Сосуды и сосудистая оболочка глазного яблока │ Часть 2


Описание
Дилятатор зрачка (dilatator pupillae) (рис. 3.8.28).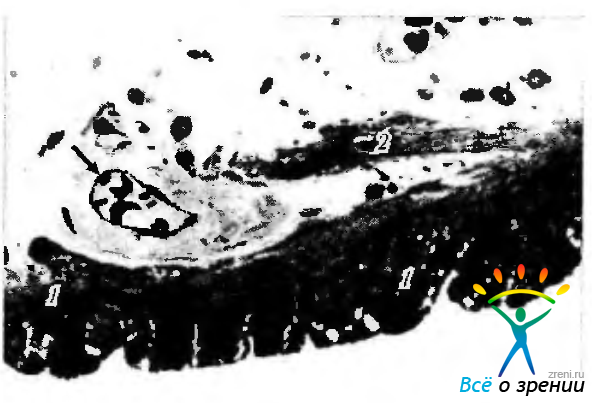
Рис. 3.8.28. Прикорневая область радужной оболочки: хорошо видны двухслойный пигментный эпителий ( 1 ) и дилятатор зрачка (2). Стрелкой указан кровеносный сосуд радужки, отличающийся резко утолщенной стенкой
Дилятатор, в отличие от сфинктера, представляет собой не что иное, как слой клеток пигментного эпителия. Эти клетки обладают способностью формировать в цитоплазме миофибриллы. То есть одна часть клетки практически не отличается от клеток пигментного эпителия, а другая — от гладкой мышцы.
«Мышечная» часть клеток переднего эпителия имеет толщину 4 мкм. Отростки клеток пигментного эпителия, содержащие миофибриллы, веретеновидной формы, шириной 7 мкм. Длина их равна 60 мкм. Диаметр миофиламентов равняется 3 нм. Эти отростки лежат в плоскости радужки и формируют 3—5 слоев мышечных клеток. Ядра мышечных клеток, размером 4—6x14 мкм, лежат в «эпителиальной» части клетки. Там же локализуются органоиды и зерна нейромеланина.
Как и в гладких мышцах другой локализации, миоциты обладают базальной мембраной. Вдоль клеточной мембраны располагаются полудесмосомы, а также «плотные соединения», напоминающие обнаруживаемые в сфинктере.
Вблизи «мышечных» отростков распределяются немиелинизированные нервные волокна, аксоны которых входят в плотный контакт с мышечными волокнами, будучи отделенными пространством в 200 мкм.
Нервы радужной оболочки. Нервы радужка получает от длинных и коротких ресничных нервов, которые сопровождают соответствующие артерии. Нервные стволы прободают склеру вокруг зрительного нерва и направляются между сосудистой оболочкой и склерой к ресничному телу, образуя сплетение. В области сплетения отделяются многочисленные веточки, значительная часть которых немиелинизирована. Эти волокна, в свою очередь, образуют еще три сплетения. Первое из них лежит в переднем пограничном слое. Предполагают, что оно преимущественно сенсорное. Второе сплетение образуется вокруг крупных кровеносных сосудов радужки. Третье сплетение прилегает к передней поверхности дилятатора. Указанные сплетения обеспечивают иннервацию всех слоев радужки, кроме заднего слоя пигментного эпителия.
От сплетения, расположенного вблизи дилятатора, отходят немиелинизированные волокна, проникающие к мышечным клеткам. Дилятатор получает симпатическую, сфинктер парасимпатическую иннервацию. При этом в обеих мышцах обнаруживается как адренэргическая, так и холинэргическая иннервация. От 60 до 75% рецепторов относятся к мускариновым третьего типа. Подробно изучена физиологическая роль мускариновых рецепторов при сокращении и расслаблении мышц радужки, влияние на этот процесс различных агонистов и антагонистов.
Распространение жидкости и веществ в радужной оболочке. Передняя поверхность радужки и ее строма свободно пропускают жидкость камерной влаги и растворенные в ней вещества. В этом отношении передняя и задняя камеры отличаются в отношении проходимости жидкости в строму радужки.
Передняя камера не герметична. Жидкость оттекает через эндотелий роговой оболочки, строму радужки или строму ресничного тела. Кроме того, существуют и специальные системы — дренажная и увеосклеральная. Задняя камера полностью изолирована эпителием радужки и ресничного тела.
Особенности строения капилляров радужки и ресничного тела различны. Капилляры ресничного тела проницаемы для воды, а капилляры радужки нет. Эндотелий сосудов радужки не обладает способностью активно транспортировать вещества через цитоплазму.
Таким образом, в нормальных условиях лишь незначительное количество белков плазмы достигает передней камеры посредством сосудов радужки. Белок может «просачиваться» в строму ресничного тела, а затем достигать передней камеры в области корня радужки.
Непрерывный слой нефенестрированных эндотелиальных клеток радужки предотвращает выход белков и молекул радиоактивных индикаторов из просвета сосуда в строму радужки. При воспалении радужки (ирит) этот барьер нарушается, приводя к отеку стромы. Freddo, Sacks-Wilner наблюдал упрощение и разрушение межклеточных контактов при эндотоксин-индуцированном увеите у кроликов. При этом радиоактивный трейсер свободно проходил между эндотелиальными клетками. Хотя сосуды сетчатки подобны по проходимости сосудам радужки, они реагируют на медиаторы воспаления по-другому. Сосуд радужки теряет «герметичность» в отношении частиц углерода или торотраста после введения гистамина. Проницаемость сосудов сетчатки не увеличивается в подобных условиях.
Существуют и видовые различия в реакции сосудов радужки. Так, при проведении парацентеза проницаемость сосудов радужки повышается у кошек, кроликов и крысы, но не повышается у обезьяны.
Ресничное тело
Ресничное тело (corpus ciliare) выполняет многообразные функции. Основными из них являются синтез камерной влаги, синтез химических компонентов стекловидного тела, а также участие в процессах аккомодации.
При макроскопическом исследовании ресничное тело имеет треугольную форму. Основание этого треугольника лежит у корня радужки, а вершина — у зубчатой линии (рис. 3.8.29).
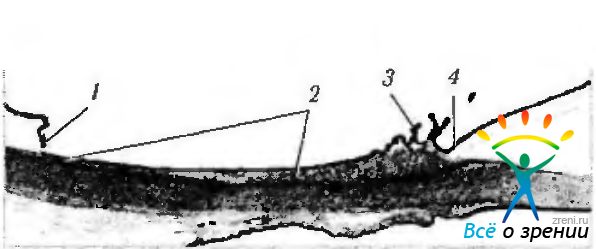
Рис. 3.8.29. Топографические особенности ресничного тела: 1 — зубчатая линия; 2 — ресничный кружок; 3— ресничный венец; 4 — корень радужной оболочки; 5 — хрусталик
Передняя поверхность ресничного тела формирует часть угла передней камеры и продолжается кпереди в виде увеальной трабекулярной сети и корня радужки. Кзади от зубчатой линии ресничное тело постепенно переходит в сосудистую оболочку глаза.
Ширина (передне-задний размер) ресничного тела колеблется от 6,0 до 6,5 мм. Шире оно снизу и темпорально (5,6—6,3 мм), а уже — вверху и назально (4.6—5,2 мм). Необходимо помнить, что размер ресничного тела прямо коррелирует с передне-задним размером глаза. Так, при буфтальме оно достигает 10 и более миллиметров.
