Анатомия головного мозга │ Часть 1


Описание
Все функции глазного яблока, его придаточного аппарата и глазницы контролируются многими отделами головного мозга. В этой связи необходимо хотя бы кратко остановиться на анатомическом строении и функциях структур головного мозга, обеспечивающих функцию зрительного анализатора.Центральная нервная система складывается из головного и спинного мозга. Головной мозг условно разделяют на большой мозг (cerebrum), малый мозг (cerebellum) (табл. 4.1.1, рис. 4.1.1).

Таблица 4.1.1. Структуры головного мозга
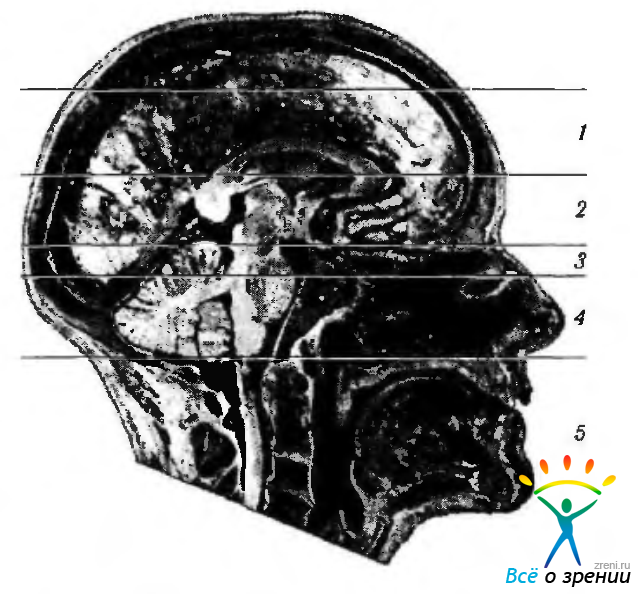
Рис. 4.1.1. Среднесагиттальный разрез головного мозга. Отделы головного мозга (ядерный парамагнитный резонанс): 1 — конечный мозг; 2 — промежуточный мозг; 3 — задний мозг; 4 — продолговатый мозг; 5 — спинной мозг
На основании особенностей эмбрионального развития головной мозг можно разделить на следующие отделы, располагающиеся, начиная с каудального конца, в таком порядке:
- Ромбовидный (rhombencephalon), или задний, мозг, который, в свою очередь, состоит из:
а) продолговатого мозга fmyelencephalon);
б) собственно заднего мозга (metencepha-Ion). - Средний мозг (mesencephalon).
- Передний мозг (prosencephalon), в кото ром различают:
а) промежуточный мозг (diencephalon);
б) конечный мозг (telencephalon).
Все названные отделы, кроме мозжечка и конечного мозга, составляют ствол мозга.
Кроме этих отделов, выделяют еще перешеек (istmus rhombensephali), расположенный между задним и средним мозгом.
Некоторые обобщенные сведения об анатомических структурах головного мозга, могущие помочь читателю в дальнейшем, приведены в табл. 4.1.2.
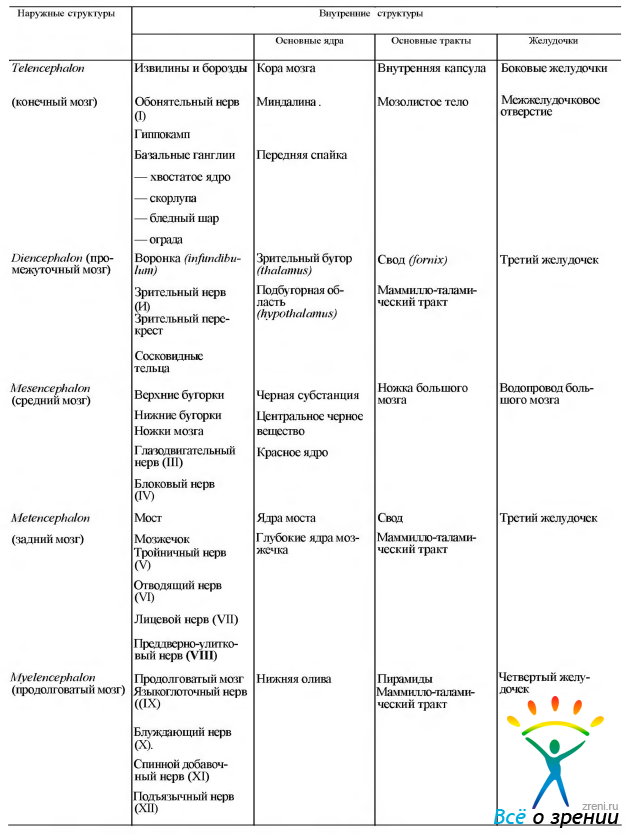
Таблица 4.1.2. Отделы мозга
Целью настоящего раздела является, в первую очередь, напомнить основные анатомические образования мозга, принимающие участие в столь сложном акте, как зрительное восприятие.
Конечный (концевой) мозг
Конечный мозг (telencephalon) представлен двумя полушариями (hemispheria cerebri). В состав каждого полушария входят: плащ, или мантия (pallium), обонятельный мозг (rhinencephalon) и базальные ганглии (узлы). Остатком первоначальных полостей обоих пузырей конечного мозга являются боковые желудочки (ventriculi lateralis).
Передний мозг, из которого выделяется конечный, вначале возникает в связи с обонятельным рецептором, а затем он становится органом управления поведением животного. При этом в нем возникают центры инстинктивного поведения — подкорковые ядра и центры индивидуального поведения, основанного на индивидуальном опыте, — кора большого мозга. Соответственно этому, в конечном мозге различают в порядке исторического развития следующие группы центров:
- Обонятельный мозг (rhinencephalon) — самая древняя и вместе с тем самая меньшая часть мозга, расположенная вентрально.
