Движения глаз │ Часть 2


Описание
Анатомический субстрат плавного слежения. Надъядерный контроль плавного слежения пока точно не выяснен. На рис. 4.4.17— 4.4.19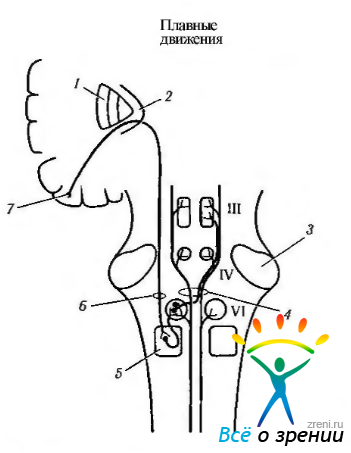
Рис. 4.4.17. Надъядерный контроль плавного слежения в горизонтальном плоскости в левую сторону (цель появляется в контрлатеральном поле) (по Reek et al., 1981): 1—базальные ганглии; 2— внутренняя капсула; 3 — ножка мозжечка; 4 — медиальный продольный пучок; 5 — парамедианная ретикулярная формация варольева моста; 6—кортикобульбарный тракт; 7 — ассоциативные поля затылочной коры. Основной путь обозначен красным цветом, остальные — черным[/center]
[center]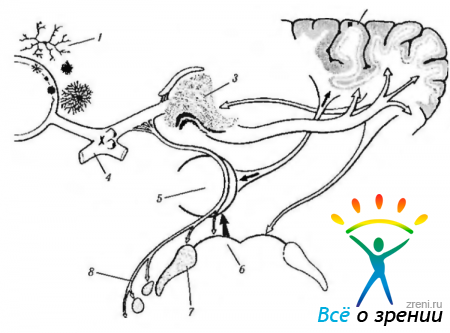
[center]
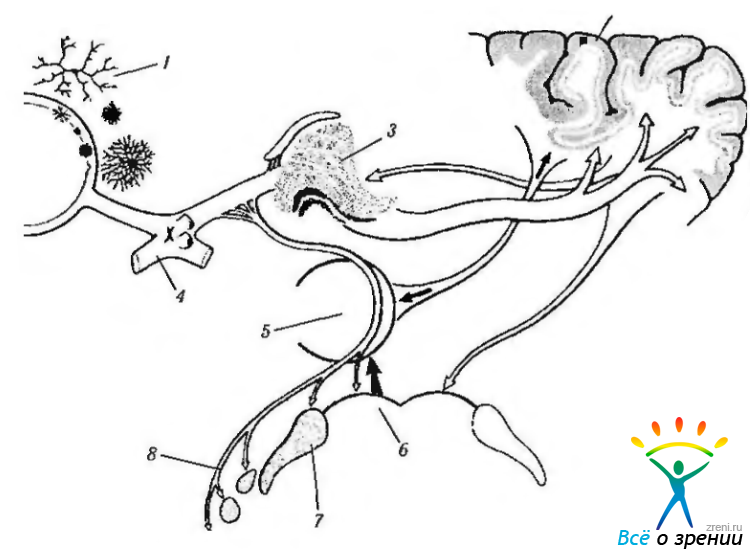
Рис. 4.4.18. Схема связей зрительного тракта с верхними бугорками четверохолмия, подушкой зрительного бугра и «добавочной зрительной системой»: 1 — нейроны сетчатки; 2 — кора; 3 — наружное коленчатое тело; 4 — зрительный перекрест; 5 — подушка зрительного бугра; б — верхние бугорки четверохолмия; 7— претектальная область; 8— добавочная зрительная система

Рис. 4.4.19. Связи мозжечка, определяющие движения глаз (по Bron et al., 1997): а — со стволовой частью мозга (1 — нижняя мозжечковая ножка; 2 — ретикулярное ядро крыши моста; 3— парамедианная ретикулярная формация моста; 4— верхняя мозжечковая ножка; 5 — полушарие; 6— червь; II, III, VI — ядра черепно-мозговых нервов); б — с вестибулярными ядрами (1—добавочный тракт Дейтерса; 2— соединительное плечо; 3— медиальный продольный пучок; 4 — узелок мозжечка; 5 — necl. prepositus hypoglossi; 6 — nucl. perihypoglossi; 7 — вестибулярные ядра; 8 — первичные вестибулярные афференты; 9 — червь; 10 — язычок червя)
приведены лишь упрощенные схемы. От контроля саккад контроль плавного слежения отличается тем, что проекция от коры головного мозга на парамедианную ретикулярную формацию осуществляется с этой же стороны.
Предполагают, что сигнал, индуцированный зрительным стимулом, возникает в затылочной коре головного мозга. Нейроны коры этой области проецируются на среднее височное зрительное поле. В свою очередь, нейроны среднего височного зрительного поля проецируются на медиальное верхнее височное зрительное поле, оказывающее помощь в слежении за зрительной целью в тех случаях, когда она временно исчезает из поля зрения. Обе височные проекции, в свою очередь, проецируются на затылочную кору. В плавном слежении участвуют также теменная и лобная зрительная кора.
Нисходящий путь от коры головного мозга направляется с этой же стороны к ретикулярной формации моста, латеральным ядрам моста и латеральным ядрам добавочной зрительной системы (рис. 4.4.17—4.4.19). Добавочная зрительная система, в свою очередь, проецируется на мозжечок, который затем проецируется на медиальные вестибулярные ядра. Конечной точкой проекции являются ядра глазодвигательных нервов. При этом волокна дважды перекрещиваются (в среднем мозге — перекрест Мейнерта (Meynert) и в области моста).
Добавочная зрительная система состоит из скопления нейронов, расположенных вне первичной зрительной коры и получающих проекции от сетчатки (рис. 4.4.18). К ней относятся претектальные ядра, в состав которых входит ядро зрительного тракта, ядра крыши моста, медиальное ядро моста, л. perihypoglossi, вестибулярная комиссура.
Основной функцией добавочной зрительной системы является интеграция получаемой зрительной информации, а также информации о положении тела и головы в пространстве, получаемой от вестибулярной, соматосенсорной и зрительной систем. Благодаря этой интеграции получаемой информации и формируются сигналы, идущие к исполнительным нервным центрам, участвующим в движении глаза.
В контроле горизонтальных саккадических движений, как указано выше, принимают участие ретикулярная формация среднего мозга и ряд структур мозжечка. Ретикулярная формация среднего мозга (formatio reticularis) содержит нейроны, «кодирующие» положение глаза. Некоторые из них активны в течение ипсилатерального плавного слежения или в течение медленной стадии вестибулярного рефлекса. Другие нейроны, находящиеся ниже ядра отводящего нерва, формируют премоторную команду, определяющую соответствие скорости движения глаз скорости слежения за целью.
Мозжечок является ключевой структурой в функционировании плавного слежения. Структурой, соединяющей кору головного мозга, глазодвигательную часть мозжечка и ствол мозга, является дорзолатеральное ядро ствола мозга. Это ядро содержит нейроны, кодирующие направление и скорость движения цели и глаза и координируют их. Проецируется дорзомедиальное ядро ствола мозга, в первую очередь, на мозжечок.