Граница между ресничным телом и сетчаткой при транссклеральной диафаноскопии выглядит темной полосой, поскольку с обеих сторон зубчатой линии пигментный эпителий интенсивно окрашен. Ширина этой полосы — 1,5—2,0 мм. Передняя часть полосы шириной 1,0—1,5 мм принадлежит ресничному телу, что необходимо учитывать при хирургическом проникновении в глазное яблоко через ресничное тело.
Ресничное тело легко механически отделить от склеры. Для этого достаточно пересечь место его прикрепления к склеральной шпоре. Поверхность склеры, соответствующая локализации ресничного тела, гладкая и слабо пигментирована. Видны лишь каналы, через которые проникают ресничные артерии и нервы.
Ресничное тело четко разделяется на 2 части: часть, имеющую многочисленные гребешки (ресничный венец; corona ciliaris), и широкую плоскую заднюю часть (ресничный кружок; orbicuhis ciliaris; pars plana). Ширина ресничного венца равняется 2,0 мм, а плоской части — 4,0—4,5 мм.
Ресничный венец состоит приблизительно из 70—80 небольших гребешков, ориентированных радиально (рис. 3.8.30; 3.8.31, 3.8.32).
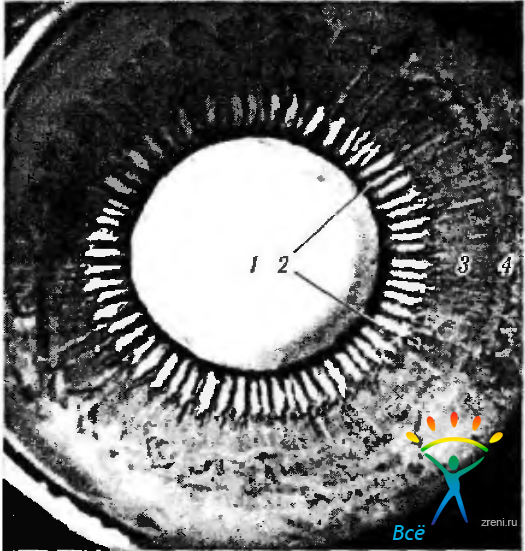
Рис. 3.8.30. Строение ресничного тела при рассмотрении его со стороны стекловидного тела: 1 — задняя поверхность хрусталика; 2— ресничный венец; 3 - ресничный кружок; 4 — зубчатая линия

Рис. 3.8.31. Строение ресничного тела: а — особенности взаимоотношения ресничного тела со структурами переднего отдела глаза (окрашивание гематоксилином и эозином) (1 — роговица; 2— угол передней камеры; 3 — радужная оболочка; 4 - ресничный поясок; 5 — ресничная мышца; 6 — хрусталик; 7—шлеммов канал); б - микроскопическое строение ресничных отростков (трехцветный метод окрашивания по Массону) (1—строма; 2—ресничная вена; 3—непигментированный эпителий; 4—пигментированный эпителий
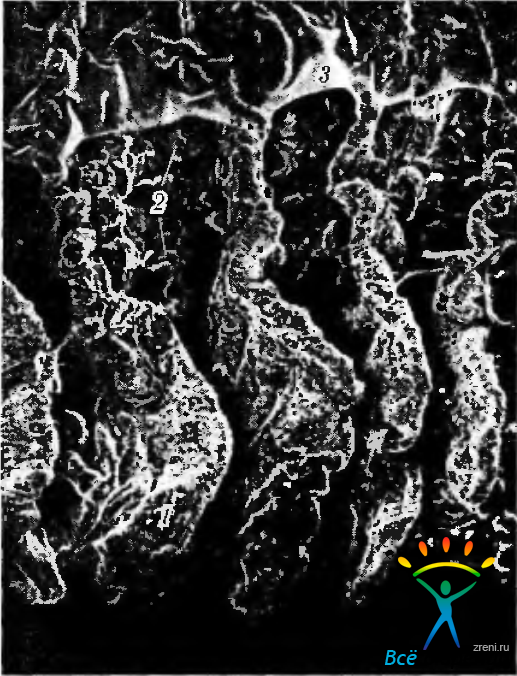
Рис. 3.8.32. Сканограмма отростков ресничного тела: 1- большие отростки; 2 - маленькие отростки; 3 - остатки стекловидного тела, прилежащие к ресничному кружку
В пространстве между гребешками лежат маленькие, неравномерно пигментированные складки (ресничные складки; plicae ciliaris).
Ресничные отростки располагаются симметрично и разнообразны в размере (длина 2,0 мм; ширина 0,5 мм). Иногда (16% случаев) видны гигантские отростки, чаще располагающиеся с назальной стороны. Гигантские отростки нередко сочетаются с аномалиями развития цинновой связки и наличием периферической дегенерации сетчатки.
Плоская часть ресничного тела простирается от заднего края ресничных гребешков до зубчатой линии (4 мм). Таким образом, отношение ширины плоской и отростчатой частей ресничного тела на меридиональных срезах равно 2:1. Плоская часть ресничного тела неравномерно пигментирована. Более пигментирована она с темпоральной стороны.
Плоская часть ресничного тела содержит относительно небольшое количество сосудов. По этой причине оперативное вмешательство в этой области не сопровождается массивным кровотечением и повреждением сетчатки. У новорожденных ресничное тело короче, а его плоская часть смещена кпереди.
Более подробно размеры ресничного тела и его структурных образований приведены в табл. 3.8.1.
Таблица 3.8.1. Размеры ресничного тела
[banner_centerrs] {banner_centerrs} [/banner_centerrs]

В соответствии с особенностями микроскопического строения ресничное тело можно разделить на 6 слоев (рис. 3.8.33, 3.8.34):
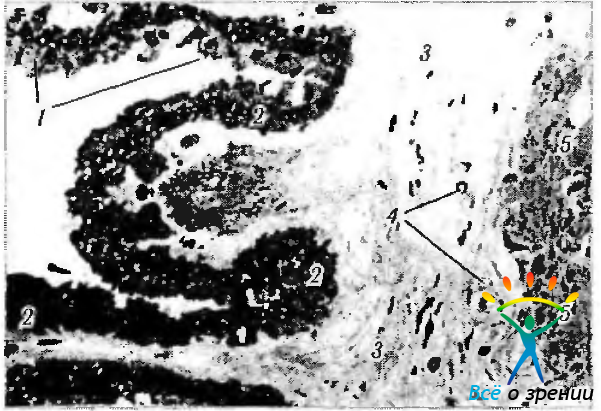
Рис. 3.8.33. Микрофотография ресничного венца ресничного тела (по Hogan et al., 1971): 1 — беспигментный эпителий (внутренний слой), содержащий незначительное количество пигментных гранул; 2 — клетки пигментированного слоя (наружного), содержащие большое количество зерен меланина в апикальной части; 3— строма ресничного тела; 4 — кровеносные сосуды стромы; 5 — участок ресничной мышцы

Рис. 3.8.34. Схематическое изображение строения пигментного эпителия ресничного тела (по Poliquen, 1969): 1 — капиллярный сосуд; 2 — фибробласт; 3 — базальная мембрана; 4 — нервный ствол; 5 — наружный слой клеток пигментного эпителия; 6— внутренний слой клеток пигментного эпителия, 7 — базальная мембрана
- супрацилиарный слой (супрахориоидальное пространство);
- ресничная мышца;
- слой сосудов;
- наружная базальная мембрана;
- эпителий;
- внутренняя базальная мембрана.