- Базальные ганглии, «подкорка» — старая часть конечного мозга (paleencephalon), скрытая в глубине.
- Серое вещество коры (cortex) — самая молодая часть и вместе с тем самая большая часть, покрывающая остальные как бы плащом, откуда и ее название плащ, или мантия (pallium).
Так как в процессе эволюции из всех отделов центральной нервной системы быстрее всего растет конечный мозг, то он у человека становится самой большой частью головного мозга и приобретает вид двух полушарий — правого и левого (hemisphera dextrum et sinistrum). В глубине продольной щели мозга оба полушария соединены между собой толстой горизонтальной пластинкой — мозолистым телом (corpus callosum), которое состоит из нервных волокон, идущих поперечно из одного полушария в другое.
Поверхность каждого полушария покрыта большим количеством различной глубины и протяженности борозд, между которыми располагаются извилины. На каждом полушарии различают следующие поверхности (рис. 4.1.2— 4.1.4):
- верхнелатеральную,
- медиальную
- и нижнюю.
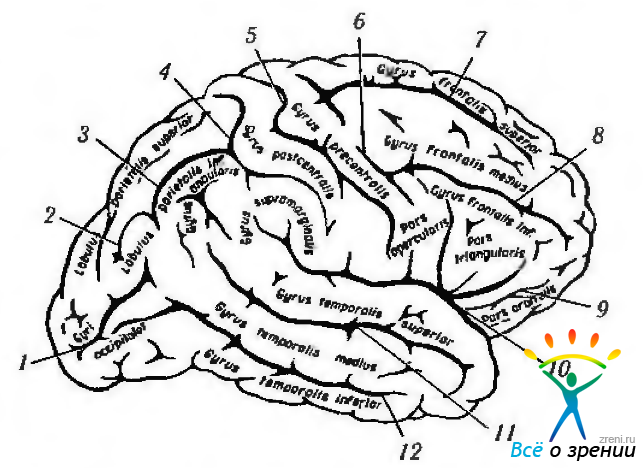
Рис. 4.1.2. Борозды и извилины головного мозга (верхне-боковой вид): 1—полулунная борозда: 2—поперечная затылочная борозда: 3—внутритеменная борозда; 4—постцентральная борозда; 5 — центральная борозда; 6 — предцентральная борозда; 7 — верхняя лобная борозда; 8 — нижняя лобная борозда; 9 — передняя ветвь; 10 — латеральная борозда; 11 — верхняя височная борозда; 12 — нижняя височная борозда

Рис. 4.1.3. Борозды и извилины мозга (медиальная поверхность): 1—медиальная лобная извилина; 2 - поясная борозда; 3—борозда мозолистого тела; 4 — подтеменная борозда, 5 — теменно-затылочная борозда; 6 — шпорная борозда; 7 — затылочный полюс
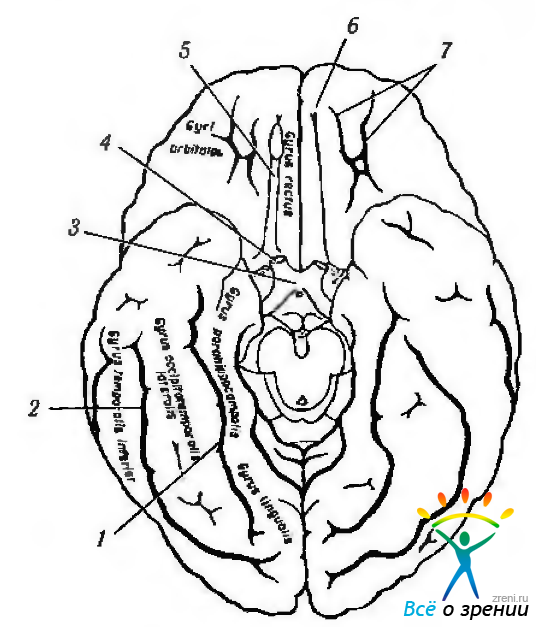
Рис. 4.1.4. Борозды и извилины мозга (вид снизу): 1 — коллатеральная борозда; 2 — затылчно-височная извилина; 3 — зрительный перекрест; 4 — зрительный нерв; 5 — обонятельный тракт; 6 - обонятельная борозда; 7 — глазничные борозды
Выделяют три края: верхний, нижний и медиальный и полюса: передний (polus frontalis), задний (polus occipitalis) и височный (polus temporalis).
Поверхность полушария (плащ) образована равномерным слоем серого вещества толщиной 1,3—4,5 мм, содержащего нервные клетки. Слой этот, называемый корой большого мозга (cortex cerebri), представляется как бы сложенным в складки, благодаря чему поверхность плаща имеет в высшей степени сложный рисунок, состоящий из чередующихся между собой в различных направлениях борозд и валиков между ними, называемых извилинами (gyri). Каждое полушарие бороздами разделяется на большие участки, называемые долями (lobi). Последние в свою очередь разделяются на дольки и извилины. Долей каждого полушария пять: лобная (lobus frontalis), теменная (lobus parietalis), височная (lobus temporalis), затылочная (lobus occipitalis) и долька, скрытая на дне латеральной борозды, так называемый островок (insula).
Верхнелатеральная поверхность полушария разграничена на доли посредством трех борозд: латеральной, центральной и верхнего конца теменно-затылочной борозды, которая находится на медиальной стороне полушария. Латеральная борозда (sulcus cerebri lateralis) начинается на базальной поверхности полушария из латеральной ямки и затем переходит на верхнелатеральную поверхность, направляясь назад и несколько вверх. Она оканчивается приблизительно на границе средней и задней третей верхнелатеральной поверхности полушария. В передней части латеральной борозды от нее отходят две небольшие ветви: ramus ascendens и ramus anterior, направляющиеся в лобную долю.