Клочок мозжечка (flocculus) получает зрительную информацию посредством мшистых волокон. Клетки Пуркине (определяющие скорость слежения) клочка суммируют сигналы, поступающие от вестибулярной и зрительной системы. При этом они формируют сигналы, определяющие скорость движения глаза за целью. Клетки Пуркине проецируются на n. perihypoglossi или парамедианную ретикулярную формацию моста, т. е. структуры, важные в заключительном синтезе команды, определяющей направление взгляда (рис. 4.4.19).
Зубчатое ядро (nucleus dentatus) и лежащая вблизи Y-образная группа вестибулярных ядер (вероятно, получающая проекции от клочка) вовлечены в контроль вертикального плавного слежения и отмены вестибуло-глазного рефлекса.
Система плавного слежения нередко повреждается, причем раньше, чем саккады. Наступает это при двухстороннем поражении базальных ганглиев и ствола мозга. Подобные состояния характерны для алкогольного и барбитуратного отравления. При этом скорость плавного слежения снижается и больной начинает использовать саккады для контроля цели даже при ее небольшой скорости движения.
Система, контролирующая фиксацию глаз
Фиксация взгляда на неподвижную цель длится, по крайней мере, пять секунд. При этом глазное яблоко, тем не менее, находится в движении. Эти движения глаза включают быстрые движения (микросаккады) и медленные клонические движения.
Микросаккады никогда не прекращаются. Их амплитуда всего несколько угловых секунд, а частота находится в переделах 20—150 Гц.
В результате медленного клонического «дрейфа» точка фиксации уходит от фиксируемого объекта. С помощью микросаккад происходит компенсация этих дрейфов и восстановление правильной фиксации объекта.
Вергентные движения. Вергентные движения относятся к так называемым версиям. Отличие сводится к тому, что при версионных движениях зрительные оси движущихся глаз остаются параллельными, а при вергентных угол между ними изменяется.
Способностью вергентных движений обладает только небольшое число видов позвоночных и приматы. Функцией вергентных движений является достижение проекции изображения на область центральной ямки каждого глаза. Это достигается управлением изменения угла между зрительными осями двух глаз.
Различают три источника стимула начала вергентных движений. Первый стимул — это так называемое «бинокулярное неравенство». Понятие «бинокулярное неравенство» характеризует состояние, при котором изображение проецируется на сетчатку двух глаз, но на различные ее участки. Вторым стимулом является нерезкость (стертость) изображения, связанная с отсутствием адекватной аккомодации. По всей видимости, состояние хрусталиков обоих глаз контролируется высшими центрами нервной системы и используется для вычисления расстояния до зрительной цели. Третий стимул, относящийся к так называемым «тоническим», учитывает состояние системы при отсутствии конвергенции, что бывает, например, в условиях полной темноты. У человека в подобных условиях угол конвергенции равен 3°. Последний тип информации, используемый в вергентных движениях, — это информация о монокулярной линейной глубине (линейная перспектива), определяющая расстояние до цели в монокулярных условиях.
Вергентные движения глаз относятся к медленным движениям. Система вступает в действие при смещении точки фиксации в пространстве. При этом направление движения одного глаза является зеркальным отображением движения другого. Если фиксируемая точка на значительном расстоянии от наблюдателя, то зрительные оси параллельны. Если взор смещается на более близкий предмет, зрительные оси конвергируют. При необходимости перевода взора на отдаленный предмет оси разводятся (дивергенция). В обычных условиях акту конвергенции осей глаза сопутствует сужение зрачка и аккомодация. Эти три компонента формируют так называемый «конвергентно-аккомодационно-зрачковый рефлекс» (аккомодационный рефлекс).
Вергентная система начинает работать после саккадического движения, спустя римерно 160 мс. Скорость движения глаз при этом равняется 20° в секунду. Конвергенция или дивергенция зрительных осей происходит со скоростью 10° в секунду. Это медленное движение может длиться почти секунду.
Анатомический субстрат вергентных движений. Надъядерный контроль вергентной системы изучен далеко не достаточно.
Известно, что высшие зрительные центры коры головного мозга особо важны в восприятии неравенства изображений на сетчатках и инициирования вергентных движений глаза. Об этом свидетельствует то, что одновременная стимуляция зрительных полей лобной и затылочной коры приводит к конвергенции (стимуляция полей 19 и 22 коры затылочной доли). На диспарантность сетчаток реагируют также поля коры средней височной и затылочной областей. Промежуточная премоторная инициация вергенции определяется также стимуляцией нейронов, расположенных вблизи ядер глазодвигательного нерва, которые проецируются на ядро внутренней прямой мышцы глаза и ядро отводящего нерва.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Выявлены еще другие участки мозга, участвующие в индукции вергентных движений. Это — средний мозг, мост. Установлено, что участок среднего мозга, лежащий латеральней глазодвигательного ядра, контролирует угол конвергенции. К вергентным движениям приводит также стимуляция моста между медиальными продольными пучками. В вергентных движениях участвует и мозжечок, о чем свидетельствует тот факт, что его ампутация приводит к нарушению движения глаз.
В последние годы появились данные о том, что «центром вергентных движений» являются скопления двух типов нейронов ретикулярной формации моста. По своим функциям он напоминает «центр саккад». Первый тип нейронов, «пакетные» нейроны, формирует импульсы перед началом вергентного движения. При этом частота импульсов связана со скоростью движения, а их количество — с амплитудой. Вовлечены при этом «пакетные» нейроны, обеспечивающие как схождение, так и расхождение зрительных осей. «Тонические вергентные нейроны» объединяют динамические и статические реакции вергентной системы. Необходимо отметить и то, что вергентные движения интимно связаны с саккадами. Эта связь проявляется рядом этапов совместных действий.
Рефлекторные системы, не связанные со зрительной системой (вестибулярная). Рефлекторные изменения движения глаз происходят всякий раз, когда изменяется положение головы и шеи. При этом возбуждаются рецепторы полукружных каналов или макулярных органов. Эти рефлекторные движения глаз осуществляются за счет прямых связей между нейронами вестибулярных ядер, мотонейронами глазных мышц и нейронами парамедианной ретикулярной формации моста и ретикулярной формации среднего мозга.
Сигналы от рецепторов шеи, вестибулярного аппарата и от наружных мышц глаза определяют положение головы относительно тела и в пространстве, а также положение глазных яблок. Таким образом, формируется «эфферентная копия», обходящая зрительный путь.
Вестибулярная система может компенсировать значительно более существенные движения изображения на сетчатке, чем система плавного слежения. Поворот туловища является стимулом компенсационных движений глаза, контролируемых вестибулярной системой. Скорость движений глаза, индуцированных вестибулярной системой, достигает до 300° в секунду. Движение головы более 10—15° приводит к саккадическому движению глаз в противоположном направлении.
Несмотря на то, что вестибулярная система не контролируется зрительной системой, все же между ними существует взаимодействие, называемое вестибуло-глазным рефлексом.
