Современные данные, обосновывающие и уточняющие теорию трехкомпонентности цветового зрения. Часть 3.


Содержание:
Описание
↑ О наличии различных светореагирующих веществ в колбочках
Оставаясь на почве фотохимического понимания, мы оказываемся перед новым вопросом, как же надлежит представлять себе различие трех цветореагирующих («цветоощущающих») аппаратов сетчатки, трех видов колбочек? Самый простой ответ на этот вопрос был дан Гельмгольцем, а позднее развит Лазаревым и некоторыми другими учеными. Ответ этот состоит в допущении, что в колбочках сетчатки имеются три светочувствительных вещества с различными спектрами поглощения. «В концевых аппаратах волокон зрительного нерва имеются три рода фотохимически разлагаемых веществ, имеющих различную чувствительность по отношению к различным частям спектра»,— читаем у Гельмгольца (1896,). Лазарев же (1916,) пишет: «...мы должны допустить, что пигментов должно быть три».
Максимум поглощения одного пигмента должен лежать в красной части спектра, максимум поглощения другого — в зеленой и третьего — в синей части спектра. Данные Блисса и других авторов, находивших, что колбочки сетчаток характеризуются кривой поглощения, имеющей один максимум (около 560 m?), допущению существования трех таких светопоглощающих веществ отнюдь не противоречат. Смесь трех веществ с максимумами поглощения, лежащими в трех различных участках спектра, может давать в качестве результирующей кривую лишь с одним максимумом.
Считая вероятным, что в колбочках сетчатки имеются три различных светореагирующих вещества, естественно думать, что одни колбочки содержат в себе одно из этих веществ, другие — другое и третьи — третье. При таком допущении можно говорить о красно-, зелено- и синереагирующих колбочках как о концевых аппаратах зрительного нерва, позволяющих разлагать попадающие в глаз излучения на три составляющих и посредством этого разложения дающих возможность воспринимать цвета предметов.
Только что описанное представление об устройстве фотохимических рецепторных аппаратов цветового зрения не является, однако, общепризнанным. Имеются возражения против допущения существования в колбочковой сетчатке трех светореагирующих веществ и трех видов колбочек, различных по содержащемуся в них светореагирующему веществу. На основании общих соображений и некоторых экспериментальных данных Федоровы (1939), например, высказываются за то, что в колбочках лишь одно реагирующее на свет вещество, но три нервных окончания, связанные с тремя разными центрами, ощущающими красный, зеленый или синий цвета. Эти нервные окончания лежат в разных местах (на разных уровнях) наружного членика колбочек. Внутренние же членики колбочек обусловливают то, что падающие на сетчатку лучи различной преломляемости концентрируются на том или ином уровне наружного членика колбочек. Благодаря этому лучи разной длины волны возбуждают в различной мере красно-, зелено-и синереагирующие окончания зрительного нерва в колбочках, несмотря на то, что колбочки содержат лишь одно светореагирующее вещество (Федоров, 1937).
Изложенная точка зрения нам кажется мало вероятной. Если бы возбуждение того или иного цветореагирующего аппарата в колбочке определялось степенью схождения (конвергенции) лучей в ней, то следовало бы ожидать, что изменение аккомодации глаза должно было бы менять видимую цветность светового раздражителя, чего на самом деле нет. Удаленная белая поверхность видится нами белой, аккомодируем ли мы на нее или на более близкое расстояние.
Могло, далее, показаться, что допущение одного светореагирующего вещества во всех колбочках можно примирить с их способностью различно реагировать на различные длины волн спектра, если признать, что перед колбочками имеются различные светофильтры: одни из них пропускают преимущественно лучи красные, другие — зеленые и третьи — синие. Мы действительно встречаем в сетчатках некоторых животных (черепах, ящериц, птиц) нечто подобное в виде цветных маслянистых шариков, расположенных во внутренних члениках колбочек. Цвет этих шариков чаще всего красный, оранжевый и желтый, реже синеватый и зеленовато-желтый. Последние два цвета некоторыми авторами (Уоллс), впрочем, относятся за счет несовершенств микроскопической оптики. В сетчатке человека цветных маслянистых шариков, однако, не найдено.
