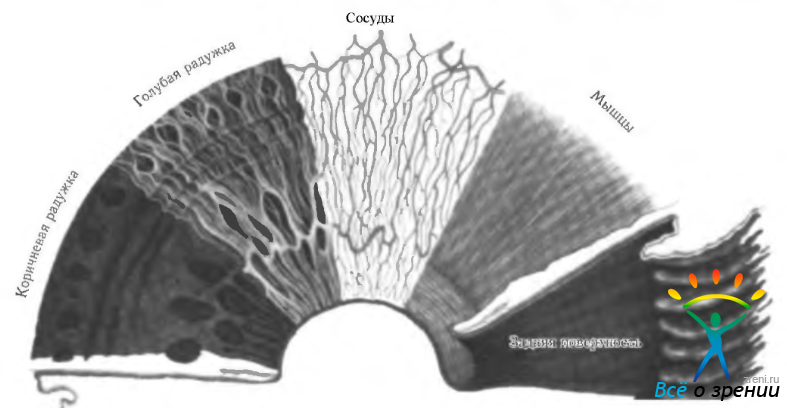
Рис. 3.8.8. Слои радужной оболочки (по Hog <ап et al., 1971)
Глубокий cлой стромы распространяется от корня радужки до зрачкового края. В светлых радужках глубокий слой имеет волокнистое строение с радиальным распространением пучков волокон. Этот слой слабо соединен с поверхностным слоем. По этой причине при сокращении радужной оболочки происходит как бы скольжение между ними, в результате чего «воротничок» приближается к зрачковому краю.
Цвет радужки обусловлен количественным содержанием в ее строме меланоцитов и является наследуемым признаком. Коричневая радужка наследуется доминантно, а голубая рецессивно. У большинства новорожденных радужная оболочка голубая, поскольку увеальный тракт в это время слабо пигментирован. На 3—6-м месяце жизни у многих радужка темнеет, поскольку число меланоцитов и степень их пигментации увеличивается. Если строма радужки бедна пигментированными клетками, радужка имеет голубой цвет (рис. 3.8.7, 3.8.8). У альбиносов отсутствуют меланосомы не только в стромальных меланоцитах, но и в клетках пигментного эпителия. Радужка при этом розовая. У некоторых людей имеются различия в степени пигментации правого и левого глаза. Это состояние называется гетерохромией. Меланоциты радужной оболочки являются источником развития доброкачественных и злокачественных меланом.
Перед тем как остановиться на микроскопическом строении радужной оболочки, необходимо описать макроскопическое строение ее передней и задней поверхностей.
На передней поверхности различают уже упомянутый «воротничок», крипты Фукса, брыжи зрачкового края, а также сфинктер зрачка (рис. 3.8.6—3.8.8).
«Воротничок» складывается из соединительнотканных трабекул, распространяющихся от корня радужки и прерывающихся в виде гребня приблизительно в 1,6 мм от края зрачка. Он делит переднюю поверхность на две зоны: внешнюю ресничную и внутреннюю зрачковую зоны. Эти две зоны часто отличаются по цвету. Именно в этой области радужка утолщена в связи с расположением в ней малого круга кровообращения радужки.
В эмбриональном периоде сосуды малого круга кровообращения были связаны с сосудистой сумкой хрусталика. В последующем эта связь исчезает, но сохраняется «зрачковая мембрана», которая может быть обнаружена и после рождения, особенно у недоношенных детей.
Крипты Фукса располагаются на передней поверхности радужки и имеют вид ямкоподобных углублений довольно большого размера.
Эпителий радужки, покрывающий ее заднюю поверхность, иногда простирается на зрачковый край в виде брыжей. Более выражены брыжи при миозе. Именно брыжи определяют границу переднего края глазного бокала.
В голубых глазах или при атрофии стромы радужки иногда виден сфинктер зрачка в виде ленты, циркулярно расположенной вблизи зрачкового края.
На задней поверхности радужки видны складки, возникающие при сокращении дилятатора зрачка (складки Швальбе, борозды Швальбе, циркулярные борозды) (рис. 3.8.9).