Движения глаза в вестибуло-глазной системе содружественные и состоят из двух стадий. Первая стадия — это плавное следящее движение. Прерывается оно саккадами, которые повторно приводят глаза к средней позиции в глазнице. Этот тип повторяющихся медленных и быстрых движений глаза, относится к одному из типов нистагма (вестибулярный нистагм).
Необходимо помнить то, что тип нистагма определяется направлением быстрой стадии. Так, например, левосторонний нистагм характеризуется быстрым движением влево. Только медленная стадия нистагма компенсирует движения туловища.
Нистагм может быть индуцирован обычными физиологическими стимулами или являться выражением различных заболеваний, нарушающих нормальный контроль движений глаза.
Анатомический субстрат вестибулярного и оптокинетического рефлексов. Основной функцией вестибулярной системы и мозжечка является определение направления взора при любом положении и движении тела.
Перед тем как остановиться на рефлексах, нам представляется необходимым более подробно описать строение вестибулярного аппарата и мозжечка, т. е. структур, во многом определяющих возможность этой функции.
Первоначально необходимо охарактеризовать саму вестибулярную рефлекторную дугу.
Строение вестибулярного аппарата. В костном лабиринте височной кости лежат три (с каждой стороны) мембранных полукружных канала (canalis semicircularis anterior, posterior, lateralis), выполненных эндолимфой. Как показано на приведенных рисунках (рис. 4.4.21—4.4.23),
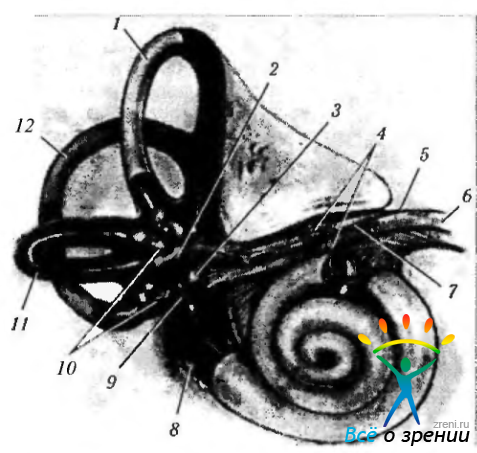
Рис. 4.4.21. Органы и нервы внутреннего уха (по Walsh. 1969): 1 — верхний полукружный канал; 2 - маточка; 3 — мешочек; 4 — вестибулярный ганглий; 5 — внутренний слуховой проход; б — лицевой нерв; 7—вестибулярная ветвь VIII нерва; 8—круглое окно; 9 — овальное окно; 10 — ампульные нервы; 11 — горизонтальный полукружный канал; 12— задний полукружный канал. Вид спереди лабиринта и раковины (кохлеа). Обращает на себя внимание наличие тесных анатомических отношений между вестибулярной и кохлеарной ветвями пред-дверно-улиткового (VIII) и лицевого (VII) нервов во внутреннем слуховом канале. Также видны три ампулярных нерва (терминальные концы нервов на иллюстрации обрезаны), которые собирают информацию от каждого полукружного канала
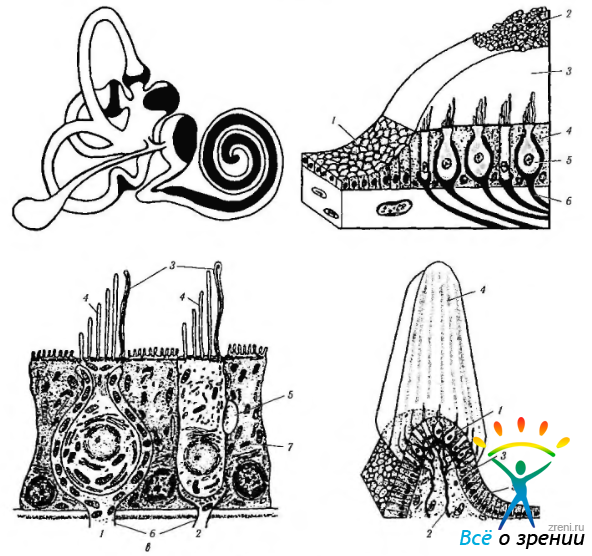
Рис. 4.4.22. Строение вестибулярного аппарата: а — распределение сенсорного эпителия в перепончатом лабиринте и улитке (темные области); б — схема строения вестибулярного аппарата млекопитающих (1 — эпителий перепончатого канала; 2 — отолиты; 3— отолитовая мембрана; 4 — поддерживающие клетки; 5 — волосковые клетки; б — нервные окончания); в — ультраструктура волосковых клеток органа равновесия (1 — волосковые клетки первого типа; 2— волосковые клетки второго типа; 3— стереоцилии; 4 — киноцилии; 5 — афферентные нервные окончания; 6 — эфферентные нервные окончания; 7 — поддерживающие клетки); г — ампулярный гребешок (1 — волосковые клетки; 2 — нервные волокна; 3 — поддерживающие клетки; 4 — купол: 5 — эпителий перепончатого канала)
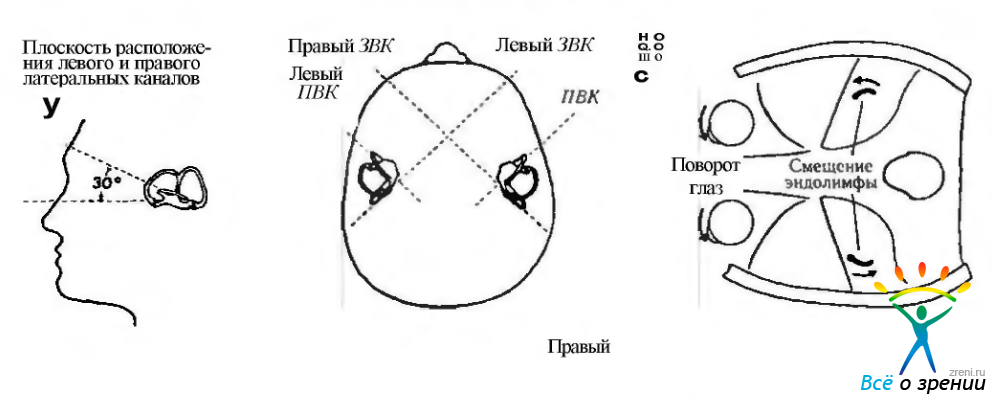
Рис. 4.4.23. Плоскости расположения полукружных каналов с правой и левой стороны (а) (по Barber, Stockwell, 1976) и направление смещения эндолимфы при повороте головы в правую стороны (б) (по Cogan, 1966). Глаза при этом поворачиваются влево: ПВК передний вертикальный канал; ЗВК — задний вертикальный канал
существует один горизонтальный и два вертикальных канала, которые лежат в трех плоскостях, перпендикулярных друг другу. Горизонтальные (латеральные) каналы с двух сторон лежат в одной плоскости, в то время как плоскость переднего канала одной стороны параллельна плоскости заднего канала противоположной стороны. Горизонтальные каналы соединяются с одного конца с маточкой, которая, по сути, является большим расширением перепончатой части лабиринта. Вертикальные каналы (передний и задний) соединяются с маточкой своим одним концом, а другим концом соединяются друг с другом. Расширения одного конца полукружных каналов называются ампулой (ampula ossea anterior, posterior, lateralis). Каждая ампула содержит «волосковые» чувствительные клетки, погруженные в желатиноподобный материал, называемый капсулой (рис. 4.4.22).