Центральная борозда (sulcus centralis) начинается на верхнем краю полушария, несколько кзади от его середины, и идет вперед и вниз. Нижний конец центральной борозды не доходит до латеральной борозды. Участок полушария, находящийся впереди центральной борозды, относится к лобной доле. Часть мозговой поверхности, лежащая сзади от центральной борозды, составляет теменную долю, которая, посредством задней части латеральной борозы, отграничивается от лежащей ниже височной доли. Задней границей теменной доли служит конец вышеупомянутой теменно-затылочной борозды (sulcus parietooccipitalis), расположенной на медиальной поверхности полушария.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Эта граница неполная, поскольку названная борозда не заходит далеко на верхнелатеральную поверхность. Вследствие этого теменная доля непосредственно переходит в затылочную долю. Затылочная доля также не имеет четкой границы с лежащей впереди височной долей. Граница между ними проводится искусственно. Идет эта граница по линии от теменно-затылочной борозды к нижнему краю полушария.
Каждая доля состоит из ряда извилин, называемых в отдельных местах дольками, которые ограничиваются бороздами мозговой поверхности.
Лобная доля. В заднем отделе наружной поверхности этой доли проходит прецентральная борозда (sulcus precentralis), лежащая практически параллельно центральной борозде (sulcus centralis). От нее в продольном направлении отходят две борозды: верхняя и нижняя лобные борозды (sulcus frontalis superior et sulcus frontalis inferior). Благодаря этому лобная доля разделяется на четыре извилины — одну вертикальную и три горизонтальные. Вертикальная извилина (gyrus precentralis) находится между центральной и прецентральной бороздами.
Горизонтальные извилины лобной доли следующие:
- верхняя лобная извилина (gyrus frontalis superior), идущая выше верхней лобной борозды и параллельно верхнему краю полушария. При этом она заходит на медиальную поверхность полушария;
- средняя лобная извилина (gyrus frontalis medius) тянется между верхней и нижней лобными бороздами;
- нижняя лобная извилина (gyrus frontalis inferior) помещается между нижней лобной и латеральной бороздами. Ветви латеральной борозды, вдающиеся в нижнюю лобную извилину, делят последнюю на три части: pars opercularis, pars triangularis и, наконец, pars orbitalis.
Теменная доля. На теменной доле приблизительно параллельно центральной борозде располагается постцентральная борозда (sulcus postcentralis). Теменная доля разделяется на три извилины, из которых одна вертикальная, а две другие горизонтальные. Вертикальная извилина (gyrus postcentralis) идет позади центральной борозды в одном направлении с прецентральной извилиной (gyrus precentralis), отделенная от нее центральной бороздой. Выше sulcus intraprecentralis помещается верхняя теменная извилина, или долька (lobulus parietalis superior), которая распространяется и на медиальную поверхность полушария.
Ниже sulcus intraparietalis лежит нижняя теменная долька (lobulus parietalis inferior), которая, направляясь назад, огибает концы латеральной борозды и теряется в области затылочной доли. Часть нижней теменной дольки называется надкраевой извилиной (gyrus supramarginalis), а другая часть, огибающая верхнюю височную извилину (sulcus temporalis superior), носит название угловой извилины (gyrus angularis).
Височная доля. Латеральная поверхность этой доли имеет три продольные извилины, отграниченные друг от друга верхней и нижней височными извилинами. Верхняя из извилин (gyrus temporalis superior) находится между латеральной бороздой и верхней височной. Верхняя ее поверхность, скрытая в глубине латеральной борозды, несет 2—3 короткие извилинки, называемые поперечными височными извилинами (gyrus temporalis transversus). Между верхней и нижней височными бороздами распространяется средняя височная извилина (gyrus temporalis medius). Ниже последней проходит нижняя височная извилина (gyrus temporalis inferior).
Затылочная доля. Борозды латеральной поверхности этой доли изменчивы и непостоянны. Из них выделяют поперечную затылочную извилину (sulcus occipitalis transversus). соединяющуюся обычно с концом sulcus intraparietalis.
Островок (insula). Для того чтобы увидеть островок, необходимо раздвинуть или удалить нависающие над ним края латеральной борозды. Островок имеет форму треугольника, верхушка которого обращена вперед и вниз.
Нижняя поверхность полушария в той ее части, которая лежит кпереди от латеральной ямки, относится к лобной доле. Здесь параллельно медиальному краю полушария проходит обонятельная борозда (sulcus olfactorius), в которой лежат обонятельная луковица и обонятельный тракт (bulbus et tractus olfactorius). Между этой бороздой и медиальным краем полушария расположена прямая извилина (gyrus rectus), представляющая собой продолжение верхней лобной извилины. Латерально от обонятельной борозды на нижней поверхности находится несколько непостоянных бороздок, орбитальные борозды (sulci orbitales), ограничивающие орбитальные извилины (gyri orbitales).
Задний участок базальной поверхности полушария образован нижними поверхностями височной и затылочной долей, которые здесь не имеют определенных границ. На этом участке видны две борозды: латеральная затылочновисочная борозда (sulcus occipitotemporalis lateralis) и идущая параллельно ей коллатеральная борозда (sulcus collateralis). Между ними располагается медиальная затылочно-височная извилина (gyrus occipitotemporalis medialis). Медиально от коллатеральной борозды расположены две извилины: между задним отделом этой борозды и бороздой птичьей шпоры (sulcus calcarinus) лежит язычная извилина (gyrus lingualis), а также парагиппокампальная извилина (gyrus parahyppocampalis). Последняя извилина находится уже на медиальной поверхности полушария.
Медиальная поверхность полушария. На этой поверхности находится борозда мозолистого тела (sulcus corporis callosi). Параллельно и выше этой борозды проходит по медиальной поверхности полушария поясная борозда (sulcus cinguli). которая начинается спереди под клювом мозолистого тела, затем идет назад и оканчивается своим задним концом на верхнем краю полушария. Пространство, располагающееся между этим краем полушария и sulcus cinguli, относится к лобной доле, к верхней лобной извилине. Небольшой участок над sulcus cinguli называется парацентральной долькой (lobulus paracentralis), так как он соответствует медиальной поверхности верхних концов обеих центральных извилин, переходящих здесь друг в друга.