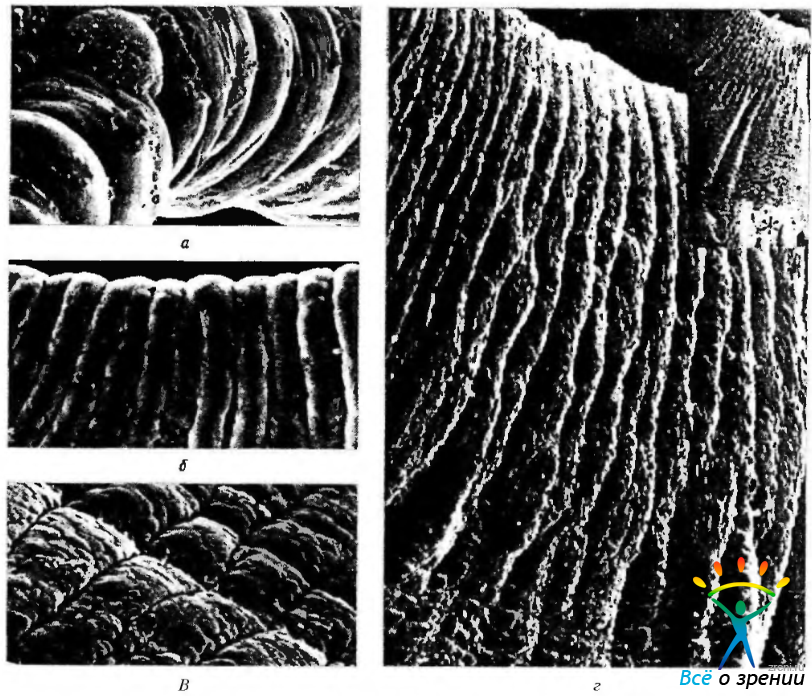
Рис. 3.8.9. Сканирующая электронная микроскопия задней поверхности радужной оболочки: а — зрачковый край (по Fredo); б—складчатость задней поверхности в области зрачкового края при миозе (по Fredo); в — борозды сокращения при мидриазе (по Fredo); г—продольные борозды на задней поверхности в области зрачкового края. Видны ресничные отростки (*) по периферии радужки (по Rodrigues et al., 1988)
Эти складки располагаются радиально. Поскольку пигментный эпителий переходит на зрачковый край, эти складки придают зрачковому краю зазубренный вид. Складки Швальбе простираются как на эпителий радужки, так и строму. Они начинаются в 1,5 мм от края зрачка, при этом образуя здесь узкую и глубокую дугу. Глубина их уменьшается по мере продвижения к периферии радужки. Постепенно они переходят на ресничное тело между ресничными отростками.
Циркулярные борозды пересекают складки Швальбе. Образуются они в результате неравномерной толщины слоя эпителиальных клеток и стромы. Наименее всего циркулярные борозды развиты в проекции расположения сфинктера. У корня радужки они более выражены.
На задней поверхности видны и так называемые «впадины». «Впадины» равномерно распределены по всей поверхности пигментного эпителия и соответствуют местам расположения десмосом между эпителиальными клетками.
При микроскопическом исследовании радужку условно подразделяют на следующие части:
- строма (мезодермальная часть),
- мышцы (производные нейроэктодермы),
- пигментный эпителий (производные нейроэктодермы).
Строма радужной оболочки. Строма радужки. как было указано выше, содержит два слоя: передний пограничный слой и строму (рис. 3.8.10).
Рис. 3.8.10. Микроскопическое строение радужной оболочки: 1 — строма; 2 - пигментный эпителий; 3— дилятатор; 4 — сфинктер; 5 - передний пограничный слой; 6 - скопление глыбистых клеток; 7 — капсула хрусталика; 8 — эпителий капсулы хрусталика; 9—хрусталиковые волокна
Передний пограничный слой стромы радужной оболочки. Длительное время считалось, что передняя поверхность радужки покрыта слоем эндотелиальных клеток. В 50-е годы XX столетия было показано, что эндотелиальные клетки на передней поверхности радужки человека обнаруживаются только при рождении и исчезают спустя 1—2 года. Замещаются они фибробластами и меланоцитами.
Передний пограничный слой радужки есть не что иное, как видоизмененный слой стромы. Отличия сводятся лишь к более плотному расположению клеток, волокон и сосудов. Плотность этого слоя существенно варьирует у разных индивидуумов.
В переднем пограничном слое можно выявить и невусоподобные структуры, источником которых являются шванновские клетки нервных стволов. При врожденной гетерохромии (повышенная пигментация радужной оболочки) количество меланоцитов в переднем пограничном слое значительно больше, чем в норме. При этом они образуют клеточные скопления, напоминающие невус. Нередко по периферии встречаются белые и желтоватые пятна (пятна Вольфа), наиболее часто обнаруживаемые в серой радужке.
Передний пограничный слой насыщен капиллярными сосудами, число которых увеличивается при воспалении радужки и при диабете. Избыточное развитие сети сосудов приводит к состоянию, известному у клиницистов как неоваскуляризация радужки, или рубеоз. Необходимо подчеркнуть, что в одной и той же радужке плотность расположения клеток меняется в различных участках.
Передний пограничный слой радужки беден соединительной тканью. На периферии он внезапно оканчивается у корня радужки. В 57% случаев выявляются соединительнотканные тяжи, распространяющиеся от поверхности раной шпоре. Иногда их путают с периферическими передними синехиями.
В переднем пограничном слое различают следующие типы клеток:
- фибробласты,
- меланоциты,
- тучные клетки,
- лимфоциты «глыбистые клетки»
- и моноциты (рис. 3.8.11, 3.8.12).