Ряд фактов, установленных в специальных экспериментах, может также служить аргументом в пользу допущения в сетчатке не одного, а трех цветореагирующих веществ. Нами изучался ход спадания яркости цветового ощущения при длящемся действии монохроматического раздражения на глаз (Кравков, 1928). Речь шла, следовательно, о ходе цветового утомления глаза к применявшемуся спектральному раздражителю. В спектральном аппарате (аппарате Гельмгольца для смешения цветов) в одной половине поля зрения давалось «утомляющее» раздражение, яркость которого при длящемся действии раздражителя постепенно спадала; в другой половине имелась та или иная стационарная яркость излучения той же длины волны. Стационарность этой яркости достигалась тем,что свет предварительно уже действовал на глаз в течение 90 сек. Подопытные лица должны были отметить время, когда яркость первой половины поля зрения (бывшая всегда большей в первый момент действия раздражителя на глаз) делалась равной яркости второй («стационарной») половины. В качестве утомляющих раздражителей, затухающая яркость которых определялась, были взяты две пары подравненных по яркости спектральных цветов: красный (656 m?) и синий (451 m?) и красный (656 m?), но более яркий, чем в первой паре, и зеленый (550 m?).
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
В опытах на четырех лицах было найдено, что спадание яркости (или, соответственно, снижение чувствительности) для спектральных раздражителей различной длины волны идет с различной скоростью. Чувствительность быстрее всего снижается для синего и медленнее всего для зеленого раздражителя (рис. 23).
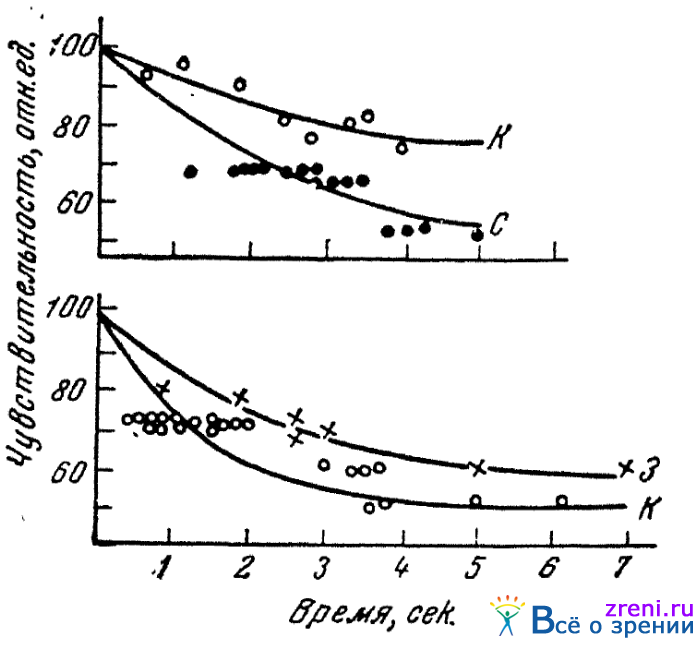
Рис. 23. Кривые цветовой адаптации к различным цветам (по Кравкову)
Найденное неодинаковое спадание чувствительности для разных цветореагирующих аппаратов глаза свидетельствует о существовании разных химических реакции, являющихся началом возбуждения этих аппаратов, т. е. говорит в пользу признания существования нескольких различных светореагирующих аппаратов в колбочках сетчатки. Фотохимический распад трех светореагирующих веществ, будучи пропорционален количеству поглощенной энергии, на стационарном уровне ощущения имеет для красно-, зелено- и синереагирующих аппаратов глаза различные коэффициенты пропорциональности.