Кзади от парацентральной дольки находится четырехугольная поверхность (предклинье, рrеcuneus), ограниченная спереди концом sulcus cinguli, снизу небольшой sulcus subparietalis, а сзади глубокой sulcus parietoocipitalis. Предклинье относится к теменной доле. Позади предклинья лежит резко обособленный участок коры, относящийся к затылочной доле, — клин (cuneus), который ограничен спереди теменно-затылочной бороздой (sulcus parietoocipitalis), а сзади бороздой птичьей шпоры (sulcus calcarinus). Между sulcus cinguli и бороздой мозолистого тела протягивается поясная извилина (gyrus cinguli).
Поясная извилина, перешеек и парагиппокампальная извилины вместе образутот сводчатую извилину (gyrus fornicatus), которая описывает почти полный крут, открытый только снизу и спереди. Сводчатая извилина относится к лимбической системе. Лимбическая область (regio limbica) является частью новой коры полушарий большого мозга. Занимает она поясную и парагиппокампальную извилины.
Даже при поверхностном изучении мозга выявляются различия между полушариями мозга и стволом мозга. Ствол мозга и спинной мозг состоят из центрально расположенного скопления серого вещества (тела нервных клеток). Окружено серое вещество белым веществом (аксоны восходящих и нисходящих путей). В то же время в полушариях мозга серое вещество формирует наружный слой или кору, а внутренний состоит из белого вещества. Белое вещество представляет собой аксоны кортикальных нейронов, а также главные восходящие пути от зрительного бугра и других областей ствола мозга. Следовательно, термин «кора головного мозга» относится к серому веществу полушарий мозга.
Белое вещество полушарий. Все пространство между серым веществом мозговой коры и базальными ядрами занято белым веществом. Оно состоит из большого количества нервных волокон, идущих в различных направлениях и образующих проводящие пути конечного мозга. Внутренние слои белого вещества могут быть разделены на три системы волокон (рис. 4.1.5):
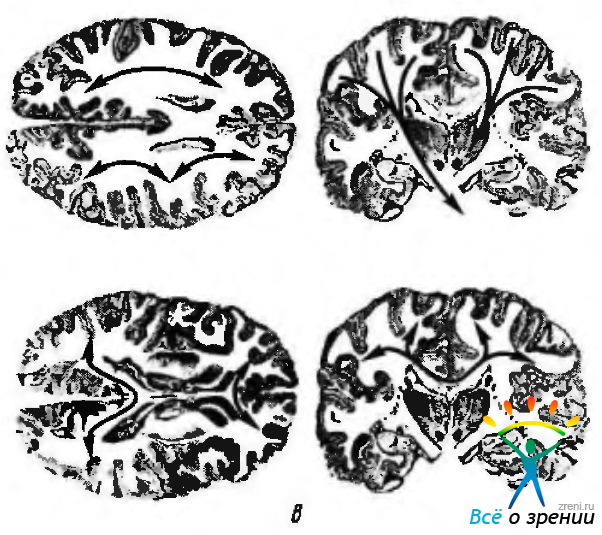
Рис. 4.1.5. Кортикальные проекции левого полушария. Схематическое изображение связей коры: а — ассоциативные связи; б — проекционные связи; в — комиссурные связи
- ассоциативные;
- комиссурные;
- проекционные.
Ассоциативные волокна связывают между собой различные участки коры одного и того же полушария. Они разделяются на короткие волокна и длинные. Короткие волокна (fibrae arcuatae cerebri) связывают между собой соседние извилины в форме дугообразных пучков. Данные ассоциативные волокна соединяют более отдаленные друг от друга участки коры. Таких пучков волокон существует несколько. Пояс (cyngulum) представляет собой пучок волокон, проходящий в извилину свода (gyrus fomicatus) и соединяющий различные участки коры как между собой, так и с соседними извилинами медиальной поверхности полушария. Лобная доля соединяется с нижней теменной долькой, затылочной долей и задней частью височной доли посредством верхнего продольного пучка (fasciculus longitudinalis superior). Височная и затылочная доли связываются между собой через нижний продольный пучок (fasciculus longitudinalis inferior). Наконец, орбитальную поверхность лобной доли соединяет с височным полюсом так называемый крючковидный пучок (fasciculus uncinatua).
Комиссурные волокна, входящие в состав так называемых спаек мозга, соединяют симметричные части обоих полушарий. Самая большая мозговая спайка — мозолистое тело (corpus callosum) — связывает между собой части обоих полушарий.
Две мозговые спайки, передняя и нижняя спайки мозга (comissura anterior и comissura inferior), гораздо меньшие по своим размерам, относятся к обонятельному мозгу (rhinencephalon).
Проекционные волокна можно разделить на следующие типы — восходящие и нисходящие. Восходящие приводящие связи, идущие от подкорковых структур, образуют лучистый венец (corona radiata) и состоят из аксонов нейронов, расположенных в ядрах зрительного бугра. Исключением является обонятельный путь, который проецируется непосредственно на обонятельную кору без формирования синапсов в зрительном бугре.
К нисходящим трактам от коры относятся — кортикоталамический (к зрительному бугру), кортикоспинальный (к ядрам спинного мозга), кортикобульбарный (к ядрам ствола мозга), котикопонтинный (к ядрам моста), кортикорубральный (к красному ядру) и кортикостриатныи (к базальным ганглиям).
Проекционные волокна в белом веществе полушария, расположенные ближе к коре, образуют лучистый венец, большая часть волокон которого сходится во внутренней капсуле.