Основываясь на данных эмбриологии, т. е. учитывая особенности происхождения той или иной структуры, ресничное тело может быть разделено на две части (слоя): внутренняя часть (нейроэпителиальная) и наружная (увеальная, мезодермальная). Эту классификацию мы и будем использовать при последующем изложении материала.
Нейроэпителиальная часть (рис. 3.8.31, 3.8.33—3.8.36).
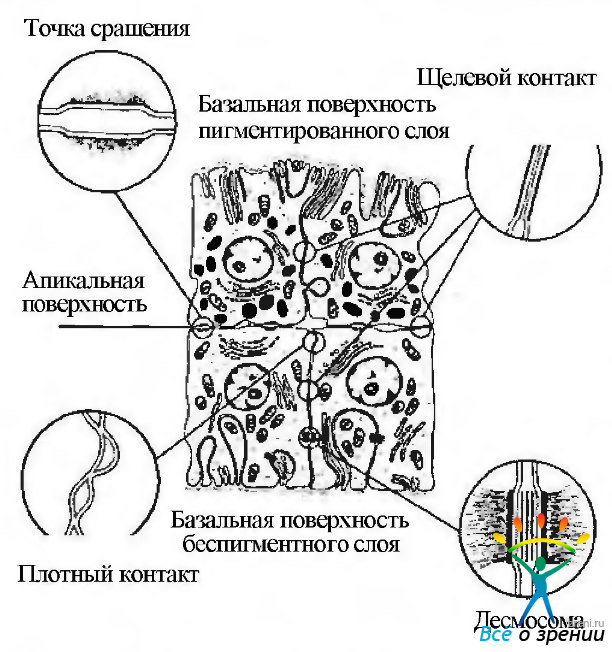
Рис. 3.8.35. Типы межклеточных контактов, обнаруживаемых между клетками пигментного эпителия ресничного тела (по Raviola et al., 1977)
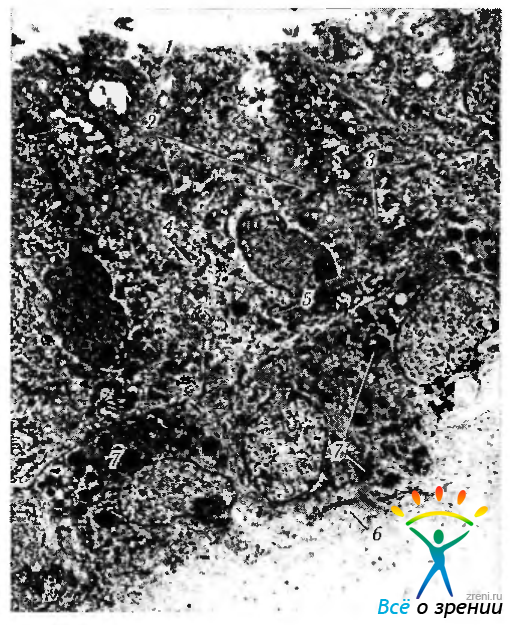
Рис. 3.8.36. Электроннограмма эпителия ресничного тела: 1—внутренняя пограничная мембрана; 2—митохондрии; 3 -гладкий эндоплазматический ретикулум; 4 — аппарат Гольджи; 5 — цитоплазматические отростки, распространяющиеся в подлежащую строму пигментных эпителиальных клеток; 6 — базальная мембрана пигментных эпителиальных клеток; 7 — меланиновые гранулы пигментированного слоя эпителия
Нейроэпителиальная часть ресничного тела формируется в эмбриональном периоде и представляет собой два слоя эпителиальных клеток зрительного бокала. При этом клетки эпителиальных слоев прилежат друг к другу своими апикальными поверхностями. Наружный (пигментированный) слой ресничного эпителия является продолжением пигментного эпителия сетчатки. При этом клетки этого слоя практически не отличаются от клеток пигментного эпителия сетчатки.
Клетки внутреннего слоя ресничного эпителия по происхождению аналогичны клеткам сенсорной части сетчатки. Сенсорная часть сетчатки внезапно прерывается у зубчатой линии, продолжаясь кпереди на ресничное тело, формируя внутренний беспигментный слой ресничного эпителия.
В области ресничного тела просвет зрительного пузырька полностью облитерируется. При этом два слоя эпителиальных клеток плотно сращены благодаря наличию многочисленных межклеточных контактов (рис. 3.8.34). Попытка их механического разделения приводит к разрушению апикальной части клеток.
Кроме указанных контактов между двумя эпителиальными клеточными слоями обнаруживается и третий компонент, участвующий в межклеточном соединении — межклеточная цементирующая субстанция. Она расположена внеклеточно между апикальными поверхностями клеточных мембран и имеет вид сплошной линии.
Беспигментный слой пигментного эпителия и внутренняя базальная мембрана. Внутренняя базальная мембрана ресничного тела располагается с базальной (витреальной) стороны беспигментных клеток эпителия. Она связана с внутренней пограничной мембраной сетчатки сзади и радужки — спереди.
У новорожденных внутренняя базальная мембрана имеет типичное строение базальной мембраны. Состоит она из зернистого слоя толщиной 30 нм и прозрачного слоя (lamina lucida), толщина которого равна 50 нм. Состоит она из коллагеновых волокон I, III и IV типов, а также ламинина.
В возрасте 3 лет базальная мембрана начинает утолщаться и становится многослойной.
По направлению базальной мембраны базальные поверхности беспигментных клеток образуют многочисленные складки и отростки, которые оплетаются волокнами мембраны и коллагеновыми волокнами основания стекловидного тела.
Беспигментные эпителиоциты имеют кубическую форму в отростчатой части ресничного тела (ширина 12—15 мкм и высота 10—15). В плоской части ресничного тела они цилиндрические (ширина 6—9 мкм и высота 30 мкм). Наиболее высокие клетки лежат вблизи зубчатой линии, возможно, в результате их тракции стекловидным телом. Многими исследователями отмечено увеличение высоты клеток с возрастом.
Между лежащими по соседству беспигментными эпителиальными клетками обнаруживаются щелевидные пространства различной ширины. Эти пространства заполнены прозрачным внеклеточным муцинозным материалом, положительно окрашивающимся при проведении гистохимических реакций, направленных на выявление кислых гликозаминогликанов.
Цитоплазма эпителиоцитов насыщена органоидами, что указывает на их высокую секреторную функцию. Особенно много в цитоплазме митохондрий. Митохондрии являются основным источником энергии для обеспечения активного транспорта электролитов и молекул небольшого размера через цитоплазму эпителиальных клеток в процессе формирования камерной влаги.
В эпителиальных беспигментных клетках также хорошо развит шероховатый и гладкий эндоплазматический ретикулум. Цитоплазма эпителиоцитов содержит большое количество промежуточных микрофиламентов и филаментов актина. Необходимо отметить, что микрофиламенты положительно окрашиваются при выявлении виментина и цитокератана.
Боковые поверхности беспигментных клеток образуют многочисленные складки цитоплазматической мембраны, погруженные в цитоплазму соседних клеток (интердигитации). Складки значительно увеличивают площадь контакта между клетками, что наиболее выражено в передней части ресничных отростков.
Апикальные поверхности беспигментных эпителиоцитов ресничных отростков при макроскопическом исследовании белые, что связано с высоким содержанием в их цитоплазме гликогена.
На переднем склоне ресничных гребешков по мере приближения к радужной оболочке в части эпителиальных клеток появляются зерна меланина. Степень пигментации постепенно нарастает, пока они не переходят в слой пигментного эпителия корня радужки.