Аналогичные опыты были поставлены позже Федоровыми (1939). Они не нашли какой-либо разницы в скорости спадания яркости цветных раздражителей различного цвета. Причина этого несогласия нам не ясна. Критические замечания Федоровых относительно того, что в наших опытах подравнивание «утомляющего» раздражителя другому (стационарной яркости) производилось лишь по яркости (цветовой же тон и насыщенность могли оказаться неподравненными), не могут опровергнуть наших выводов. Дело в том, что именно изменение яркости при длящемся раздражителе и является наиболее резко выраженным изменением ощущения, происходящим вследствие падения чувствительности.
За существование в колбочках разных светореагирующих веществ говорит, далее, и факт различной скорости развития цветовых ощущений от раздражителей, затрагивающих различные цветореагирующие аппараты глаза. Различной скоростью развития цветовых возбуждений объясняются некоторые интересные явления, в существовании которых можно убедиться в простых опытах. Так, если привести во вращение диск, разделенный на белые и черные секторы (рис. 24),

Рис. 24. Диск для демонстрации различной скорости возникновения цветовых ощущений
то при некоторой, не слишком большой скорости в наружном кольце диска мы увидим красноватый налет, а во внутренней части — синеватый оттенок. При ускорении вращения красноватость переходит и на внутреннюю часть диска. Понятно, что продолжительность действия белого раздражителя на глаз будет тем меньше, чем меньше угловой размер белого сектора. Из содержащихся в белом свете лучей с различной длиной волны красные лучи, следовательно, скорее дают себя почувствовать, чем синие.
То же самое сказывается в кажущемся пространственном смещении относительно друг друга двух полос, если одна из них — синяя, а другая — красная и обе они наклеены по радиусу на черный вращающийся диск. При вращении диска кажется, что синяя полоска несколько отстает от красной.
Известен также опыт с так называемым диском Бенхэма (рис. 25).
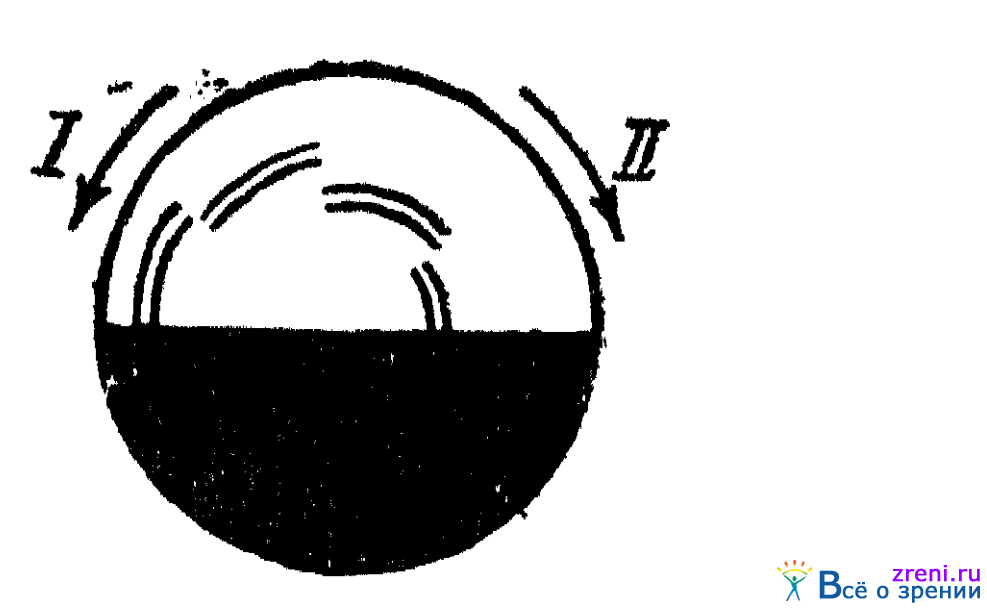
Рис. 25. Диск для демонстрации различной скорости возникновения цветовых ощущений
Половина диска закрашена черным, на белой половине нанесены черные дуги по 45° каждая. Нанесены они по паре на разных расстояниях от центра, как показано на рисунке. При некоторой скорости вращения диска дуги кажутся цветными. При вращении диска в направлении стрелки I эти пары дуг (по порядку — от внешних периферических к центральным) кажутся красноватого, желтоватого, зеленоватого и сине-фиолетового оттенка. При вращении диска в направлении стрелки II порядок цветов на кольцах будет обратным, т. е. внешние дуги кажутся фиолетово-синеватыми, центральные — красноватыми. Вращение диска должно происходить с некоторой, не очень большой и не очень малой скоростью, иначе описанное явление наблюдаться не будет.