Внутренняя капсула (capsula interna) представляет собой слой белого вещества между чечевицеобразным ядром (nucleus lentiformis), с одной стороны, и хвостатым ядром и зрительным бугром — с другой. На фронтальном разрезе мозга внутренняя капсула имеет вид косо идущей белой полосы, продолжающейся в ножку мозга. На горизонтальном разрезе она представляется в форме угла, открытого в латеральную сторону. Вследствие этого во внутренней капсуле различают переднюю ножку (crus anteriiis capsulae inteniae), лежащую между хвостатым ядром и передней половиной внутренней поверхности чечевицеобразного ядра, заднюю ножку (crus posterior), расположенную между зрительным бугром и задней половиной чечевицеобразного ядра, и колено (genu capsulae), лежащее на месте перегиба между обеими частями внутренней капсулы.
Проекционные волокна по их длине могут быть разделены на следующие системы, начиная с самых длинных волокон:
- Пирамидный путь (tractus corticospinalis (piramidis)) проводит двигательные болевые импульсы к мышцам туловища и конечностей. Начавшись от пирамидных клеток коры средней и верхней частей предцентральной извилины и парацентральной дольки (lobulus раrаcentralis), волокна пирамидного пути идут в составе лучистого венца, а затем проходят через внутреннюю капсулу, занимая передние две трети ее задней ножки, причем волокна для верхней конечности идут спереди волокон для нижней конечности. Далее они проходят через ножку мозга (pednnculus cerebri), а оттуда через мост — в продолговатый мозг.
- Корково-ядерный путь (tractus cortico-nuclearis) представляет собой комплекс проводящих путей к двигательным ядрам черепномозговых нервов. Начавшись от пирамидных клеток коры нижней части предцентральной извилины, они проходят через колено внутренней капсулы и через ножку мозга, затем вступают в мост и, переходя на другую сторону, оканчиваются в двигательных ядрах противоположной стороны, образуя перекрест. Небольшая часть волокон оканчивается без перекреста. Так как все двигательные волокна собраны на небольшом пространстве во внутренней капсуле (колено и передние две трети задней ножки ее), то при повреждении их в этом месте наблюдается односторонний паралич противоположной стороны тела.
- Корково-мостовой путь (tractus cortico-pontini) идет от мозговой коры к ядрам моста. Выделяются пути, направляющиеся от коры лобной доли (tractus frontopontimis). затылоч ной (tractus occipitopontimis), височной (tractus temporopontimis) и теменной (tractus parieto-pontinus). В качестве продолжения этих путей из ядер моста идут волокна в мозжечок в со ставе его средних ножек. При помощи этих путей кора большого мозга оказывает тормозное и регулирующее влияние на деятельность мозжечка.
- Таламо-кортикальные и корково-таламические волокна (fibrae thalamocorticalis et corticotalamici) направляются от таламуса к коре и обратно от коры к таламусу. Из волокон, идущих от таламуса, необходимо выделить так называемую центральную таламическую лучистость. Таламическая лучистость является конечной частью чувствительного пути, направляющегося к центру кожного чувства в постцентральную извилину. Выходя из латеральных ядер таламуса, волокна этого пути проходят через заднюю ножку внутренней капсулы позади пирамидного пути. Это место названо чувствительным перекрестом, так как здесь проходят и другие чувствительные пути, а именно: зрительная лучистость (radiacio optica) и слуховая лучистость (radiacio acustica).
Цитоархитектоника. Кора мозга довольно строго организована как в горизонтальной, так и вертикальной плоскостях. Горизонтальная организация сводится к тому, что существует иерархия распределения различных типов нейронов в виде слоев. Вертикальная организация основана на существовании определенной вертикальной ориентации отростков нейронов различных типов.
Слоистость. На всем протяжении кора состоит из слоев тел клеток. Учитывая количество слоев в различных участках, кору можно разделить на два основных типа: неокортекс и аллокортекс. Неокортекс состоит из 6 слоев нервных клеток, а в аллокортексе их меньше. Специфическим типом аллокортекса является архикортекс, который состоит из трех клеточных слоев. У человека к архикортексу относится гиппокамп. Большая часть коры (90%) относится к неокортексу.
Увеличение площади коры головного мозга в филогенезе происходило путем образования извилин и борозд. Фактически две трети коры мозга человека погружены в толще тканей в пределах этих извилин.
Как вертикальная, так и тангенциальная организация неокортекса отличается особенностями строения составляющих ее нейронов. Морфологических типов нейронов насчитывается более 60. Различают следующие два основных типа клеток — пирамидные и непирамидные (рис. 4.1.6).
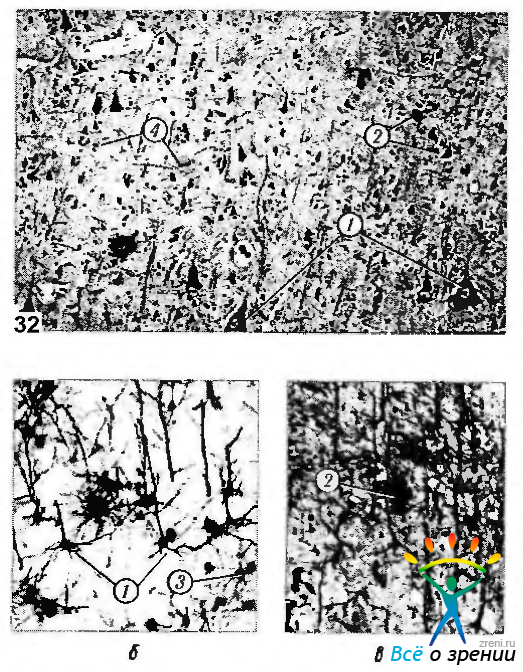
Рис. 4.1.6. Цитологические особенности нейронов коры головного мозга: а — окрашивание по Нисслю; б, в - импрегнация серебром (1 — пирамидные; 2 — корзинкоподобные; 3 — звездчатые; 4— глиальные клетки)
Пирамидные клетки по разным оценкам составляют от 50 до 80% всех нейронов коры. Пирамидные клетки характеризуются треугольной формой тела. От их апикальной поверхности отходит длинный покрытый шипиками дендрит, направляющийся в молекулярный слой коры, где он и ветвится. От базальной и латеральной частей тела нейрона отходит 5—16 коротких дендритов, которые ветвятся в том же слое, где располагается тело нейрона. От середины базальной поверхности тела отходит аксон, направляющийся в белое вещество. На расстоянии 60—90 мкм от тела клетки аксон начинает разветвляться.