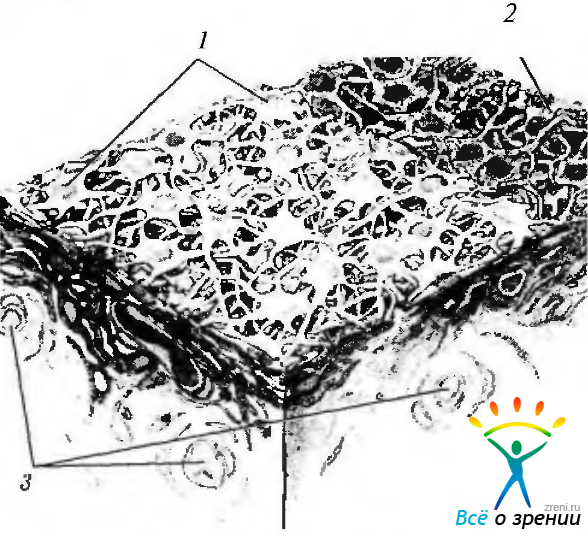
Рис. 3.8.11. Схематическое изображение взаимоотношения клеточных элементов переднего пограничного слоя радужной оболочки (по Hogan et al., 1971): 1 — фибробласты; 2 — стромальные меланоциты; 3 — кровеносные сосуды

Рис. 3.8.12. Электроннограмма (малое увеличение) переднего пограничного- слоя и стромы радужной оболочки: 1— фибробласты; 2 — меланоциты
Реже обнаруживаются макрофаги и дендритические клетки.
Фибробласты на поверхности радужки образуют скопление в виде широкой полосы, идущей от корня до зрачкового края. От тела фибробластов отходят многочисленные отростки, расходящиеся во всех направлениях, образуя при этом по поверхности радужки густую сеть. Некоторые фибробласты обладают микроворсинками и ресничками, обращенными в переднюю камеру глаза. Фибробласты лежат в сети рыхло расположенных коллагеновых волокон, пропитанных гликозаминогликанами и межтканевой жидкостью.
На периферии радужки фибробласты постепенно переходят в строму ресничного тела, а в области зрачкового края они контактируют с клетками пигментного эпителия.
В наиболее толстых участках переднего пограничного слоя радужки преобладают увеальные меланоциты. Лежат они под фибробластами, а их отростки ориентированы параллельно поверхности радужки. Подобно фибробластам, стромальные меланоциты контактируют как между собой, так и с фибробластами. В местах контакта выявляются щелевые контакты. Передний пограничный слой отсутствует в криптах и истончается в «бороздах сокращения». Он наиболее толстый в области зрачка и вблизи ресничной зоны. Коллагеновые фибриллы складываются в маленькие и большие пучки, пересекающиеся под тупым углом с образованием пространств различного размера (рис. 3.8.13, 3.8.14).

Рис. 3.8.13. Фазово-контрастная микроскопия среза стромы радужной оболочки (по Fine, Yanoff, 1972): видны пучки коллагеновых волокон (в верхнем правом углу коллагеновое волокно при большом увеличении), концентрирующиеся вокруг кровеносных сосудов и нервов

Рис. 3.8.14. Электроннограмма стромы радужной оболочки. Видны пучки коллагеновых волокон, концентрирующиеся вокруг кровеносных сосудов
Коллагеновая ткань обильна вокруг сосудов и нервов, а также в трабекулах и между мышечными пучками сфинктера. Вокруг зрачкового края пучки коллагеновых волокон ориентируются циркулярно, в то время как в области сфинктера — меридианально.
В коллагеновую сеть погружены сфинктер, кровеносные сосуды и нервы. Коллагеновый остов стромы радужки прикрепляется к переднему пограничному слою, к сфинктеру и дилятатору и продолжается на ресничное тело в виде стромы.
Выявлена четкая закономерность в трехмерной организации пучков коллагеновых волокон у многих млекопитающих. Пучки фибрилл ориентируются полуциркулярно и формируют широкие арки, идущие от зрачкового края к ресничному телу. Часть этих арок направлена по часовой стрелке, а часть — против часовой стрелки. При этом образуется система дугообразных арок, покрывающих друг друга. Схожее расположение волокон выявляется и у человека. По этой причине радужка человека содержит многочисленные межтканевые пространства, часть которых расположена радиально, а часть — меридианально (рис. 3.8.15, 3.8.16).
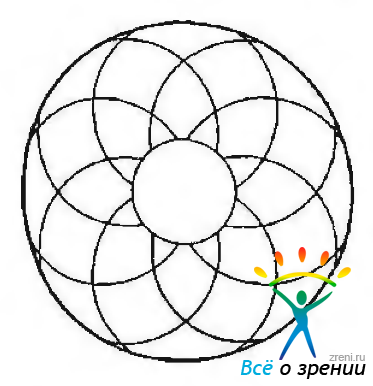
Рис. 3.8.15. Аркоподобное распределение пучков коллагеновых волокон в строме радужной оболочки
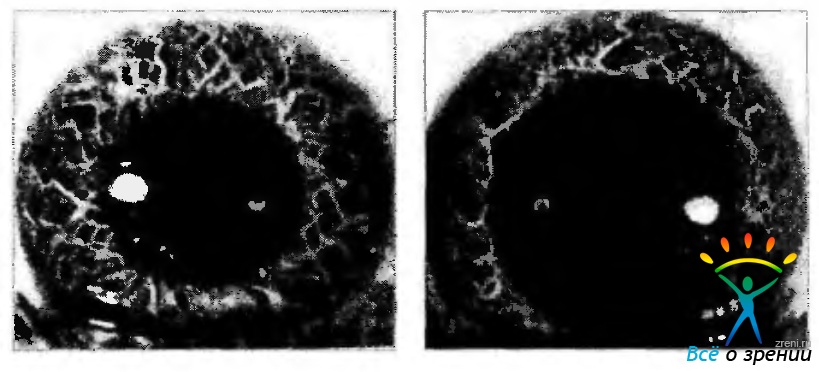
Рис. 3.8.16. Изменение структуры передней поверхности радужной оболочки при расширении зрачка. Трабекулы определяют особенности пространственной организации пучков коллагеновых волокон
Если в переднюю камеру глаза инъецировать индийскую тушь, убитые микроорганизмы (кокки) или вещества различного молекулярного веса (торотраст, декстран), то они проникают в строму радужки довольно глубоко через ущелья и крипты Фукса. При этом они концентрируются вокруг эндотелия сосудов и дилятатора. Радужка проницаема для частиц размерами не более 50—200 мкм.