Возникновение хроматических цветов в дугах на диске объясняется тем, что при начале действия белого цвета на глаз, в каждом из 45-градусных секторов белого полукруга возникает тот или иной хроматический цвет в силу того, что все три цветореагирующих аппарата глаза возбуждаются с различной скоростью. Цвет же дуг является цветом, видимым по контрасту к этим хроматическим оттенкам фона.
Что касается скорости возбуждения цветореагирующих аппаратов глаза, то из анализа опытных данных Пьерона (1925) следует прийти к выводу, что вероятный порядок цветовых возбуждений в направлении убывающей скорости таков: зеленый, красный, синий. Недавние опыты Мкртычевой (1949), посвященные изучению скорости нарастания цветовых возбуждений, принесли прямое экспериментальное подтверждение этому заключению. Для фовеального зрения скорость нарастания возбуждения для зеленого (520 m?) света оказалась большей, чем для красного (650 m?); для синего же (470 m?) была найдена наименьшая скорость.
В связи с основной линией наших рассуждений важно подчеркнуть, что простым объяснением различной скорости возникновения цветовых ощущений может служить допущение нескольких различных светореагирующих веществ в колбочках, веществ, характеризуемых различными константами для происходящих в них химических реакций. Поскольку, таким образом, следует допустить несколько светореагирующих веществ в колбочках, можно, в согласии с теорией трехкомпонентности, принять это число равным трем.
Укажем, далее, на известный факт затухания цветового последовательного образа от белого раздражителя. Если посмотреть на яркий белый свет и перевести глаз в темноту, то в течение некоторого времени мы будем видеть светлый последовательный образ, постепенно затухающий. Как описывалось еще старыми авторами (Фехнер, Гельмгольц) и как то подтвердилось позже, после краткого раздражения глаза белым светом последовательный образ, затухая, проходит через фазы зеленоватую, синеватую и красноватую.
Есть основания полагать, что явления последовательных образов в значительной мере определяются сетчаточными процессами. За это говорят факты образования последовательных образов и тогда, когда, например, нажимая на глазное яблоко, мы делаем глаз на время невидящим, нарушая нормальное кровоснабжение сетчаточных клеток. Световое раздражение сетчатки такого (невидящего) глаза по восстановлении нормальных условий оставляет после себя последовательный образ. Между тем несомненно, что чувствующие мозговые центры зрительного анализатора световым раздражителем в описанном опыте не возбуждались. Если же последовательный образ определяется в основном сетчаточными процессами, то в качестве таковых естественно представить себе процессы постепенного удаления из сетчатки продуктов фотохимического распада светореагирующих веществ сетчатки. Разработана теория подобного рода процессов (Лазарев, 1950) и выведена формула, определяющая ход затухания последовательного образа по окончании светового раздражения. Эта формула включает коэффициент ?3, характеризующий скорость уноса из сетчатки раздражающих продуктов распада.
Из различной скорости затухания последовательных образов для разных цветовых возбуждений вытекает, таким образом, необходимость признать различие в величине коэффициента ?3 для разных цветореагирующих аппаратов глаза, что говорит за вероятность существования в колбочках различных светореагирующих веществ.