Хотя внутренний слой ресничного эпителия и называется непигментированным, с возрастом возможно появление в их цитоплазме зерен меланина и липофусцина.
Большое значение в функционировании эпителия ресничного тела имеют межклеточные контакты, расположенные как между беспигментными клетками, так и между беспигментными и пигментированными клетками (рис. 3.8.34, 3.8.35). Наиболее важным в функциональном смысле типом контакта является так называемая лента замыкания, располагающаяся на боковых поверхностях беспигментных клеток в апикальной их части. Встречаются также контакты типа «кепки», соединяющие боковые поверхности пигментированных и реже беспигментных клеток, щелевые контакты и десмосомы. Особое внимание в настоящее время уделяется изучению щелевых контактов. Предполагают, что эти контакты обеспечивают существование так называемого «электрического» взаимодействия эпителиальных клеток между собой. Именно подобный тип взаимодействия позволяет эпителиальному пласту функционировать как синцитий и, возможно, обеспечивает координацию секреторной деятельности всего эпителиального пласта.
Наличие перечисленных контактов между клетками, особенно ленты замыкания, делает практически невозможным проникновение между клетками высокомолекулярных веществ путем диффузии.
Как неоднократно указывалось выше, беспигментный слой эпителиальных клеток ресничного тела является основным структурным элементом, обеспечивающим секрецию камерной влаги. Вышеприведенные светооптические, ультраструктурные особенности клеток, набор органоидов и плотность межклеточных контактов способствуют этому процессу. Из кровяного русла капилляров ресничных отростков в переднюю камеру ионы и вещества небольшой молекулярной массы проходят благодаря функционированию энергозависимой транспортной системы. В соответствии с Cole непигментированные клетки ресничного эпителия выборочно поглощают ионы натрия из стромы ресничного тела и транспортируют их через межклеточные пространства. Гиперосмотичность, возникающая в межклеточных пространствах в результате этого процесса, приводит к осмотическому потоку воды первоначально из стромы в межклеточные пространства, а затем в камерную влагу. О правильности подобного предположения свидетельствует обнаружение в мембранах непигментированных клеток ресничного эпителия ряда ферментов, у частвующих в этом процессе, в частности активности Na+/К+ -ATO-a3bi. угольной ангидразы.
Подтверждением тому являются и данные, относительно того, что применение средств, ингибирующих активность Na+/К+ -АТФ-азы, уменьшает секрецию камерной влаги. Прохождение ионов хлорида, бикарбоната и калия также обеспечивается активным транспортом. Ряд веществ проходит через эпителий пассивно, благодаря перепаду градиента концентрации. Это относится к сахарам и аминокислотам.
Секреция бикарбонатов снижается при применении ингибиторов угольной ангидразы (карбоангидраза), что и используют при глаукоме для снижения секреции камерной влаги. Такими же свойствами обладают бета-блокаторы.
Беспигментные эпителиоциты ресничного тела, помимо секреции камерной влаги, участвуют еще во многих процессах.
Они синтезируют компоненты стекловидного тела — коллагены различного типа и глюкуроновую кислоту. Участвуют эти клетки и в синтезе неколлагеновых белков — оптицина, фибулина-1 и нидогена-1, играющих немаловажнуто роль в стабилизации геля стекловидного тела и организации базальных мембран.
Помимо основных секретирующих функций беспигментный эпителий принимает участие во многих репаративных и адаптационных процессах. Так, совсем недавно установлено, что он способен ингибировать пролиферативную активность и функции T-лимфоцитов, регулируй тем самым иммунные механизмы внутри глазного яблока.
Пигментированный эпителий и наружная базальная мембрана (рис. 3.8.32—3.8.36). Как было указано выше, пигментный слой эпителия ресничного тела развивается из наружного слоя зрительного бокала. По направлению к задним отделам глаза он переходит в пигментный эпителий сетчатой оболочки, а по направлению кпереди в передний слой пигментного эпителия радужки.
Размер пигментированных эпителиальных клеток колеблется от 8—10 мкм в плоской части ресничного тела, до 10—15 мкм в ресничных отростках.
В цитоплазме клеток обнаруживаются крупные круглые зерна меланина (0,8—2,0 мкм, т. е. в 3—4 раза больше, чем те в клетках сосудистой оболочки). Видны также пучки микрофиламентов промежуточного типа. Цитоплазма клеток насыщена органоидами. Базальная мембрана пигментных эпителиоцитов образует многочисленные складки. Таким образом, микроскопически эти клетки практически не отличаются от клеток пигментного эпителия сетчатки.
Увеальная (мезодермальная) часть. Увеальная часть ресничного тела имеет сложное строение и состоит из сосудов, мышечной ткани и стромы. К увеальной части относится также наружная базальная мембрана. На перечисленных структурных элементах мы сейчас остановимся более подробно.
Слой сосудов увеальной части ресничного тела является продолжением сосудистого слоя хориоидеи. Между ним и базальной мембраной эпителия лежит слой коллагеновых волокон большого диаметра. По мере продвижения кпереди сосудистая ткань как бы собирается в складки, формируя соединительнотканную основу ресничных отростков.
Строма ресничного тела состоит из волокон, клеток и межклеточного вещества. Основными типами клеток стромы являются фибробласт и меланоцит. Обнаруживаются также моноциты, макрофаги, лимфоциты и дендритические клетки.
В последние годы обращено особое внимание на изучение функции моноцитов, макрофагов и дендритических клеток. Это связано с тем, что названные клетки определяют иммунную резистентность тканей глаза. Именно макрофаги сталкиваются с чужеродным антигеном, распознают его и передают информацию другим элементам иммунной системы.
Как и макрофаги других органов и тканей, макрофаги ресничного тела обладают необходимым набором рецепторов опознания чужеродного антигена, что показано при использовании методов иммуноморфологии . Более того, при воспроизведении экспериментального увеита или возникновении заболевания у человека в моноцитах и макрофагах ресничного тела и сосудистой оболочки происходит вся цепь структурных и функциональных изменений, свойственных макрофагам других локализаций при встрече с чужеродным антигеном.
Основной объем ресничного тела занимает соединительнотканная строма. В зависимости от расположения различают несколько слоев.
Внутренний слой соединительной ткани находится между мышцей и базальной мембраной пигментного эпителия. Этот слой довольно тонкий в плоской части ресничного тела, но утолщается в отростчатой части. В поверхностных слоях коллагеновые волокна этого слоя смешиваются с волокнами более плотной соединительной ткани ресничной мышцы. По направлению кпереди этот волокнистый слой переходит в строму радужной оболочки.
Передний слой соединительной ткани ресничного тела располагается на границе между ресничной мышцей и передней камерой глаза (рис. 3.8.39).

Рис. 3.8.39. Меридианальный срез ресничного тела: 1 — продольный пучок ресничной мышцы, прикрепляющейся к склеральной шпоре; 2— склеральная шпора; 3— циркулярный пучок ресничной мышцы; 4 — угол передней камеры; 5 — сосуд большого круга кровообращения радужной оболочки; 6 — радиальный пучок ресничной мышцы; 7 — пигментный эпителий ресничного тела
Строма ресничного тела несколько отличается от плотной неоформленной соединительной ткани, обнаруживаемой в других местах организма человека. Эти отличия сводятся, в первую очередь, к структуре эластических волокон. Эластические волокна на поперечном разрезе имеют вид трубочек, напоминая при этом волокна зонулярного аппарата. При этом эластические волокна формируют довольно густую сеть, в которую и помещены мышечные волокна. Иммуногистохимически выявляется большое количество микрофибриллярного белка фибриллина.