Клетки стромы радужной оболочки. Как было указано выше, в строме радужки, помимо описанных выше фибробластов и меланоцитов, можно обнаружить макрофаги, моноциты, дендритические клетки и лимфоциты. Встречаются также тучные клетки двух типов. Первый тип схож с тучной клеткой конъюнктивы. Цитоплазма подобной клетки выполнена палочковидным гранулярным содержимым, имеющим на поперечных срезах вид завитка. Второй тип тучных клеток содержит большее количество гранул, выполненных аморфным электронноплотным материалом. Возможно, что эти два типа тучных клеток отражают различные стадии развития гранул и активности клеток. Описание глыбистых клеток будет приведено несколько ниже.
Большое функциональное значение имеют в радужке моноциты, макрофаги и дендритические клетки. Указанные последними клетки практически неотличимы от клеток Ларгенганса, обнаруживаемых в роговой оболочке. Дендритические клетки обнаружены также в строме ресничного тела и сосудистой оболочки. Все указанные типы клеток определяют местный иммунитет радужной оболочки, обладают рецепторами медиаторов воспаления. При воспалительной патологии глаза их количество увеличивается, и они проходят все стадии дифференциации, выполняя при этом функцию распознания чужеродного антигена и передачи полученной информации иммунным органом более высокого уровня.
Межклеточное вещество. Строма вокруг сфинктера зрачка, внутренней стенки кровеносных сосудов радужки, в эндоневрии нервов, а также вокруг мышечных волокон дилятатора состоит из коллагена VI типа. Часть волокон содержит коллаген IV типа, который также выявляется в базальных мембранах сосудов и в эндоневрии. В межклеточном веществе содержатся ламинин и фибронектин. Наибольшая их концентрация обнаружена вокруш мышц радужки. Обнаруживается и микрофибриллярный белок фибриллин.
Кровеносные сосуды радужной оболочки. Как указано ранее, большой круг кровообращения радужки сформирован в основном задними длинными ресничными артериями, в то время как внутримышечный крут кровообращения — ветвями передних ресничных артерий.
Большой круг кровообращения (рис. 3.8.17). Большой круг кровообращения радужки располагается в ресничном теле, впереди циркулярной части ресничной мышцы и впереди мышечного крута кровообращения.
Артерии радужки. Артерии радужки берут свое начало из большого круга кровообращения. Радужка также кровоснабжается перфорирующими ветвями передней ресничной артерии. Анастомозируя, они радиально сходятся у зрачкового края (рис. 3.8.18).
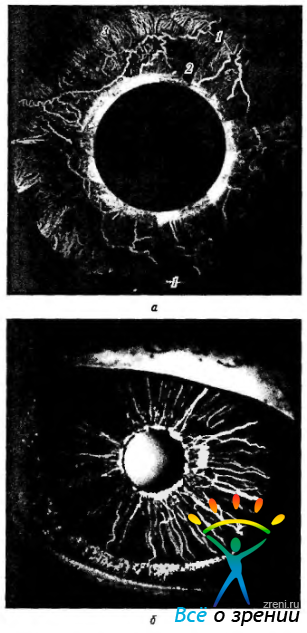
Рис. 3.8.18. Распределение артериальных сосудов переднего отдела глаза: а — архитектоника сосудов переднего отдела глаза (наполнение сосудов метилметакрилатом; монтаж по Вrоп et at., 1997) (1 — передние ресничные артерии; 2 — ресничная мышца; 3 — ресничные вены); б—флюоресцентная ангиограмма сосудов радужки
При миозе артерии прямые, но их путь становится извилистым при расширении зрачка. Подобно венам, стенка артерии толстая.
Сосуды хорошо видны при офтальмоскопии, особенно в голубых радужках, и более четко — в ресничной части. В интенсивно пигментированных радужках сосуды определяются хуже или вообще не видны. В области «воротничка» анастомозов между отдельными сосудистыми стволами наибольшее количество. Именно в этом месте образуется малый артериальный круг радужки. У зрачкового края радужки артериолы переходят в капилляры, а затем возвращаются к корню радужки в виде вен.
Капиллярное сплетение. Вокруг сфинктера и дилятатора располагается довольно густое сплетение капиллярных сосудов. В ресничной области радужки капиллярное сплетение становится менее плотным. Выражено оно слабо или полностью отсутствует в переднем пограничном слое радужки.
Строение сосудов радужки. Сосуды радужки обладают исключительно толстой адвентицией (рис. 3.8.19, 3.8.20).