Обнаружение и изучение различных цветореагирующих веществ является одной из очередных, ждущих своего разрешения задач физиологии цветового зрения.
↑ Кривая чувствительности колбочкового зрения к различным лучам спектра
Согласно воззрениям современной физики, свет излучается всегда определенными порциями — световыми квантами (фотонами). Энергия светового кванта равняется hv, где v —число колебаний электромагнитной волны в секунду, h — некоторая постоянная, равная 6,54-10“27 эрг/сек. Чтобы в светореагирующем веществе сетчатки наступили фотохимические изменения, нужно, чтобы фотон был поглощен молекулой этого вещества и вызвал ее распад. Вероятность подобного события зависит, с одной стороны, от природы светореагирующего вещества, с другой — от длины волны действующего света, или, что то же, от энергии фотона. Очевидно, что, чем больше такая вероятность, тем меньше энергии излучения потребуется для того, чтобы вызвать фотохимический распад данного вещества. Но так как с распада светореагирующего вещества колбочек начинается процесс возбуждения цветового зрения, то, чем больше вероятность такого распада, тем больше чувствительность нашего колбочкового зрения к тому или иному световому раздражителю.
Многочисленные опыты установили, что чувствительность нашего колбочкового зрения к различным спектральным лучам не одинакова. Это зависит, правда, не только от различной вероятности распада молекул светореагирующих веществ под влиянием фотонов той или иной энергии, но и от различной прозрачности для тех или иных спектральных лучей глазных сред, лежащих впереди колбочек. Кроме того, поскольку отдельные цветореагирующие вещества колбочек не выделены, возможно определять чувствительность лишь суммарно для всех имеющихся в колбочках веществ. Тем не менее, кривая спектральной чувствительности колбочкового зрения, или, как ее еще часто называют, кривая видности спектральных лучей обнаруживает весьма правильную форму. Это колоколообразная довольно симметричная кривая. Результаты определения чувствительности колбочкового зрения к различным лучам спектра, произведенного Пинегиным в Гос. Оптическом институте в Ленинграде, приведены на рис. 26.
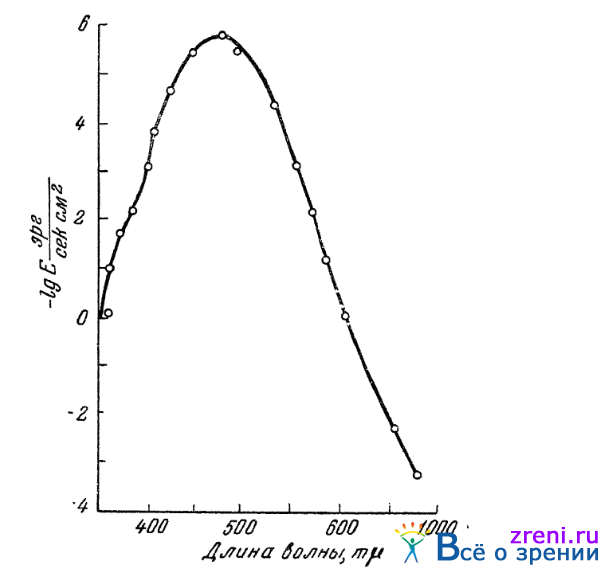
Рис. 26. Кривая спектральной чувствительности колбочек (по Пинегину)
По ординате отложены отрицательные значения силы порогового раздражителя:
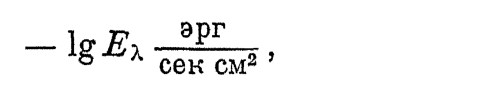
где Е — соответствующая порогу ощущения освещенность на зрачке; таким образом, чем выше ордината кривой, тем больше чувствительность колбочек к данной длине волны. По абсциссе отложены длины волн равноэнергетического спектра, чувствительность к которым определялась. Максимум кривой приходится приблизительно на 550 m?. — зеленовато-желтые лучи. К ним наши колбочки и оказываются наиболее чувствительными.