Определенные отличия выявлены и в отношении преобладающего типа коллагена, из которого состоят коллагеновые волокна стромы. Наиболее распространен коллаген VI типа, хотя выявляются и коллагены других типов. Коллаген VI типа преимущественно локализуется вокруг капиллярных сосудов. При этом количество данного типа коллагена больше со стороны, обращенной к эпителию. Коллаген VI типа образует также оболочки передних сухожилий ресничной мышцы, направляющихся к трабекулярной сети.
Коллагеновые и эластические волокна окутывают мышечные волокна, образуя межмышечную соединительную ткань. Именно указанное сочетание мышечных, коллагеновых и эластических волокон образует жесткую систему, передающую силу, возникающую при сокращении мышечных клеток, так называемым «сухожилиям», распространяющимся в ресничном теле особым образом (см. ниже).
В заключение необходимо указать на значительное количество в строме ресничного тела нервных стволов и кровеносных сосудов различного калибра (рис. 3.8.33, 3.8.37, 3.8.38).
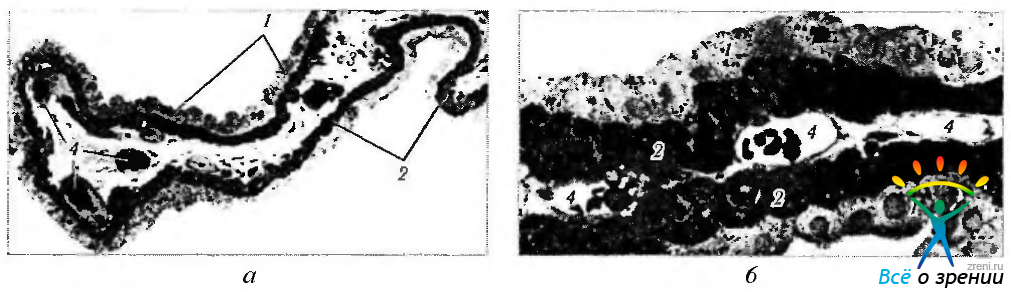
Рис. 3.8.37. Ресничный отросток (а — малое увеличение; б — большое увеличение): 1 — непигментированный ресничный эпителий; 2—пигментированный ресничный эпителий; 3— строма; 4— капиллярные сосуды, часть которых прилежит непосредственно к ресничному эпителию
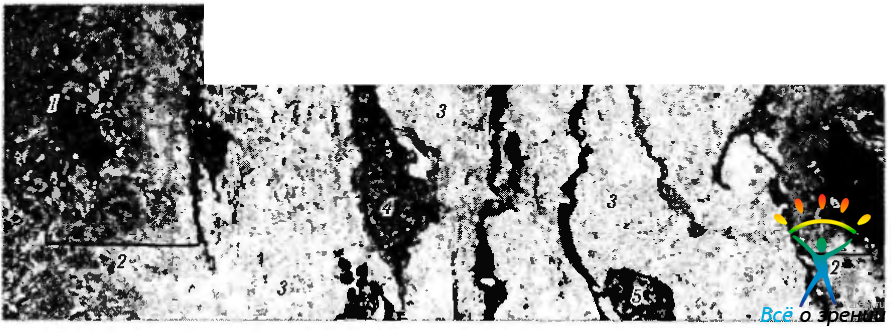
Рис. 3.8.38. Строма ресничного отростка ресничного кружка (по Hogan, 1972): базальная часть пигментного эпителия видна слева (/) и прилежит к толстой базальной мембране (2). Стромальный коллаген (3) состоит из плотных пучков коллагеновых волокон, ориентированных в различных направлениях. Видны также фибробласты (4) и нервные волокна (5)
Мембрана Бруха ресничного тела. Котикулярный слой мембраны Бруха продолжается на ресничное тело в виде базальной мембраны пигментного эпителия. Эластическая и коллагеновая части мембраны Бруха в области сосудистой оболочки отделяются от базальной мембраны слоем коллагена и. в конечном счете, исчезают в строме ресничного тела вблизи плоской части.
Ресничная мышца (т. ciliaris) (рис. 3.8.39— 3.8.42).
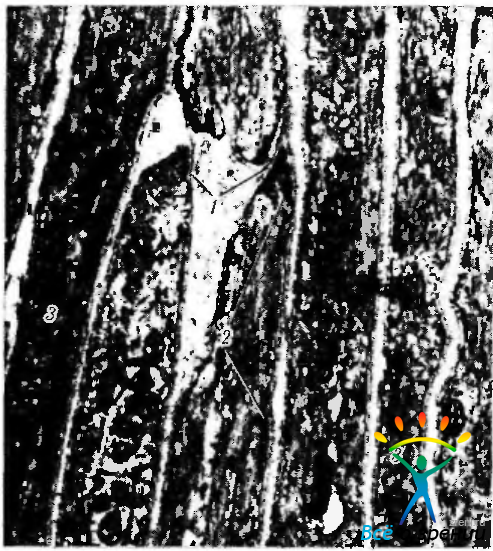
Рис. 3.8.40. Электроннограмма продольного среза ресничной мышцы (по Hogan et ah, 1971): 1 — базальная мембрана, окружающая мышечные клетки; 2 — десмосомоподобные уплотнения, расположенные на внутренней поверхности цитоплазматической мембраны мышечных клеток; 3 — миофиламенты
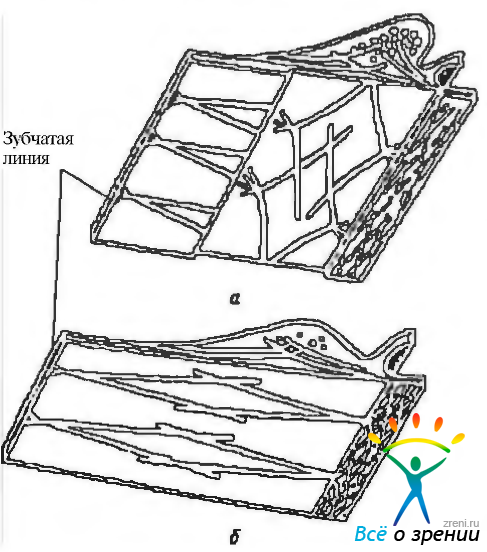
Рис. 3.8.41. Деформация ресничного тела и подтягивание кпереди зубчатой линии при сокращении ресничной мышцы: а — сокращение ресничной мышцы; б — расслабление ресничной мышцы
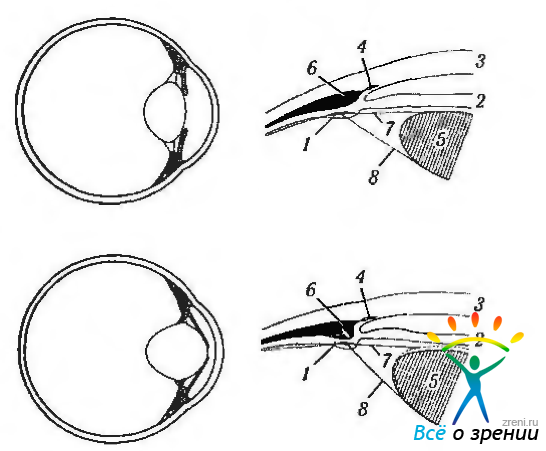
Рис. 3.8.42. Схема изменения геометрии глаза при расслаблении ресничной мышцы (о, б) и аккомодации (в, г) (по Rohen, 1979): 1 - система фибрилл зонулярного аппарата; 2 - радужная оболочка; 3 — роговица; 4 — шлеммов канал; 5 — хрусталик; 6 — ресничная мышца; 7 — волокна передней части цинновой связки; 8 — волокна задней части цинновой связки. Стрелка указывает направление движения ресничной мышцы в процессе аккомодации. Ресничная мышца при сокращении смещает внутренний край ресничного тела по направлению экватора хрусталика. При этом волокна передней цинновой связки расслабляются и хрусталик принимает более сферическую форму (пунктирная линия). Сосудистая оболочка подтягивается к центру и кпереди
Особое место, как в структурном, так и функциональном отношениях, занимает реcничная мышца. Именно благодаря деятельности мышцы осуществляется аккомодация глазного яблока.