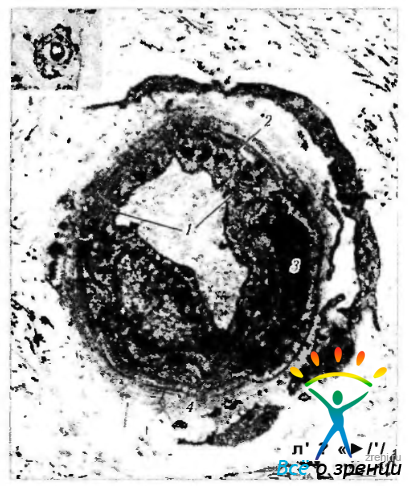
Рис. 3.8.19. Ультраструктурные особенности кровеносного сосуда стромы радужной оболочки: 1 - межклеточные контакты на апикальной поверхности эндотелиальных клеток; 2 — базальная мембрана; 3 — перицит; 4 — адвентиция
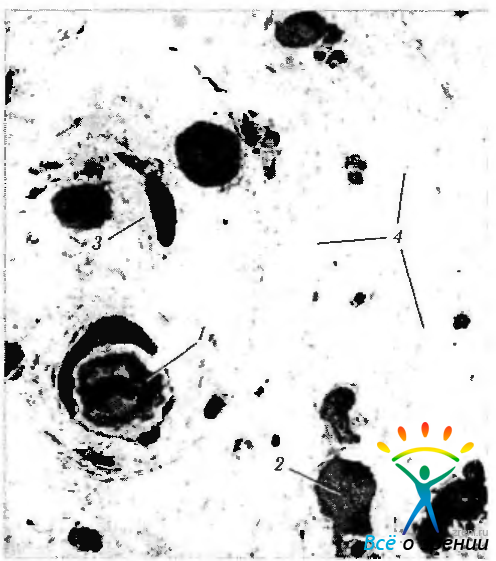
Рис. 3.8.20. Строма радужной оболочки: 1 — кровеносный сосуд; 2 — фибробласт; 3 — стромальный меланоцит; 4 — диффузно-распределенные поперечно срезанные коллагеновые волокна
При исследовании стенки сосудов после окрашивания по Маллори выявляется довольно сложное ее строение. Стенка сосуда как будто бы состоит из двух вложенных друг в друга цилиндров. Наружный слой, окрашивающийся в интенсивно синий цвет, складывается из нежных коллагеновых волокон, в то время как внутренний изнутри покрыт слоем эндотелиальных клеток и содержит мышечные и эластические волокна. Между ними определяются довольно широкая светлая зона, состоящая из коллагеновых волокон — tunica media, являющаяся специфическим признаком сосудов радужки. Эта зона предотвращает спадение стенок сосудов при деформации радужки во время сужения и расширения зрачка. Благодаря этому внутриартериальное давление сохраняется постоянным.
Необходимо указать, что классифицировать тип сосудов радужной оболочки довольно сложно. Только капилляры с калибром просвета, равным 10—15 мкм, можно легко отнести к капиллярам. Сосуды другого калибра чаще относятся к артериолам и посткапиллярным венулам.
В артериальной стенке выявляется четыре слоя:
- Слой эндотелиальных клеток.
- Слой мышечных клеток.
- Средняя оболочка, содержащая фиброциты и коллагеновые волокна.
- Адвентиция. Внутренняя зона адвентиции состоит из нежных коллагеновых фибрилл VI типа (30—60 нм), соединяющих базальную мембрану с наружной зоной. Наружная зона структурно отличается от внутренней зоны. В ней коллагеновые волокна имеют различный диаметр (30—125 нм). Артериолы обладают более толстой адвентицией, чем венулы.
Эндотелий сосудов. Эндотелий сосудов радужки человека не имеет «фенестр». Аналогичное строение сосудов и у обезьян, кроликов, свиньи, морской свинки и кошек.
Поры обнаруживаются только в эндотелии котят. У крыс обнаружен как фенестрированный, так и нефенестрированный эндотелий. Эндотелиальная выстилка сосудов обезьян непрерывна и клетки плотно скреплены между собой при помощи запирательных пластин.
Эндотелиальные клетки сосудов радужки соединены между собой еще двумя типами межклеточных контактов. Это «замыкающая пластинка» и «щелевые контакты». По этой причине через межклеточные пространства не проникают высокомолекулярные вещества (например, пероксидаза хрена).
Цитоплазма эндотелиальных клеток при светооптической микроскопии светлая и бесструктурная. Характерно для нее наличие телец Вейбель Паладе (Weibel Palade) (палочковидные включения) и кристаллоидных включений. Хорошо развита шероховатая эндоплазматическая сеть. Количество приведенных включений существенно увеличивается при хронической простой глаукоме.
Перициты (рис. 3.8.19). Перициты сосудов радужки имеют обычное для других тканей строение, включая сосуды сетчатки. Капилляры большого калибра окутаны непрерывным слоем перицитов и толстой базальной пластинкой. С уменьшением калибра сосудов уменьшается и плотность перицитов.
Сфинктер зрачка (т. sphincter pupillae). В зрачковой зоне стромы радужки концентрически распространяется гладкая мышца шириной 0,75—0,8 мм и толщиной 0,1—0,17 мм (рис. 3.8.21, 3.8.22).