Размер пирамидных клеток колеблется от 10 до 50 мкм. Гигантские пирамидные клетки (клетки Беца) имеют размер 100 мкм и обнаруживаются в пятом слое 4-го и 6-го полей коры. Основной функцией пирамидных клеток является интеграция поступающей информации внутри коры и образование эфферентных путей.
Клетки, не относящиеся к пирамидным нейронам, подразделяются на несколько типов в зависимости от их морфологии. Наиболее распространенными являются звездоподобные, корзинчатые, аксо-аксонные клетки, клетки «канделябры», клетки с двойным «букетом» дендритов, веретенообразные, горизонтальные (клетки Кахала) и клетки Мартинотти (рис. 4.1.6, 4.1.7).
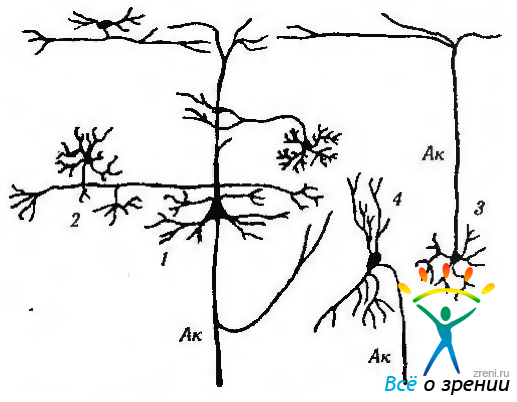
Рис. 4.1.7. Некоторые типы нейронов коры большого мозга: 1 — пирамидный нейрон; 2 — корзинкоподобный нейрон; 3 — звездчатый нейрон: 4 — веретенообразный нейрон. Ак — аксон
Основной функцией непирамидных клеток является интеграция нейронных цепей внутри коры.
Слои неокортекса (рис. 4.1.8).

Рис. 4.1.8. Схематическое изображение пяти основных типов послойной организации коры головного мозга (по Williams et аl., 1995): а—двигательная кора. Отмечается отсутствие гранулярных слоев и наличие скоплений гигантских пирамидных клеток (Беца); б — премоторная кора. Отсутствуют гранулярные слои и гигантские пирамидные клетки. Крупные пирамидные клетки обнаруживаются в V слое; в - сенсорная кора (постцентральное поле). Хорошо выражен гранулярный слой; г — зрительная чувствительная кора. Редукция III слоя и существенное утолщение IV слоя. Последний пересекается зрительной полоской; д— зрительная «психическая» кора. Гранулярный слой хорошо развит, но крупные клетки в V слое отсутствуют
Как указано выше, кора головного мозга обладает послойной организацией нейронов и их отростков. Большинство авторов выделяют шесть слоев:
- Молекулярный (плексиформный) слой (молекулярная пластинка; lamina molecularis (plexiformis)).
Этот слой наиболее поверхностный (расположен под мягкой мозговой оболочкой). Состоит он из плотно переплетенных аксонов и дендритов. Источником волокон являются нейроны коры головного мозга (главным образом, звездчатые клетки), пирамидные клетки (расположены в более глубоких слоях). В этом слое располагаются и терминалы афферентных волокон из других отделов центральной нервной системы. Содержит он сравнительно небольшое количество горизонтальных клеток Кахала с длинными ветвящимися дендритами. Их аксоны участвуют в образовании тангенциального сплетения волокон этого слоя. - Наружный зернистый слой (наружная зернистая пластинка; lamina granularis externa).
Этот слой лежит кнутри от плексиформного слоя. Он называется зернистым из-за большого скопления ядер клеток. Состоит этот слой из тел нейронов, их дендритов и аксонов. Часть клеток пирамидной формы, некоторые звездчатые (мультиполярные). В этом слое аксоны зрительного нерва и дендриты входят в контакт с дендритами нейронов коры. Афферентные волокна формируют многочисленные синапсы с нейронами коры, особенно с апикальными дендритами пирамидных клеток. Дендриты нейронов ветвятся и поднимаются в молекулярный слой, а аксоны либо уходят в белое вещество, либо образуют дуги и также направляются в молекулярный слой. - Слой пирамидных клеток (наружная пирамидная пластинка; lamina pyramidalis externa).
Слой пирамидных клеток варьирует по ширине и состоит из клеток конической формы. Максимально выражен он в ассоциативных и сенсомоторных областях коры. На апикальной поверхности нейронов располагаются дендриты и аксоны. В этом слое обнаруживаются также многочисленные вставочные нейроны (интернейроны) звездчатой формы, нейроны с отростками. ориентированными как в вертикальной плоскости (веретенообразные клетки), так и горизонтальной (корзинкоподобные клетки) плоскости. Их аксоны и дендриты распространяются и на другие слои коры. Слой выполняет преимущественно ассоциативные функции. - Внутренний зернистый слой (внутреняя зернистая пластинка; lamina granularis interna).