Таким образом, вестибулярный орган состоит из двух морфологических субъединиц, а именно:
- статолитового аппарата (macula utriculi и macula sacculi)
- и полукружных каналов (передний и задний вертикальные и горизонтальный каналы).
В области макул и в полукружных каналах вблизи от так называемых ампул расположен содержащий рецепторы сенсорный эпителий, покрытый желеобразной массой. В макулах эта масса покрывает сенсорные клетки и содержит соли кальция в виде кристаллов кальцита (отолитовая мембрана). В полукружных каналах желеобразная масса кристаллов не содержит.
В вестибулярном органе имеется два типа клеток («волосковые» клетки), на апикальной поверхности которых располагаются реснички. Различают два типа ресничек — стереоцилии (по 60—80 ресничек в одной клетке) и киноцилии (по одной на каждой клетке) (рис. 4.4.22).
Рецепторы относятся к вторичным сенсорным клеткам. Как указано выше, они не обладают собственными отростками, а иннервируются афферентными волокнами нейронов вестибулярного ганглия, входящими в состав вестибулярного нерва (рис. 4.4.21). На рецепторных клетках также оканчиваются эфферентные нервные волокна.
Афферентные волокна передают информацию об уровне возбуждения периферического органа в центральную нервную систему, а эфферентные волокна изменяют чувствительность рецепторов.
Как было показано выше, реснички рецепторных клеток входят в отолитовую мембрану. Поскольку она содержит соли кальция, ее удельный вес больше, чем эндолимфы, заполняющей полость мешочка (sacculus) и маточки (utricuius). По этой причине при действии на организм линейного ускорения отолитовый аппарат скользит по сенсорному эпителию. В результате этого реснички отклоняются и рецептор получает стимул.
Когда тело выпрямлено, а голова находится в нормальном положении, макула маточки оказывается под углом и тяжелая отолитовая мембрана соскальзывает по сенсорному эпителию. При этом реснички изгибаются и происходит стимуляция рецептора. В зависимости от направления наклона частота нервной импульсации в афферентных волокнах увеличивается или уменьшается.
Стимуляция макулы мешочка происходит так же, но эта структура расположена почти вертикально при нормальном положении головы. Таким образом, при любом положении головы каждая из отолитовых мембран занимает определенное положение относительно эпителия. При этом создается определенная картина возбуждения, отражающая расположение головы и тела в пространстве, которая затем обрабатывается в центральном отделе вестибулярной системы.
Второй вид адекватных стимулов для ресничек вестибулярных рецепторов воспринимается в полукружных каналах. Капсула, расположенная в полукружных каналах, обладает такой же плотностью, что и эндолимфа. По этой причине линейное ускорение не влияет на этот орган. Возбуждение происходит при угловом ускорении (вращении). При повороте головы полукружные каналы поворачиваются вместе с ней. Однако эндолимфа в силу своей инерции в первый момент остается неподвижной. Поэтому капсула отклоняется в направлении, противоположном направлению вращения. Это отклонение сдвигает реснички и таким образом изменяет активность в афферентном нерве. В горизонтальных каналах все рецепторы ориентированы так, что киноцилии обращены к маточке, поэтому активность в афферентных волокнах увеличивается, когда капулы отклоняются в сторону маточки. В левом горизонтальном полукружном канале это происходит при вращении влево.
В вертикальных каналах активация нервных волокон происходит при утрикофугальном (в направлении, противоположном утрикулусу) отклонении капсулы.
По картине активности нервных волокон, приходящих от трех полукружных каналов с каждой стороны, мозг получает информацию относительно углового ускорения головы.
Очевидно, что система наиболее чувствительна к вращениям, плоскость которых совпадает с плоскостью канала. Для «вычисления» скорости вращения головы в трех измерениях используются три отдельных канала, ориентированных в различных плоскостях. Фактически, имеется шесть каналов, по три с каждой стороны. Вращение головы в левую сторону активизирует два канала, расположенных в горизонтальной плоскости. «Волосковые» клетки в одном из этих каналов будут деполяризоваться, поскольку капула отклоняет их киноцилии. «Волосковые» клетки другого канала, наоборот, гиперполяризуются, так как капула отклоняется в противоположном направлении, отклоняя при этом киноцилии в противоположную сторону относительно первого канала. Регистрация сигнала только от одного канала выявила, что при этом точной информации о скорости вращения один канал не дает.
Вестибулярные нервы. Нервные волокна от макул достигают преддверного ганглия (g. vestibulare, Scarpa), расположенного в наружной части внутреннего слухового прохода. Преддверно-улитковый нерв (n. vestibulocochlearis), сформированный центростремительными волокнами, проходит поперек лицевого нерва снизу и позади нерва улитки (n. cochlea) и поступает в ствол мозга. Эти первичные вестибулярные аксоны проецируются, главным образом, на вестибулярные ядра (nuclei vestibulares) и мозжечок (рис. 4.4.19).
Вестибулярные ядра (nuclei vestibulares). Первичные афферентные волокна вестибулярного нерва оканчиваются в области вестибулярных ядер в продолговатом мозге под дном четвертого желудочка. С каждой стороны имеется по четыре ядра, отличающихся как структурно, так и функционально. Это верхнее ядро (ядро Бехтерева), медиальное (ядро Швальбе), латеральное (ядро Дейтерса) и нижнее (ядро Роллера) (рис. 4.4.19, 4.4.24, 4.4.25).