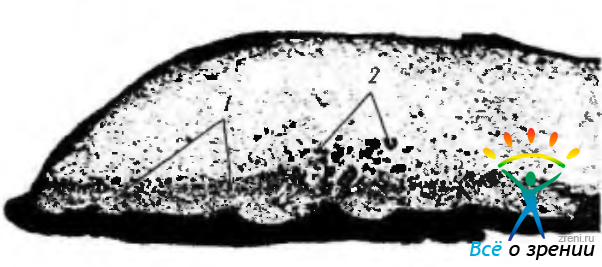
Рис. 3.8.21. Зрачковый край радужной оболочки: вокруг сфинктера зрачка (1 ) определяется скопление интенсивно пигментированных «глыбистых» клеток (2)
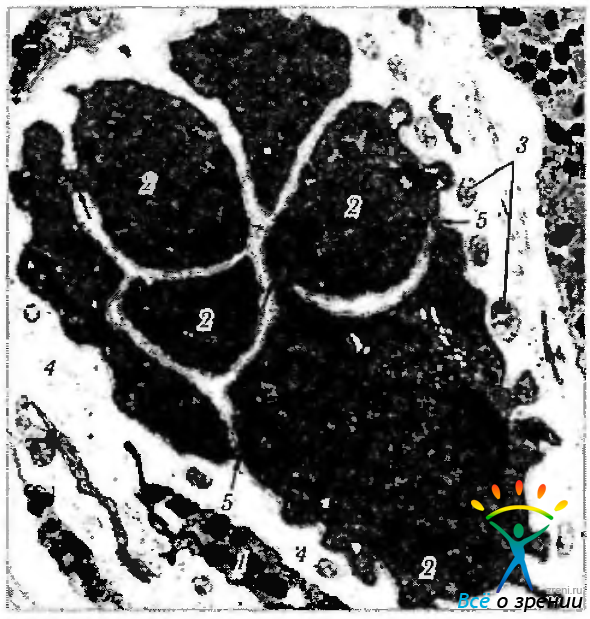
Рис. 3.8.22. Электроннограмма меридианального среза сфинктера радужной оболочки (по Hogan et al., 1971): 1 — стромальный меланоцит; 2 — мышечные волокна; 3 — нервы, окружающие мышечные волокна; 4 — базальная мембрана, окружающая мышечные волокна; 5 — электронноплотные уплотнения, расположенные на мембране мышечных волокон
При ее сокращении зрачок суживается.
Когда зрачок уменьшает свой диаметр от 8,0 до 1,5 мм, сфинктер укорачивается на 87% своей длины. При этом толщина мышцы существенно не увеличивается. По этой причине сфинктер является уникальной мышцей, поскольку обладает так называемым телескопическим типом сокращения. Подобный тип сокращения характерен только для поперечнополосатой мышцы. Сзади мышца отделена от пигментного эпителия радужки слоем соединительной ткани.
Мышечные клетки соответствуют всем критериям гладких мышц. Они веретеновидной формы и ориентированы параллельно зрачковому краю. Пучки мышечных клеток плотно упакованы и отделены тонкими прослойками соединительной ткани. Среди пучков коллагеновых волокон распределяются артериолы, капилляры, чувствительные и двигательные нервы.
Электронномикроскопически выявлено, что «мышечный пучок» сфинктера образован 5—8 мышечными клетками, плотно соединенными между собой при помощи специализированных органоидов.
Как и в других мышечных тканях, нервы не проникают в глубь группы мышечных клеток, а прилежат к их поверхности. В связи с указанным взаимоотношением нервов и мышечных клеток многие исследователи предполагают, что группы мышечных клеток образуют «функциональные единицы». По-видимому, только одна клетка «функциональной единицы» иннервирована, а плотные межклеточные контакты позволяют распространяться деполяризации и на другие клетки.
Ядра гладкомышечных клеток лежат в центре цитоплазмы. Видны многочисленные внутри-цитоплазматические органоиды. Это аппарат Гольджи, шероховатый эндоплазматический ретикулум. множество полирибосом и миофиламентов. Вдоль внутренней поверхности клеточной мембраны располагаются пиноцитозные пузырьки.
Базальная мембрана сфинктера радужки не отличается от базальной мембраны других гладкомышечных клеток. Эта мембрана входит в контакт с коллагеновыми фибриллами, отделяющими мышечные группы, между которыми лежат нервные волокна. На отдельных группах мышечных клеток нервы формируют пучки. Обычно пучок состоит из 2—4 нервных аксонов, окруженных шванновскими клетками. Аксоны без шванновской оболочки оканчиваются непосредственно на мышечной клетке.
Несмотря на очень схожее строение сфинктера с другими гладкими мышцами, сфинктер является необычной мышцей, поскольку целиком образуется из нейроэктодермальных клеток, которые мигрируют в строму из нейроэпителия в эмбриональном периоде. Подтверждением тому может служить присутствие в этих мышечных клетках меланиновых гранул, которые по форме и размерам не отличаются от меланосом нейроэпителия.
Мышца иннервируется парасимпатическими нервными волокнами, исходящими из ядра Якубовича—Эдингера—Вестфаля.