Этот слой имеет различную толщину. Наиболее широкий он в области зрительной и слуховой коры мозга. Состоит слой, главным образом, из вставочных нейронов звездчатой формы и небольшого количества пирамидных клеток. В вертикальной плоскости в нем распространяются многочисленные аксоны и дендриты. Выявляются «уплотнения», состоящие из горизонтально распространяющихся отростков, так называемая наружная связка Бейларгера (Baillarger), которая наиболее выделяется в первичной зрительной коре. Слой подразделен на подслои IVA, IVB и IVC (см. «Зрительный путь»), В этом слое заканчивается основная масса таламических афферентных волокон. Аксоны клеток слоя образуют связи с клетками выше- и нижележащих слоев коры. - Слой ганглиозных клеток (внутренняя пирамидная пластинка; lamina pyramidalis interna (ganglionaris)).
Слой ганглиозных клеток также содержит звездчатые и крупные пирамидные клетки (гигантские клетки Беца). Определяются в составе 4-го и 6-го полей (по Бродману). Подобно другим слоям коры в этом слое между клетками распространяются дендриты и аксоны нейронов. Аксоны гигантских и крупных пирамидных клеток проецируются на ядра головного и спинного мозга. Наиболее длинные из них достигают каудальных сегментов спинного мозга. В V слое сосредоточено большинство корковых проекционных эфферентов. - Полиморфный, или веретенообразный, слой (мультиформная пластинка; lamina multiformis).
Последний слой граничит с белым веществом головного мозга. Состоит он из маленьких нейронов, главным образом «зернистых», веретеновидных, звездоподобных вставочных нейронов. Встречаются и маленькие пирамидные клетки. Некоторые из пирамидных клеток (нейроны Мартинотти) отдают длинный двигательный аксон в молекулярный (плексиформный) слой, а вертикально ориентированные его дендриты разветвляются в более глубоких слоях коры. Большинство отростков вставочных нейронов и пирамидных клеток отдают аксоны, которые оставляют кору и направляются к базальным ганглиям, зрительному бугру, гиппокампу, ядрам ствола головного и спинного мозга. В этом слое обнаруживаются различные типы синапсов — аксо-дендритные, аксо-соматические, аксо-аксонные.
Необходимо еще раз подчеркнуть, что типы клеток, их количество и свойственные им связи отличаются в различных участках коры. Это относится и к первичной зрительной коре, о чем более подробно будет изложено в соответствующем разделе.
Классификации неокортекса. Обычное шестислойное строение коры несколько изменяется в различных участках мозга. Эти структурные различия строения по площади коры довольно подробно изучены, и на этой основе созданы цитоархитектонические схемы строения коры, отражающие ее функциональные особенности. Наиболее распространенной классификацией является классификация Бродмана (рис. 4.1.9).
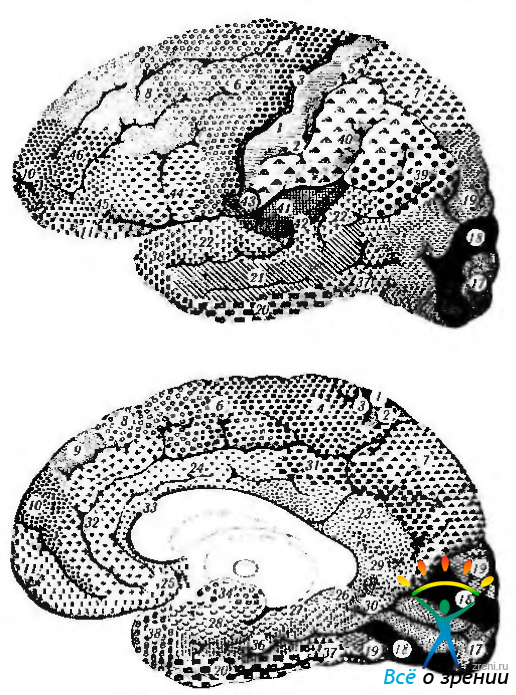
Рис. 4.1.9. Цитоархитектоническая карта коры головного мозга по Бродману: а — наружная поверхность; б—внутренняя поверхность
В соответствии с ней всю поверхность коры можно разделить на 52 поля (области). Эта классификация получила широкое распространение в связи с тем, что структурные различия хорошо коррелируют с функциональными особенностями, о чем будет изложено несколько ниже.
Вертикальная организация коры. Общий поток восходящей и нисходящей информации к коре и от нее организован вертикально. То есть информация, идущая от зрительного бугра, передается звездчатым клеткам IV слоя, которые, в свою очередь, передают ее более высоко и более низко расположенным клеточным слоям для дальнейшей обработки. Подобная вертикальная, или колоночная, организация довольно легко демонстрируется при использовании не только новых морфологических, но и физиологических методов (рис. 4.1.10).
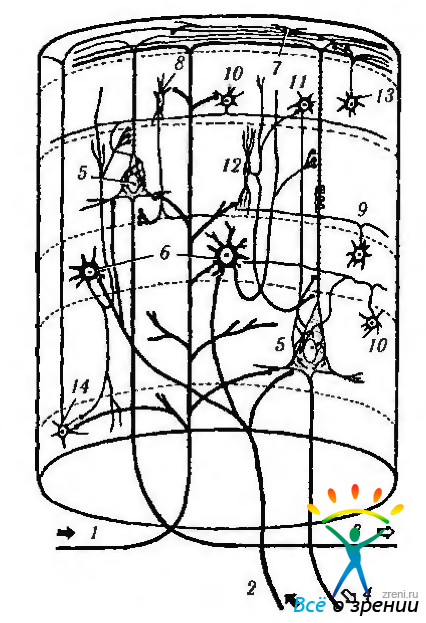
Рис. 4.1.10. Колоночная организация коры полушарий большого мозга (по В. Л. Быкову, 1997). На схеме показаны главные ассоциативные связи клеток основных типов: 1 — кортико-кортикальное афферентное волокно: 2 — таламо-кортикальное афферентное волокно; 3 — ассоциативное эфферентное волокно; 4 — проекционное эфферентное волокно; 5 — пирамидная клетка; б — шипиковая звездчатая клетка; 7 — горизонтальная клетка Кахала; 8 — аксо-аксонная клетка; 9 — клетка -«канделябр», 10—корзинчатая клетка; 11—колонковая корзинчатая клетка; 12 — клетка с двойным букетом дендритов; 13 - клетка с аксональным пучком; 14 — клетка Мартинотти. Римскими цифрами обозначены слои
Эти методы позволили точно установить, что кору мозга можно подразделить на структурно-функциональные колонки, распространяющиеся от мягкой мозговой оболочки до белого вещества. При этом в границах колонки все нейроны выполняют одну и ту же четко определенную функцию. Ширина такой функциональной колонки колеблется от 250 до 500 мкм, а их количество равняется 2—3 млн. Каждая колонка содержит примерно 5000 нейронов. Внутри колонки выделяют также более мелкие мини-колонки, включающие структуры, непосредственно окружающие апикальные дендриты пирамидных клеток. Колонка включает:
- афферентные пути;
- систему локальных связей;
- эфферентные пути.