Гладкомышечные волокна ресничного тела располагаются таким образом, что при своем сокращении в трех направлениях происходит деформация ресничного тела. Соответственно различают три группы волокон:
- Наружные меридианальные (продольные) пучки волокон (fibrcte meridionales) (мышца Брюкке), составляют большую часть ресничной мышцы. Начинаются они в подсосудистой пластинке вблизи ресничного тела и проходят до склеральной шпоры, прикрепляясь над трабекулярной сеточкой.
- Радиальные пучки волокон (fibrcte radiates), лежащие кнутри меридианальных. Направляются они изнутри кнаружи, пересекая под прямым углом меридианальные и циркулярные волокна ресничной мышцы Распространяясь назад, они прикрепляются на широком участке к соединительной ткани сосудистой оболочки.
- Циркулярные пучки волокон (fibrae radiates) (мышца Мюллера). Лежат такие волокна у внутреннего края ресничного тела вблизи его основания, кнутри от меридианальных волокон. Волокна циркулярно окружают ресничное тело.
Основная масса ресничной мышцы располагается в передних 2/3 ресничного тела. Сокращение среднего и наружного слоев ресничной мышцы в наибольшей степени приводит к смещению ресничных отростков кпереди и кнутри и расслаблению ресничного пояска.
Мышечные волокна ресничного тела, хотя и относятся к гладкомышечным волокнам, тем не менее обладают и определенными особенностями строения. Их цитоплазма содержит большое количество митохондрий. Исключительно хорошо развита эндоплазматическая сеть. Аппарат Гольджи находится в активном функциональном состоянии.
Определенные отличия от гладкомышечной ткани других органов существуют и в характере связи между отдельными мышечными клетками. Мышечные клетки ресничного тела складываются в так называемые «связки», окруженные тонким слоем фибробластов (рис. 3.8.39, 3.8.40). В определенных точках мышечные клетки прикрепляются друг к другу при помощи десмосом. Группы мышечных клеток окружены тонким слоем коллагеновой ткани и перимизием (рис. 3.8.40).
Как и все гладкомышечные клетки организма человека, миоциты ресничного тела содержат миофиламенты, преимущественно расположенные по периферии клетки. При этом основной особенностью миофиламентов является обнаружение в них структурных признаков поперечнополосатой мышцы, поскольку они располагаются практически параллельно друг другу и прикрепляются к электронноплотным структурам цитоплазматической мембраны, напоминающим Z-связки.
Необходимо отметить и то, что, как и поперечнополосатая мышца, ресничная мышца исключительно хорошо иннервирована.
В этом разделе уместно остановиться и на основной функции ресничной мышцы, а именно на ее участии в процессе аккомодации.
Аккомодация является процессом фокусировки изображения на сетчатую оболочку, а дисаккомодация представляет собой обратный процесс (расфокусировка). Основную роль при этом играет способность хрусталика к обратимой деформации в процессе сокращения и расслабления ресничной мышцы и сопровождающие этот процесс расслабление и натяжение цинновой связки хрусталика. Именно поэтому происходит деформация хрусталика.
Аккомодация сопровождается сужением зрачка, смещением радужки кпереди, увеличением кривизны передней и, в меньшей степени, задней поверхностей хрусталика, увеличением толщины хрусталика.
Вопросы функции ресничной мышцы в процессе аккомодации наиболее полно изучались у обезьян. Существуют доказательства того, что выявленные у обезьян механизмы аккомодации распространяются и на человека. Первоначально необходимо остановиться на особенностях взаимоотношения ресничной мышцы с окружающими структурами и, в частности, с хрусталиком.
Целесообразно начать с данных, относительно особенностей прикрепления «сухожилий» ресничных мышц. При этом различают передние и задние «сухожилия».
Передние «сухожилия», начинаясь от мышц, расходятся в виде веера, разделяясь на три части. Одна часть сухожильных волокон прикрепляется к передней части склеры, вторая — к склеральной шпоре, а третья — вплетается в волокнистую часть трабекулярного аппарата.
Задние «сухожилия» ресничной мышцы прикрепляются в другом месте, а именно в области плоской части ресничного тела. В отличие от передних «сухожилий» задние «сухожилия» содержат большое количество эластической ткани и вплетаются в адвентицию кровеносных сосудов, эластический слой мембраны Бруха ресничного тела, а также базальную мембрану ресничного эпителия.
Вышеприведенные особенности прикрепления передних и задних «сухожилий» создают систему, которая при сокращении или расслаблении ресничной мышцы быстро приводит к деформации и смещению ресничного тела в различных направлениях.
При синхронном сокращении всех частей мышцы диаметр «кольца» ресничного тела уменьшается. При этом внутренняя граница мышцы перемещается к хрусталику. Уменьшение диаметра «мышечного кольца» приводит к расслаблению цинновой связки, снижая напряжение капсулы хрусталика. В результате своей упругости, хрусталик изменяет форму, становясь более выпуклым (рис. 3.8.41). Передний полюс хрусталика при этом перемещается вперед, а задний остается на месте или слегка перемещается назад. Эти изменения конфигурации хрусталика и его перемещение и увеличивают силу рефракции.
Необходимо отметить, что в процессе аккомодации не изменяется кортикальная толщина хрусталика, но увеличивается сагиттальная толщина ядра. Именно благодаря этому происходит утолщение хрусталика в целом.
Много исследований было посвящено выяснению причин различной степени изменения кривизны передней и задней поверхностей хрусталика. Известно, что места прикрепления передних и задних зонулярных волокон цинновой связки к капсуле хрусталика различны. Поэтому сила напряжения зонулярных волокон направлена радиально от ресничного тела в направлении разветвления связки хрусталика в плоскости экватора (рис. 3.8.42). С этим и связывают различия в степени изменения кривизны передней и задней поверхностей хрусталика.
Другие исследователи считают, что различная степень изменения кривизны передней и задней поверхностей хрусталика связана с неодинаковой толщиной капсулы хрусталика в различных местах.
Coleman предложил «гидравлическую теорию». По его мнению, меньшее изменение кривизны задней поверхности хрусталика при аккомодации связано с наличием давления на него стекловидного тела. Тем не менее Fisher предполагает, что стекловидное тело не влияет на этот процесс. По его мнению, способность хрусталика к деформации всецело зависит от его физических свойств. Причем способность к деформации существенно отличается в центральной и экваториальной плоскостях хрусталика.