Глыбистые клетки. Глыбистые клетки (рис. 3.8.21) (клетки Коганей (Koganei)) представляют собой интенсивно пигментированные клетки округлой формы, которые обычно обнаруживаются впереди сфинктера. Эти клетки достигают размера 100 мкм и на их поверхности видны ворсинчатые отростки длиной 1 — 2 мкм и шириной 0,1 мкм. Внутрицитоплазматические пигментные гранулы имеют вид пузырьков, размер которых достигает 10 мкм. Пузырьки наполнены зернами меланина и представляют собой резидуальные тельца или вторичные лизосомы. В пузырьках выявляются также липиды и гранулярный матрикс.
Морфологические особенности зерен меланина, содержащихся в резидуальных тельцах, варьируют в зависимости от локализации глыбистых клеток. Так, в глыбистых клетках задних слоев стромы радужки гранулы идентичны по форме и размерам меланиновым гранулам пигментного эпителия радужки. В глыбистых клетках передних участков стромы преобладают мелкие меланиновые гранулы, схожие с гранулами стромальных меланоцитов.
Ряд авторов приводят морфологические, эмбриологические и экспериментальные данные, позволяющие предположить, что существует два типа глыбистых клеток. Большинство глыбистых клеток первого типа относят к макрофагам. Клетки первого типа обладают тонкими микроворсинками, а цитоплазма выполнена зернами пигмента различного размера. Ядро лежит эксцентрично. В цитоплазме определяются капельки липидов и зерна липофусцина. Эти клетки не обладают базальной мембраной. Подобные клетки редки у молодых людей и легко обнаруживаются у пожилых.
Существует также второй тип глыбистых клеток. Этот тип клеток формирует маленькие группы, окруженные базальной мембраной. Меланиновые гранулы в их цитоплазме имеют нейроэпителиальное происхождение. Группы клеток второго типа окружены базальной мембраной. Соединены между собой они при помощи десмосом, а в цитоплазме определяются микрофиламенты и микропиноцитозные пузырьки.
Предполагается, что глыбистые клетки второго типа имеют нейроэктодермальное происхождение. Эти клетки мигрируют в строму радужки в эмбриональном периоде по направлению будущего сфинктера, но, не достигая места конечной локализации, остаются в строме. Оставшись в строме, они не подвергаются окончательной дифференциации (мышечные клетки).
Пигментный эпителий радужной оболочки (радужковая часть сетчатки; pars iridica retinae) (рис. 3.8.23—3.8.26).

Рис. 3.8.23. Сканирующая электронная микроскопия задней поверхности радужной оболочки: видны складки пигментного эпителия радужки. Стрелками указаны эритроциты
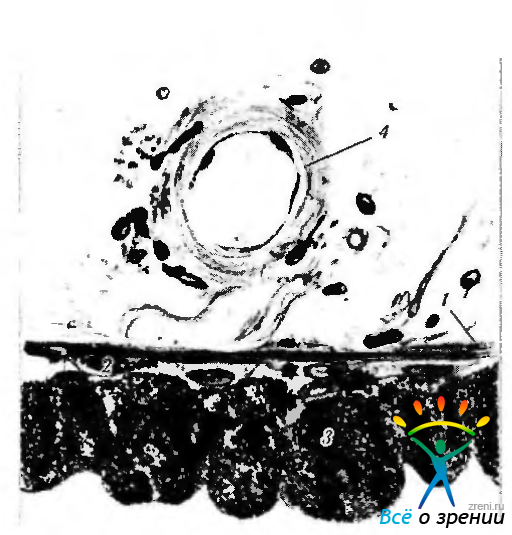
Рис. 3.8.24. Задняя поверхность радужной оболочки (прикорневая зона): 1— дилятатор зрачка; 2 — передний слой пигментного эпителия: 3 — задний слой пигментного эпителия; 4 — сосуд стромы
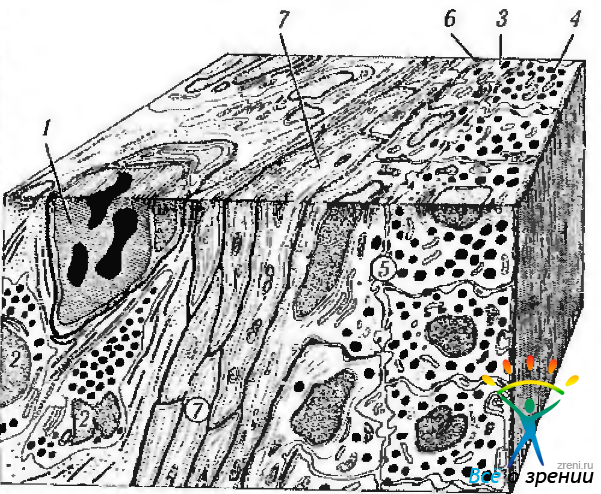
Рис. 3.8.25. Схематическое изображение задней поверхности радужной оболочки: 1 — капиллярный сосуд; 2 — стромальный меланоцит; 3— передний слой пигментного эпителия; 4 — задний слой пигментного эпителия; 5 — пространство между передним и задним слоями пигментного эпителия; 6 — базальная мембрана; 7 — мышечные волокна дилятатора
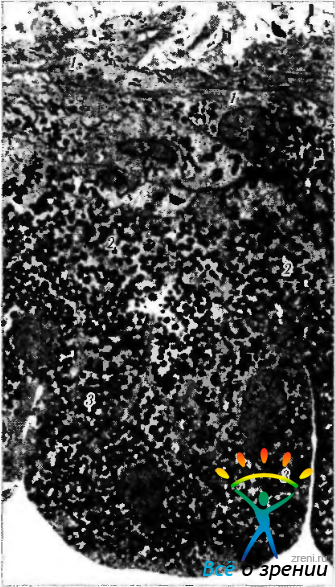
Рис. 3.8.26. Электроннограмма (малое увеличение) двухслойного пигментного эпителия радужной оболочки: 1 — дилятатор зрачка; 2 — передний слой пигментного эпителия; 3 — задний слой пигментного эпителия
Заднюю поверхность радужки покрывает двойной слой пигментного эпителия, имеющий толщину 12 мкм. Распространяясь на переднюю поверхность радужки, пигментный эпителий нередко вдоль зрачкового края образует интенсивно пигментированную полоску.