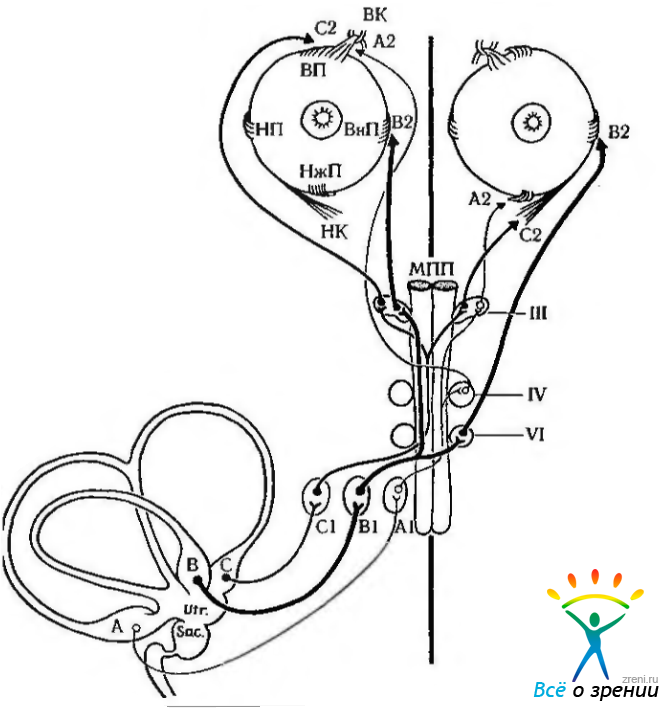
Рис. 4.4.24. Схема связи различных полукружных каналов с соответствующими парами наружных мышц глазного яблока (по Шабе, Форд, 1976): от каждого полукружного канала (А, В, С) нервные волокна идут к нейронам комплекса вестибулярных ядер, расположенных в латеральных отделах дна четвертого желудочка (A1, B1, С1). Волокна нейронов вестибулярных ядер идут в восходящем направлении в составе медиального продольного пучка (МПП) к ядрам черепно-мозговых нервов, иннервирующих мышцы глазного яблока: глазодвигательному (III), блоковому (IV) и отводящему (VI). .Аксоны глазодвигательных нервов направляются к соответствующим парам наружных мышц глаза (А2, В2, С2). Раздражение горизонтального полукружного канала (В) вызывает появление импульсов, поступающих в группу нейронов В1 и далее по медиальному продольному пучку к ядрам отводящего и глазодвигательного нервов. Эти черепно-мозговые нервы идут к внутренней прямой (ВнП) и наружной прямой (НП) мышцам глазного яблока. Подобные связи обеспечивают формирование рефлекторной реакции нижней прямой (НжП), верхней прямой (ВП) и нижней, косой (НК) мышц при раздражении переднего и заднего полукружных каналов

Рис. 4.4.25. Схема расположения ядер глазодвигательных нервов (а) и путей, активируемых во время стимуляции движения глаз вестибулярным аппаратом (полукружные каналы): а - расположение ядер глазодвигательных нервов; б - декстро-элевация; возбуждение правого переднего канала (торможение парного левого заднего канала) стимулирует движение глаза вверх и направо. Отмечается наличие двойного перекреста волокон при возбуждении ипсилатеральной верхней прямой мышцы (путь от верхнего вестибулярного ядра через верхнее соединительное плечо к контрлатеральному ядру третьего нерва; активация мотонейронов верхней прямой мышцы приводит к стимуляции ипсилатеральной верхней прямой мышцы). Отмечается также, что отток эндолимфы в направлении от ампулы является возбуждающим в вертикальном канале, а отток в направлении ампулы возбуждающим в горизонтальном канале; в поворот глаза влево и вниз (леводепрессия). Возбуждение правого заднего канала (торможение левого переднего канала) стимулирует движение глаз налево и вниз; г — декстрадепрессия. Торможение правого переднего канала (возбуждение левого переднего канала) стимулирует начало движения глаз вверх и налево; д — левовесия. Поворот головы направо возбуждает правый горизонтальный канал (тормозит левый горизонтальный канал) и стимулирует взгляд налево; е — иллюстрируется движение налево и нистагм с быстрой фазой направо, т. е. правый нистагм
Афферентные импульсы от вестибулярных рецепторов, приходящие в вестибулярные ядра, сами по себе не обеспечивают точной информации о положении тела в пространстве. Это связано с тем, что угол поворота головы, подвижной в шейном сочленении, не зависит от положения туловища. Центральная нервная система должна получать информацию о положении головы относительно туловища и учитывать ее при определении положения тела в целом. Следовательно, вестибулярные ядра получают дополнительную афферентную информацию от шейных рецепторов (от мышц и сочленений шеи). Чувствительные афференты, идущие от других суставов, также направляются в вестибулярные ядра. Обеспечивается эта информация при помощи преддверно-спинномозгового пути (tractus vestibulospinalis). Происходит это следующим образом. Повышение активности правого вестибулярного ядра может активизировать мышцы туловища посредством правого латерального преддверно-спинномозгового пути. Клетки, дающие начало латеральному преддверно-спинномозговому пути, лежат в латеральном вестибулярном ядре (nucleus vestibularis lateralis). Аксоны этих клеток спускаются через каудальную часть ствола мозга и поступают в спинной мозг в составе вентрального канатика и иннервируют нейроны во всю длину спинного мозга. Эти проекции не перекрещиваются. Благодаря этому пути вестибулярный аппарат управляет мышцами туловища, восстанавливая его положение.
Повышение активности правого вестибулярного ядра может также активизировать мышцы тела и через второй, меньший, нисходящий путь спинного мозга. Называется он медиальным преддверно-спинномозговым путем (tractus vestibulospinalis medialis). Клетки медиального вестибулярного ядра (nucleus vestibularis medialis) дают аксоны, которые спускаются с двух сторон, занимая положение непосредственно вблизи срединной линии с дорзальной поверхности моста и продолговатого мозга. Эти нисходящие аксоны распространяются каудально и входят в спинной мозг, где лежат в медиальной части вентрального канатика. Этот путь соединяется с двигательными нейронами шеи, играющими роль в установлении нормального положения головы, иннервируя мышцы шеи.
Нервные волокна, выходящие из вестибулярных ядер, образуют связи и с другими отделами центральной нервной системы.
К ним относятся:
- Пути, обеспечивающие прямую связь вестибулярных ядер с ядрами глазодвигательного нерва, опосредующие регуляцию скорости движения глаз, вызываемой вестибулярной активностью (рис. 4.4.24).
- Пути, направляющиеся в вестибулярные ядра противоположной стороны мозга.
- Связи с мозжечком, в особенности с архицеребеллумом (рис. 4.4.19).
- Связи с ретикулярной формацией.
- Пути, проходящие через зрительный бугор—в—постцснтральную—извилину головного мозга, обеспечивающие сознательную обработку вестибулярной информации и сознательную ориентацию в пространстве.
- Волокна, направляющиеся в гипоталамус.
Приведенное множество связей дает возможность вестибулярной системе играть центральную роль в формировании моторных эфферентов. обеспечивающих поддержание позы и соответствующие глазодвигательные реакции.
Функции вестибулярной системы. Теперь мы подробней остановимся на функции вестибулярной системы и ее анатомическом обеспечении.
Движения глаза индуцируются вестибулярным аппаратом компенсаторно. Этот рефлекс сохраняет зрительную цель на центральной ямке сетчатой оболочки при изменении положения головы. Например, быстрый поворот головы вправо приводит к компенсаторному рефлекторному повороту обоих глаз влево.
Этот рефлекс называется вестибуло-глазным и состоит из трехнейронной дуги (вестибулярный нерв, вестибулярные и глазодвигательные ядра) и параллельного полисинаптического добавочного зрительного пути (ретикулярная формация, п. perihypoglossi) и промежуточного (интерстициального) ядра (рис. 4.4.19, 4.4.24—4.4.26).