Афферентные пути организованы следующим образом. В центре колонки проходит примерно 100 возбуждающих кортико-кортикаль-ных волокон — аксонов пирамидных клеток других колонок данного и противоположного полушарий. Они образуют окончания во всех слоях колонки (в том числе на клетках Мартинотти, шипиковых звездчатых клетках, латеральных дендритах пирамидных клеток) и проходят до I слоя, где образуют ветви, уходящие за ее пределы.
Специфические афферентные импульсы по таламо-кортикальным волокнам поступают на тела и дендриты пирамидных клеток и на шипиковые звездчатые клетки IVDcnoH (последние по своим аксонам передают их на апикальные и базальные дендриты пирамидных клеток).
Система локальных связей формируется вставочными нейронами колонки, которые включают более десятка типов клеток. Часть из них обладает тормозной функцией и регулирует преимущественно активность пирамидных клеток. Из тормозных нейронов колонки наибольшее значение имеют следующие (см. рис. 4.1.10):
- Аксо-аксонные клетки, тела которых лежат во II—III слоях, а аксоны идут горизонтально, отдавая многочисленные терминальные веточки, которые образуют тормозные синапсы на начальных сегментах аксонов пирамидных клеток II И III слоев.
- Клетки-«канделябры», которые встречаются во всех внутренних слоях коры. Их аксонные коллатерали идут горизонтально и дают несколько восходящих и нисходящих веточек, которые образуют спиральные ветвления во круг апикальных дендритов пирамидных клеток.
- Корзинчатые клетки, которые располагаются во II слое, на границе III и IV, а также IV и V слоев. Их аксоны проходят горизонталь но на расстояние до 3 мм и, оплетая тела крупных и средних пирамидных клеток, влияют на 20—30 соседних колонок. Колонковые корзинчатые клетки обеспечивают торможение пирамидных клеток по вертикали внутри данной колонки.
- Клетки с двойным букетом дендритов, отходящих вертикально от полюсов клетки, расположены во II—III слоях. Их аксон дает кол латерали, образующие контакты с дендритами как пирамидных клеток, так и непирамидных (в том числе тормозных) нейронов. Первый тип контактов опосредует угнетение пирамидных клеток, а второй — их активацию путем снятия торможения.
- Клетки с аксонным пучком (кисточкой) — звездчатые нейроны II слоя, аксоны которых ветвятся в I слое, образуя связи с дистальными сегментами апикальных дендритов пирамидных клеток и с горизонтальными ветвями кортикокортикальных волокон.
Эфферентные пути формируются следующим образом. Аксоны средних пирамидных клеток III слоя устанавливают связи преимущественно с соседними колонками и колонками противоположного полушария, а аксоны крупных и гигантских пирамидных клеток V слоя, помимо этого, направляются в подкорковые центры, образуя вместе с аксонами веретеновидных клеток VI слоя систему эфферентных волокон коры
Определенные особенности вертикальной организации коры существуют в области зрительной коры. Эти сведения будут приведены в разделе, посвященном зрительной коре.
Приводящие и отводящие связи коры. Нейроны коры можно подразделить на ряд категорий в соответствии с особенностями их связей. К первой категории нейронов относятся проекционные нейроны, которые передают импульсы к подкорковым центрам, таким как зрительный бугор, ствол мозга, спинной мозг или базальные ганглии. Ко второй категории относятся вставочные нейроны, формирующие связи с другими нейронами в том же самом полушарии. К третьей категории относятся комиссурные нейроны, которые напоминают по функции вставочные нейроны за исключением того, что они посылают аксоны в противоположное полушарие через мозолистое тело или переднюю спайку мозга.
Большинство афферентов заканчивается в IV слое коры головного мозга. Эфференты, исходящие из коры, начинаются от нейронов V и VI слоев. Пирамидные нейроны V слоя проецируются непосредственно на ствол мозга или спинной мозг. Нейроны VI слоя являются основным источником прямых проекций на зрительный бугор. Эфференты, направляющиеся к другим областям коры (т. е. вставочные нейроны), исходят от нейронов III и VI слоев.
Архикортекс. Филогенетически наиболее старые структуры коры названы палеокортексом, мезокортексом и архикортексом. Они характеризуются слоистой организацией, но число слоев нейронов в этих участках коры меньше шести. Основной областью палеокортекса является обонятельная кора, мезокортекса — кора поясной извилины, а основной частью архикортекса — гиппокамп. Гиппокамп отличается от коры обратным расположением слоев. При этом серое вещество гиппокампа располагается внутри, а белое — снаружи. Часть гиппокампа состоит только из трех слоев клеток.
Нейромедиаторная система коры. В коре выявлено более десятка различных нейромедиаторов и рецепторов к ним. Это норадреналин, серотонин, ацетилхолин, вазоинтерстициальный кишечный полипептид (VIP), соматостатин, холецистокинин, глютамат и др.
Продолжение в следующей статье: Анатомия головного мозга ? Часть 2
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0