При сокращении ресничной мышцы происходит и ряд других структурных изменений в переднем отделе глаза. Так, сокращение части мышцы, прикрепляющейся к склеральной шпоре, приводит к расширению межтрабекулярных пространств, что способствует усилению фильтрации камерной влаги. Способствует этому процессу и то, что в склеральной шпоре обнаружены сократительные клетки — миофибробласты.
Необходимо еще остановиться и на возрастных изменениях структуры ресничной мышцы, видимо, играющих определенную роль в развитии довольно распространенного нарушения рефракции, называемого пресбиопией.
Пресбиопией называется состояние, характеризующееся снижением объема или величины аккомодации, развивающееся с возрастом и сопровождающееся уменьшением ближайшей точки ясного зрения. При этом точка дальнего видения не изменяется. Уменьшается также скорость (время) аккомодации и дисаккомодации.
Объем аккомодации изменяется в довольно широких пределах. Так, у молодых людей он равен 10—12 диоптрий и уменьшается к 50 годам до 2 диоптрий.
На протяжении многих десятилетий рассматриваются две причины пресбиопии. Это изменение функциональной активности ресничной мышцы и изменение упругости хрусталика. Первоначально мы остановимся на значении в этом процессе ресничной мышцы.
Начиная с первого месяца жизни и на протяжении всего первого десятилетия, количество волокон ресничной мышцы увеличивается. Начиная с 10-летнего возраста вплоть до 60 лет, нарастает количество соединительной ткани с прогрессивным замещением волокон особенно в задних отделах мышцы. По этой причине мышца постепенно утолщается в передней своей части. Выявлено также прогрессирующее уменьшение длины меридианальной части мышцы. После 60 лет продольные и радиальные части мышцы атрофируются, в то время как масса циркулярной части увеличивается. Важно отметить, что несмотря на постоянно протекающий процесс уменьшения объема мышцы и количества мышечных волокон, сила мышечного сокращения с возрастом увеличивается. Так, по данным Fisher, в возрасте 50 лет ресничная мышца на 50% мощнее, чем у молодых. Несмотря на это, скорость аккомодации ниже.
Именно по этой причине ряд исследователей считают, что более вероятной и основной причиной пресбиопии является склероз задних «сухожилий» ресничной мышцы. Это связывают с тем, что склероз задних «сухожилий» ограничивает переднее внутреннее смещение ресничной мышцы. Именно смещение ресничной мышцы в указанном направлении абсолютно необходимо для достижения расслабления зонулярного аппарата. Показано, что степень склеротических изменений коррелирует с выраженностью изменения объема аккомодации, уменьшением реакции мышцы на пилокарпин. При этом каких-либо существенных изменений плотности нервных окончаний в ресничной мышце не обнаруживается.
Вторая теория пресбиопии основной причиной ее развития считает изменения эластичности хрусталика. Так, еще в 1855 г. Helmholtz считал, что пресбиопия развивается в результате недостаточной способности хрусталика к деформации. Действительно, с возрастом хрусталик увеличивается в объеме, увеличивается его масса и уменьшается эластичность.
Таким образом, на настоящий момент времени целесообразно рассматривать как равноценные обе теории развития пресбиопии.
Кровоснабжение ресничного тела (рис. 3.8.17, 3.8.43—3.8.47).
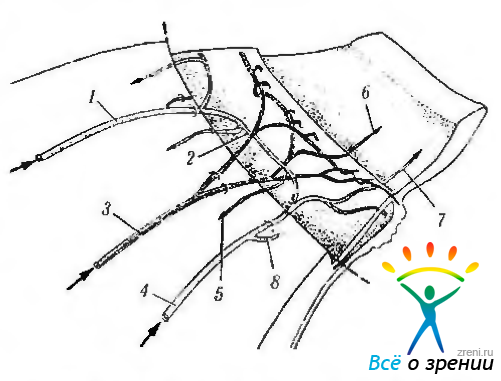
Рис. 3.8.43. Схематическое изображение архитектуры крупных сосудов переднего отдела глаза (по Funk, Rohen, 1990): вид снаружи. Лимбальная область и склера удалены для показа сосудов ресничного тела и радужки (1 — передняя ресничная артерия; 2 — интрамуральный круг кровообращения; 3 — задняя длинная ресничная артерия; 4 — передняя ресничная артерия; 5—возвратная хориоидальная артерия; 6—прямые артерии радужки, исходящие из задней длинной ресничной артерии; 7 -прямые артерии радужки, исходящие из передней ресничной артерии; 8—интерсклеральная лимбальная артерия)
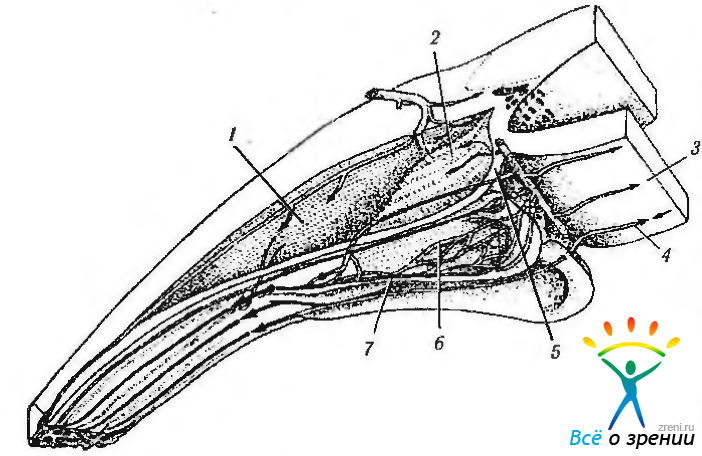
Рис. 3.8.44. Схематическое изображение сосудистой сети ресничного тела человека (по Funk, Rohen, 1990) (объяснения в тексте): 1 — наружная задняя часть; 2 — внутрення передняя зона; 3 — артерии радужки; 4 — венулы радужки; 5 — первая сосудистая территория; 6—вторая сосудистая территория; 7 — третья сосудистая территория

Рис. 3.8.45. Схема сосудистой системы ресничных отростков (вид сбоку) (по Morrison, Bushirk, 1984)
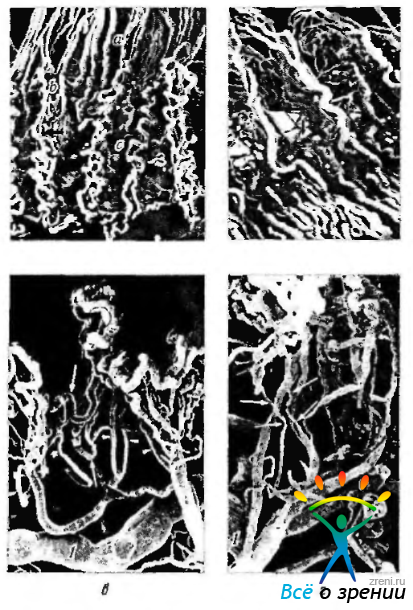
Рис. 3.8.46. Сканирующая электронная микроскопия сосудистой сети ресничного тела (по Funk, Rohen, 1990): а — вид внутренней поверхности. Строение первой (а), второй (b) и третьей (с) сосудистых территорий (стрелки указывают на краевые венулы внутреннего края ресничных отростков); б— вид сбоку Краевая венула внутреннего края ресничных отростков (стрелка указывает на венулу третьей сосудистой территории); в, г — вид спереди. Круг расположен в области большого ресничного отростка (1 — большой артериальный круг радужки; стрелка — первая сосудистая территория; * — венула этой территории)

Рис. 3.8.47. Распределение кровеносных сосудов в ресничном теле и радужной оболочке: инъекция сосудистого русла китайской тушью. Определяется крупный артериальный ствол, относящийся к большому кругу кровообращения радужной оболочки, и множество капиллярных сосудов ресничного тела и радужной оболочки
Сосудистое сплетение отростков ресничного тела (рис. 3.8.43—3.8.45). Артериолы ресничных отростков исходят из большого круга кровообращения радужки и переходят в широкие венулы.