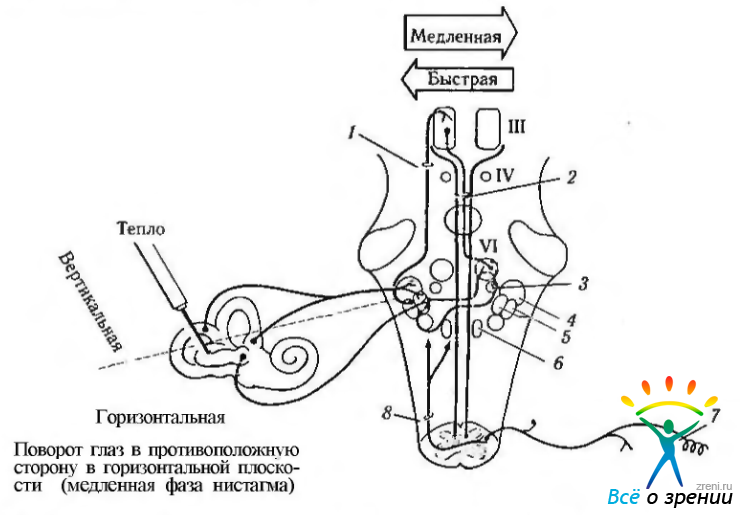
Рис. 4.4.26. Надъядерный контроль вестибулярной и проприорецептивной рефлекторных систем: 1 — восходящий тракт Дейтерса (эксцитатный); 2 - медиальный продольный пучок (эксцитатный); 3 — парамедианная ретикулярная формация моста (эксцитатная); 4 — верхнее вестибулярное ядро; 5 — медиальное и латеральное вестибулярные ядра; 6 - n. prepositus hypoglossi; 7 — рецептор мышцы; 8 спинно-вестибулярный тракт. Эксцитатный тракт выделен красным цветом. Нервные волокна, исходящие из горизонтального полукружного канала, направляются к латеральному вестибулярному ядру. Во время медленной фазы нистагма интернейроны вестибулярного ядра возбуждают контрлатеральную парамедианную ретикулярную формацию моста. В свою очередь, возбуждаются контрлатеральные нейроны отводящего нерва и субъядро ипсилатеральной внутренней прямой мышцы, благодаря медиальному продольному пучку. Во время быстрой фазы нистагма часть ингибирующих импульсов идет к ипсилатеальному субъядру внутренней прямой мышцы через восходящий тракт Дейтерса, который располагается латеральней медиального продольного пучка Контрлатеральные проприорецептивные рецепторы и восходящие спинно-вестибулярные волокна также оканчиваются вблизи вестибулярных ядер и n. prepositus hypoglossi
Происходит это следующим образом (рис. 4.4.24, 4.4.25, табл. 4.4.6).

Таблица 4.4.6. Особенности вестибуло-глазных проекций (по Precht, 1977)
Поворот головы вправо сопровождается смещением волосковых клеток правого горизонтального полукружного канала вправо, что приводит к возбуждению правого вестибулярного нерва и нейронов правого вестибулярного ядра. Клетки правого вестибулярного ядра отдают аксоны к контрлатеральной парамедианной ретикулярной формации моста, лежащей в медиальной части покрышки моста вентральней ядра отводящего нерва. Ретикулярная формация является интегративной областью, участвующей в генерации горизонтальных движений глаз (см. выше). Нейроны левой ретикулярной формации проецируются на левое ядро отводящего нерва (nucleus nervi ctbducens). Как описано выше, ядро содержит два типа нейронов. Первый тип (крупные нейроны) отдают отростки, формирующие отводящий нерв, иннервирующий наружную прямую мышцу глаза. Второй тип нейронов (мелкие) отдает аксоны, которые не покидают мозг. Эти волокна перекрещиваются или не перекрещиваются и в составе медиального продольного пучка (fasciculus longitudinalis medialis; тракт Дейтерса) оканчиваются на ядрах глазодвигательного нерва. Перекрещенные волокна иннервируют внутреннюю прямую мышцу глаза. Таким образом, при повороте головы вправо происходит возбуждение правого преддверного нерва (n. vestibularis), возбуждение правого вестибулярного ядра, возбуждение нейронов левой парамедианной ретикулярной формации моста, повышение активности крупных и мелких нейронов левого ядра отводящего нерва и рефлекторный поворот левого глаза налево (посредством левой наружной прямой мышцы глаза) и правого глаза также влево (через медиальный продольный пучок вход к правой внутренней прямой мышце; III нерв).
Как указывалось выше, описанный рефлекс называется вестибуло-глазным рефлексом. Этот феномен, вызываемой вестибулярной стимуляцией. представляет собой движения глаз, противоположные вращению, так что поддерживается постоянное направление взора. Прежде чем глаза достигают положения крайнего отклонения, они резко перемещаются в направлении вращения, так что в поле зрения оказывается другой участок пространства. Эта быстрая фаза затем сменяется медленным движением в противоположном направлении. При вращении тела вокруг вертикальной оси раздражаются только полукружные каналы. Поэтому отклонение капул двух горизонтальных полукружных каналов вызывает горизонтальный нистагм.
Аналогичную реакцию можно получить и при стимуляции полукружных каналов различными физическими факторами, например нагреванием или охлаждением наружного слухового прохода водой, что широко используется в дифференциальной диагностике различных типов нистагма. Так, стимуляция горизонтального полукружного канала приводит к возникновению нистагма, который состоит из контрлатеральной медленной стадии и ипсилатеральной быстрой.
Для вертикальных движений глаза вестибулярные нейроны проецируются через медиальный продольный пучок (и, возможно, соединительное плечо). Но эксцитатный (возбуждающий) путь, идущий от переднего канала до ипсилатеральнои верхней прямой мышцы и контрлатеральных нижних косых мышц, проходит в верхнюю мозжечковую ножку (соединительное плечо).
При стимуляции передних и задних полукружных каналов вестибулярная система формирует «медленный» компонент вертикального нистагма. Особенностью является то, что в вертикальных движениях участвуют верхние и медиальные вестибулярные ядра. При этом стимулируются как эксцитатные, так и ингибирующие пути.
Необходимо обратить внимание на то, что произвольный взгляд и вестибулярные механизмы конкурируют в парамедианной ретикулярной формации моста. При этом произвольный взгляд ингибирует вестибулярный нистагм.
Вестибуло-глазной рефлекс обладает большой пластичностью. В этом процессе большую роль играет мозжечок, изменяющий силу обратной связи. При этом мозжечок использует зрительную информацию, получаемую от сетчатки во время движения туловища, с целью определения «силы обратной связи». Именно благодаря этой информации мозжечок формирует «сигнал ошибки», который и используется для увеличения или уменьшения «силы обратной связи».
Оптокинетическая система. Когда испытуемый фиксирует равномерно движущийся зрительный стимул, происходит периодическое чередование саккад и медленных следящих движений глаз. Это и есть оптокинетический нистагм. Развивается он при стимуляции периферии сетчатки.
Подобно вестибуло-глазному рефлексу, оптокинетическая система активирует мышцы глаза для стабилизации взора в течение вращения головы. Однако она отличается тем, что использует зрительную информацию. Как и вестибуло-глазной рефлекс, оптокинетический нистагм обладает «силой обратной связи», которую можно измерить. Этот показатель представляет собой отношение скорости вращения глаза к скорости вращения изображения.
В процессе формирования оптокинетического нистагма зрительные афференты проецируются на вестибулярные ядра несколькими путями. Это облегчает интеграцию вестибуло-глазного и оптокинетического рефлексов. Схема, иллюстрирующая возможные пути оптокинетической системы, представлена на рис. 4.4.26.
вращение туловища стимулирует оба рефлекса следующим образом: сначала развивается вестибуло-глазной рефлекс и затем подкрепляется оптокинетическим рефлексом. В течение более длительного вращения с открытыми глазами вестибулярный компонент постепенно прекращается, поскольку прекращается движение эндолимфы. Оптокинетический нистагм при этом сохраняется, поскольку сохраняется активность нейронов вестибулярных ядер.