По мере продолжения кзади пигментный эпителий трансформируется в пигментный эпителий ресничного тела, а далее — в нейрональную сетчатку. Эпителиальные клетки пигментного эпителия на задней поверхности радужки образуют гроздья и борозды. Выраженное проявление борозд в области основания радужки связано чаще всего с пролиферацией эпителиоцитов. При этом формируются складки, участвующие в образовании кист или пигментированных псевдоаденом. В области сфинктера многие радиальные или продольные борозды замещены циркулярными бороздами. Несколько кнаружи сфинктера проходит нежная зрачковая радиальная борозда. Передний слой клеток пигментного эпителия отделен от стромы базальной мембраной.
Апикальная часть клеток переднего слоя пигментного эпителия обращена к апикальной поверхности клеток заднего эпителия. Между ними определяется пространство (20 нм), выполненное микроворсинами. Множество десмосом и щелевых контактов скрепляют апикальные поверхности переднего и заднего слоев пигментного эпителия.
Эпителиоцитам характерно наличие множества разнообразных внутрицитоплазматическпх органоидов. К ним относятся митохондрии, шероховатый и гладкий эндоплазматический ретикулум. Определяется большое количество рибосом, а также аппарат Гольджи (рис. 3.8.27).
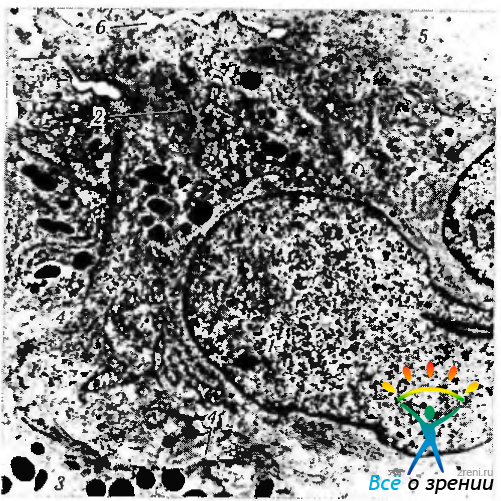
Рис. 3.8.27. Особенности ультраструктурной организации клеток переднего слоя пигментного эпителия радужной оболочки (по Hogan et al., 1971): 1 — ядра клеток; 2 — миофиламенты; 3 — клетка заднего слоя пигментного эпителия; 4 — межклеточные контакты между клетками переднего и заднего слоя эпителия: 5—питоплазматические отростки клеток, распространяющиеся в строму радужки и содержащие миофиламенты: 6 — базальная мембрана, окружающая мышечную часть клеток
Цитоплазма эпителиоцитов выполнена большими меланиновыми гранулами (сферичные имеют диаметр 0,8 мкм, а овоидные — 0,5— 1,3 мкм). Эти гранулы существенно отличаются от более мелких неправильно овальной формы меланиновых гранул стромальных меланоцитов радужки.
Клетки заднего слоя пигментного эпителия радужки сохраняют свою кубовидную форму практически на всем протяжении, за исключением прикорневой области, т. е. в области формирования дилятатора — второй гладкой мышцы радужки. Пирамидальную форму эпителиальные клетки приобретают лишь в ямках и складках задней поверхности радужки. При этом высота их колеблется от 36 до 55 мкм, а ширина — от 16 до 25 мкм.
По периферии радужки клетки постепенно депигментируются по мере перехода в эпителий ресничного тела.
При использовании электронной микроскопии базальная мембрана в основании клеток пигментного эпителия имеет обычное строение. Боковые поверхности клеток отдают интердигитации, погружающиеся в соседнюю клетку. Несмотря на наличие между соседними клетками довольно большого количества десмосом, между клетками сохраняется межклеточное пространство шириной 200 мкм.
Хорошо известно, что задний и передний слои пигментного эпителия радужки легко отделяются друг от друга При этом образуются псевдокисты.
Пигментные клетки эпителия радужки обладают многими функциями, некоторые из которых хорошо изучены, но многие неизвестны. Недавно обнаружена способность эпителиоцитов радужки, как и пигментного эпителия ресничного тела, ингибировать активность Т-лимфоцитов. что. возможно, предотвращает развитие апоптоза в условиях различных патологических состояний.
Продолжение в следующей статье: Сосуды и сосудистая оболочка глазного яблока ? Часть 2
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0