Капилляры находятся в плотном контакте с базальной мембраной клеток пигментного эпителия (рис. 3.8.37). Ширина капилляров приближается к ширине вен и напоминает хо-риокапилляры сосудистой оболочки. Диаметр их равен 15—30 мкм, стенка фенестрирована (30—100 нм) и проницаема для воды и белков плазмы. Специализированные соединения по строению идентичны соединениям, обнаруживаемым в капиллярах сосудистой оболочки.
Капилляры, снабжающие ресничную мышцу, встречаются реже. Их диаметр меньше, а эндотелиальные клетки толще. Капилляры постепенно переходят в венулы.
Высокая пропускная способность стенок капилляров ресничного тела позволяет белкам плазмы свободно диффундировать и распределяться в строме ресничного тела, а затем и в строме радужки. Поскольку строма ресничного тела примыкает к передней камере глаза, становится понятным, почему белки плазмы обнаруживаются и в камерной влаге, правда, в небольших концентрациях.
Различают три территории кровоснабжения (рис. 3.8.43—3.8.47).
Первая сосудистая территория находится в передней глубокой части гребня каждого большого ресничного отростка, состоит она из артериол, формирующих капиллярную сеть. Венулы разворачиваются и идут к основанию отростков (рис. 3.8.44).
Вторая сосудистая территория обнаруживается в передней части больших отростков и состоит из двух компонентов. Основная капиллярная сеть находится центрально и в глубине ресничного отростка, а кровь из нее оттекает в более поверхностно расположенную капиллярную сеть, а оттуда в венулы. Поверхностная капиллярная сеть — образует почти прямую связь между артериолой и большой краевой венулой, расположенной сагиттально во внутреннем крае ресничного отростка и формирующей отводящий венозный сегмент. Она впадает в венулу плоской части ресничного тела. Третья сосудистая территория состоит из капиллярных сетей маленьких ресничных отростков и сосудов задней трети больших отростков. Кровь оттекает в краевую (маргинальную) венулу и, частично, в базальную.
В ресничных отростках человека имеется еще один путь оттока, предлагающий быстрое отведение крови с высокой венозной концентрацией кислорода и высоким венозным давлением. Предполагают, что эта система участвует в механизмах фильтрации камерной влаги.
Кровоснабжение ресничной мышцы. Передняя и внутренняя части ресничной мышцы обеспечиваются кровью большим крутом кровообращения радужки, в то время как внешняя и задняя части — внутримышечным крутом кровообращения ресничного тела. Большой крут сформирован в основном длинными задними ресничными артериями, в то время как внутримышечный крут — ветвями передних ресничных артерий. Эти две системы анастомозируют между собой (рис. 3.8.43).
Иннервация ресничного тела. Задние длинные ресничные нервы отдают первые ветви в наружном слое переднего отдела сосудистой оболочки, где формируется мощное сплетение миелинизированных и немиелинизированных нервных стволов, сопровождающееся многочисленными ганглиозными клетками. Часть ганглиозных клеток лежит и среди мышечных волокон, а также вдоль внутреннего тела ресничной мышцы. Ганглиозные клетки отдают многочисленные цитоплазматические отростки, распределяющиеся в окружающих тканях ресничного тела и стромы радужной оболочки.
Парасимпатические волокна. Парасимпатические волокна, берущие свое начало в ядре Якубовича—Эдингера—Вестфаля, подходят к глазному яблоку вместе с ветвями глазодвигательного нерва. Эти волокна смешанные. Тела большинства нейронов располагаются в ресничном ганглии. Эти волокна образу ют обширное сплетение, расположенное в пределах ресничной мышцы. Парасимпатические волокна иннервируют сфинктер и ресничную мышцу.
Эктопически расположенные ганглиозные клетки обнаруживаются в области сплетения ресничного тела, вдоль задних длинных ресничных нервов, а также между ресничным ганглием и глазным яблоком.
Симпатические волокна. Симпатические волокна берут свое начало в шейном симпатическом узле. Эти волокна подходят к ресничному телу и радужке посредством длинных ресничных нервов. Симпатические волокна, сопровождающие ресничные артерии, довольно широко распределены в пределах ресничных сплетений.
Сенсорные волокна. Свое начало сенсорные волокна берут от носоресничного нерва (n. nаsociliaris). Эти волокна поступают в ресничное тело и заканчиваются в радужке, роговице и ресничной мышце.
У 12% людей в области склерального канала обнаруживаются петли ресничного нерва (интрасклеральная петля нерва Аксенфельда (Axenfeld)). Размер их 1—2 мм. Это происходит в месте перфорации склеры передними ресничными артериями.
Нервные волокна проходят от сплетения между склерой и стромой ресничного тела и формируют обширное сплетение в пределах ресничной мышцы. В дальнейшем от этого сплетения отходят волокна и образуют еще одно сплетение, иннервирующее ресничный эпителий.
Терминалы нервов видны как вблизи клеток пигментного эпителия, так и капилляров ресничного тела. Некоторые терминалы относятся к парасимпатической нервной системе, а некоторые к адренэргическим.
Иннервация ресничной мышцы. Ресничная мышца иннервируется исключительно большим количеством нервных волокон. Каждая отдельная мышечная клетка окружена примерно до 10—15 нервными окончаниями, ширина которых 0,5—1,0 мкм. Эти окончания специфически окрашиваются на синаптофизин. Волокна начинаются в нейронах ядра Якубовича—Эдингера—Вестфаля и образуют синапсы в ресничном ганглии. Плотность мускариновых и холинэргических окончаний нервов значительно больше, чем в других тканях.
Симпатическая иннервация имеет меньшее физиологическое значение. В терминалах нервов выявлены нитрэргические и пептидэргические нейротрансмиттеры.
В дополнение к вегетативной иннервации в области ресничной мышцы определяется также скопление ганглиозных клеток (plexus gangliosus ciliaris).
Лежат скопления ганглиоцитов между связками продольной и циркулярной частей ресничной мышцы. Они обычно располагаются изолированно и очень редко образуют группу из 2—3 клеток.
Различают маленькие ганглиозные клетки (70%), диаметр которых равняется 10—14 мкм, и большие (30%), с диаметром 30 мкм. Ганглиозные клетки ресничного тела меньше, чем ганглиозные клетки других органов.
Ганглиозные клетки и аксоны относятся к нитрэргическим и положительно окрашиваются при проведении реакции на НАДФ-диафоразу.
Функция ганглиозных клеток ресничного тела еще не полностью понятна. Нитрэргические волокна нервного сплетения, возможно, служат для расслабления ресничной мышцы, обеспечивая дисаккомодацию. Предполагают, что активное расслабление вносит вклад в аккомодацию.
Определяется периваскулярная сеть из нитрэргических волокон в пределах круглой части ресничной мышцы, связанных с ганглиозным сплетением. Пептидэргические нейроны также найдены в тройничном ганглии.
Продолжение в следующей статье: Сосуды и сосудистая оболочка глазного яблока ? Часть 3
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0