Структурно оптокинетический нистагм обеспечивается многими образованиями мозга. В его формировании принимают участие:
- лобно-височная кора,
- задняя часть теменной коры,
- прецентральная и задняя срединная лобная извилины,
- передний и задний островок,
- прелобная кора и средняя часть верхней лобной извилины.
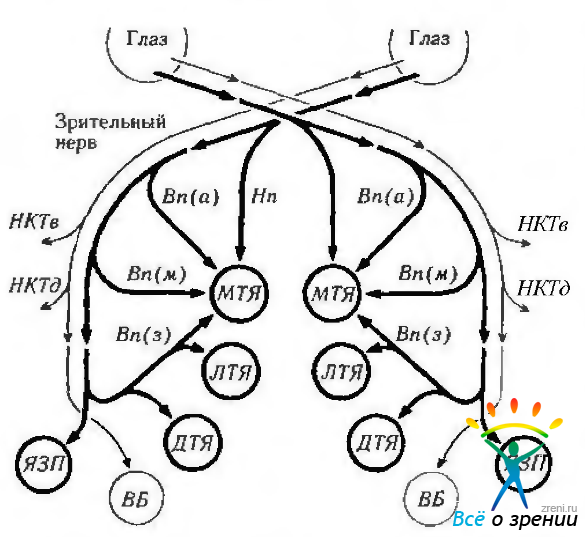
Рис. 4.4.27. Схематическое изображение состава и связей ядер дополнительной системы зрительного тракта (по Lenerstrand et ai, 1982): НКТв — вентральное наружное коленчатое тело; НКТд — дорзальное наружное коленчатое тело; МТЯ — медиальное терминальное ядро; ЛТЯ — латеральное терминальное ядро; ДТЯ — дорзальное терминальное ядро; ВБ — верхние бугорки четверохолмия; ЯЗП — ядро зрительного пути; Нп — нижний пучок, Вп(а) верхняя часть верхнего пучка. Вп(м) медиальная часть верхнего пучка; Вп(з) — задняя часть верхнего пучка
ретикулярные ядра покрышки моста (nuclei tegmenti), а также медиальное ядро моста (nucleus pontis). В процесс вовлечены также п. perihypoglossi и вестибулярная спайка.
Значение коры в оптокинетическом нистагме поддерживается экспериментальными исследованиями. Так, двусторонняя лобэктомия, проведенная у обезьян, вызывает потерю плавного компонента слежения оптокинетического нистагма, а повреждение медиальной верхней височной области (поля 19 и 39) ведет к нарушению следящих движений глаза в сторону повреждения.
У людей с односторонними теменными поражениями выявляется асимметрия плавного слежения.
Выше неоднократно подчеркивалось большое значение мозжечка в регуляции движений глаза. В чем же оно состоит и как осуществляется? Необходимо отметить, что мозжечок только коррегирует движения. Осуществляется это сложной системой связей его со многими структурами мозга. Перед тем как остановиться на этих связях, необходимо кратко напомнить строение мозжечка, частично описанное в первом разделе этой главы.
У высших млекопитающих мозжечок состоит из нового мозжечка, или неоцеребеллума (полушария и участки червя), старого, или палеоцеребеллума (участки червя, соответствующие передней доле, пирамиды, язычок и парафлокулярные отделы), и древнего мозжечка, или архицеребеллума (клочково-узелковая доля; lobus flocculonodularis (рис. 4.4.29).

Рис. 4.4.29. Вентральная поверхность мозжечка после удаления моста и продолговатого мозга (по Strother, Salomon, 1977): 1 — передняя долька; 2— язычок; 3— центральная долька; 4 — первичная щель; 5 — задне-верхняя щель; 6 — горизонтальная щель; 7 — скат; 8— задняя четырехугольнaя долька; 9 — верхняя полулунная долька; 10— извилина червя; 11 — ножка I; 12— нижняя полулунная долька; 13— бугорок червя; 14 - ножка II: 15 — двубрюшная долька; 16 — язычок; 17 — препирамидальная щель; 18 — заднелатеральная щель; 19 — клочок; 20 — узелок; 21 — флоккулонодулярная долька: 22 — парафлоккулус
Эта классификация соответствует его подразделению в зависимости от поступающих афферентных путей. По этой причине архицеребеллум называют вестибулоцеребеллум, палеоцеребеллум — спиноцеребеллум, а неоцеребеллум — понтоцеребеллум.
В белом веществе мозжечка заключено несколько ядер — ядро шатра (n. fastigii), шаровидное (n. globosus), пробковидное (n. emboliformis) и зубчатое (n. dentatus) ядра.
Сложной структурой обладает кора мозжечка (рис. 4.4.30).
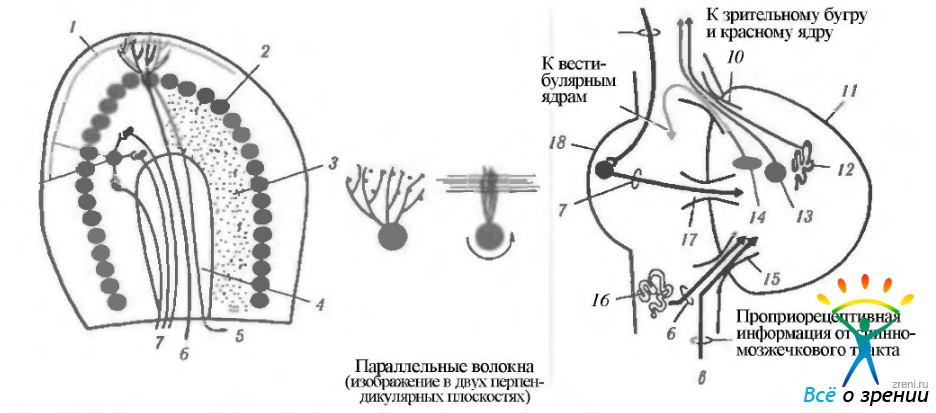
Рис. 4.4.30. Структурная и клеточная организация коры мозжечка (а, б) и основные связи мозжечка с другими структурами мозга (в): 1 — параллельные волокна; 2 — клетки Пуркине; 3 - гранулярный клеточный слой; 4 — белое вещество; 5 волокна в направлении глубоких ядер мозжечка; 6 — лиановые волокна; 7 — мшистые волокна; 8— гранулярные (зернистые) клетки; 9 — корковомостовой тракт; 10—верхняя ножка мозжечка; 11 — мозжечок; 12—зубчатое ядро; 13 — nucl. interposes; 14 — ядро шатра; 15 - нижняя ножка мозжечка; 16- верхняя олива; 17 - средняя ножка мозжечка; 18 - мост
В различных слоях коры расположены шесть типов нервных клеток и их отростки. Мелкие клетки-зерна лежат в зернистом слое, а аксоны их идут в молекулярный слой. Здесь эти аксоны Т-образно делятся, посылая в каждом направлении вдоль поверхности коры ветвь (параллельное волокно) длиной 1—2 мм. Эти ветви проходят через области ветвления остальных типов нейронов и образуют на них синапсы. В зернистом слое расположены клетки Гольджи, дендриты которых распространяются в молекулярном слое, а их аксоны направляются к клеткам-зернам.
Между этими двумя слоями лежит слой нейронов Пуркине, дендриты которых разветвляются в молекулярном слое. Аксоны клеток спускаются к ядрам мозжечка, и небольшое их количество заканчивается в вестибулярных ядрах.
Остальные три типа клеток — корзинчатые, звездчатые и клетки Лугаро — лежат в молекулярном слое. Аксоны этих клеток оканчиваются на теле и дендритах нейронов Пуркине.
В кору мозжечка входят два типа двигательных волокон. Лазящие (лиановые) волокна проходят через зернистый слой и заканчиваются на дендритах клеток Пуркине. Тела клеток, от которых отходят эти волокна, лежат в нижней оливе. Все остальные афферентные пути мозжечка представлены мшистыми волокнами, оканчивающимися на клетках-зернах. Каждое мшистое волокно отдает множество коллатералей и одно такое волокно иннервирует множество клеток коры мозжечка.
В мозжечок входят и из него выходят крупные пучки нервных волокон (рис. 4.4.30). От ядер моста идут волокна в кору противоположной стороны мозжечка. Этот пучок афферентных волокон называется средней ножкой мозжечка (pedimcuhis cerebellaris medialis). Афферентные волокна образуют и веревчатое тело, т. е. нижнюю ножку мозжечка (pedunculus cerebellaris inferior). По ней из спинного мозга поступает информация о положении конечностей и о состоянии тонуса мышц.
Массивный пучок волокон входит также в контрлатеральное веревчатое тело, а затем в ядра нижней оливы (n. olivaris) продолговатого мозга. Другие волокна в составе нижней ножки мозжечка принадлежат клеткам ретикулярной формации.
Верхняя ножка мозжечка (pedunciilus сеrеbellaris superior) состоит в основном из эфферентных волокон, проводящих импульсы из мозжечка к образованиям ствола мозга, промежуточного мозга и к полушариям головного мозга (рис. 4.4.30, в).
Мозжечок контролирует движения путем координации сокращения поперчнополосатой мышечной ткани тела, включая наружные мышцы глаза. Подобное управление требует поступления к мозжечку афферентов от зрительных и вестибулярных ядер, проприорецепторов мышц и сочленений шеи, а также проекции эфферентов на надъядерные зоны, обеспечивающие движения глаза (плавное слежение и саккады).
Наиболее важными связями являются связи с отдельными структурами мозжечка, приведенные ниже.
Клочково-узелковая доля (lobus flocculo-nodularis) (см. рис. 4.4.29).
Почти все пути от вестибулярной системы оканчиваются в узелке (nodulus) и клочке (flocculus). Давно известен передний спинномозжечковой путь (tractus spinocerebellaris anterior) (путь Говерса). Но кроме него известны еще, по крайней мере, 10 путей. Остальные пути представляют собой спинно-оливные пути (tractus spinoolivaris), переключающиеся в оливах на нейроны, посылающие лиановидные волокна к коре мозжечка.
Все пути оканчиваются главным образом в палео- или спиноцеребеллуме. К старому мозжечку идут также соматосенсорные афферентные волокна от головы и зрительные и слуховые афференты.
Зрительная информация достигает клочка через связи, вовлекающие сетчатку, претектальную область противоположной стороны, нижнее ядро оливы.
Проекции зрительной коры на ядра моста образуют синапсы в каждом околоклочке (paraflocculus) (мшистые волокна).
Ядро зрительного тракта и некоторых нейронов клочка кодируют информацию о направлении и скорости движения цели, соответствии движения тела движениям глаза при слежении за целью («сила обратной связи»).
Вестибулоцеребеллум модулирует плавное слежение и вестибуло-глазной рефлекс. Дорзальные повреждения вестибулоцеребеллума у человека приводят к потере координации движений глаза с появлением «промахивающихся саккад».
Червь (Vermis). Дорзальный червь, особенно дольки V, VI и VII. и соседнее ядро шатра участвуют в саккадических движениях. Об этом свидетельствует то, что клетки Пуркине червя возбуждаются за 25 мс перед саккадами или во время саккад. Стимуляция этой зоны приводит к содружественным саккадическим движениям.
Афферентные пути состоят из проекций на червь вестибулярных ядер и основных ядер моста. Ядра моста, в свою очередь, получают зрительные афференты от верхних бугорков четверохолмия, наружного коленчатого тела и стриарной коры. Ймеются также шейные и глазные проприоцептивные афферентные волокна.
Эфферентные пути косвенно проецируются на глазодвигательные ядра через вестибулярные связи.
Полушария мозжечка (hemispherium cerebelli). Проекции полушарий мозжечка складываются из волокон, оставляющих зубчатые ядра и через соединительное плечо достигающих глазодвигательных ядер. Проекция на ядра глазодвигательного нерва происходит непосредственно или после переключения в ретикулярном ядре моста (nucleus reticularis tegmenti pontis), которое лежит в парамедианной ретикулярной формации моста с вентральной стороны. О роли полушарий в движении глаз свидетельствует хотя бы тот факт, что стимуляция полушарий может приводить к саккадическим движениям глаза или плавному слежению.
Таким образом, нарушения функции мозжечка довольно разнообразны в своих клинических проявлениях. Диффузные паренхиматозные поражения возникают вследствие вирусного энцефалита, абсцессов, развития опухоли (медуллобластома, астроцитома), а также наследственных заболеваний типа гемангиобластомы, болезни Гиппель—Линдау (von Hippel—Lindau), мозжечковой дегенерации.
Повреждение передних частей червя приводит в атаксии и затруднению взгляда кверху. Повреждение срединных структур мозжечка сопровождается гипотонией мышц разгибателей конечностей со стороны поражения.
После разрушения мозжечка возможно и развитие повышенного мышечного тонуса, но на короткое время. Это связано с тем, что ядра мозжечка обычно проявляют тормозное влияние на латеральное вестибулярное ядро (nucleus vestibularis lateralis; Дейтерса), которое, в свою очередь, возбуждают альфа-мотонейроны мышц конечностей.
Боковые полушария мозжечка влияют на тонус мышц флексоров через верхнюю ножку мозжечка (соединительное плечо), красное ядро противоположной стороны и красноядерно-спинномозговой путь с этой же стороны. Поскольку имеется двойной перекрест перечнеленных путей (за исключением тракта красного ядра), повреждение полушарий мозжечка приводит к гипотонии со стороны поражения. В то же время повреждение красного ядра сопровождается гипотонией противоположной стороны.
Офтальмолог должен помнить и о так называемом глазном миоклоническом треугольнике, образованном полушарием мозжечка, красными ядрами и нижним ядром оливы моста. Эти структуры связаны между собой мозжечковыми ножками и центральными покрышечными путями. Поражение этой области (чаще инфаркт в области моста) приводит к развитию глазной и небной миоклонии.
Продолжение в следующей статье: Движения глаз ? Часть 3
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0