По данным других авторов (Федоровы, 1936), положение максимума колбочковой чувствительности соответствует несколько более длинноволновым лучам, равняясь (в зависимости от условий) 550—565 m?, что тоже лежит в области зеленовато-желтых лучей.
По измерениям Пинегина, минимум световой энергии, способный еще вызвать ощущение хроматического цвета, близок к 100 квантам в секунду (для ? = 546 m?). Границы же чувствительности колбочек по спектру простираются от лучей 302 m? в области лучей ультрафиолетовых до 950 m? в области лучей инфракрасных. Правда, чувствительность к этим крайним видимым лучам в сотни тысяч и миллионы раз меньше, чем чувствительность к зеленовато-желтым лучам.
Нельзя не заметить, что положение максимума чувствительности колбочкового зрения в области зеленовато-желтых цветов является биологически целесообразным для существ, живущих среди растительности в условиях солнечного освещения.
«Следует заметить,— пишет по этому поводу С. И. Вавилов (1950),— что кривая видности для дневного зрения почти совпадает со средней кривой распределения энергии солнечного света, отражаемого и рассеиваемого зелеными растениями. Это обстоятельство, конечно, весьма выгодно для существа, живущего среди растений и в значительной мере питающегося ими.
Перед нами пример удачного приспособления глаза к реальным условиям жизни на Земле и доказательство действительного родства глаза и Солнца».
↑ Электрофизиологичеcкие изменения в зрительном нерве и мозговых центрах
Свет, поглощенный молекулой светореагирующего вещества, может вызывать распад молекулы, что в свою очередь производит изменения в электрическом состоянии вещества и может являться началом нервного возбуждения. Возбуждение распространяется по нервам выше, к зрительным центрам в мозгу — в подкорке и в коре.
Возбуждение нерва сказывается появлением в нем электрических импульсов, вызываемых изменяющимся электрическим потенциалом. Колебания этого потенциала могут быть записаны на достаточно чувствительном электрорегистрирующем приборе — осциллографе в виде «токов действия». Многочисленные исследования записи биотоков нервов вообще и, в частности, зрительного нерва показывают, что при регистрации хода возбуждения в одном нервном волокне величина отдельных импульсов от силы раздражителей не зависит; от силы раздражителя меняется частота импульсов, пробегающих по нервному волокну. На рис. 27
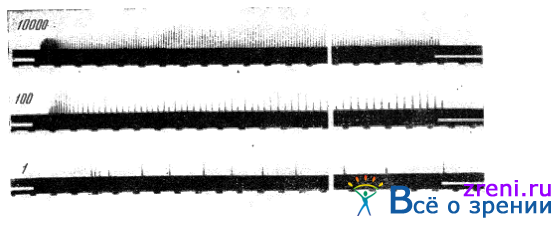
Рис. 27. Биотоки в зрительном волокне в зависимости от яркости светового раздражителя
показаны осциллографические записи биотоков в отдельном волокне зрительного нерва ракообразного Limulus (по Граниту). В каждой строке этого рисунка приведены биотоки, возникшие от светового раздражителя различной силы. На нижней строке сила раздражителя (в условных единицах) равнялась 1, на средней 100, на верхней 10 000. Ясно видно учащение импульсов по мере усиления светового раздражителя.
Применительно к процессам возбуждения нашего цветового зрения мы, очевидно, должны допустить, что под влиянием раздражения глаза световым раздражителем того или иного спектрального состава в нервных волокнах, связанных с красно-, зелено- и синеощущающими рецепторами глаза, будут
возникать электрические импульсы той или иной частоты. Соотношение этих частот, определяемое цветом рассматриваемого предмета, будет определять и наше цветовое ощущение.
Особенности в картине биотоков зрительного нерва в связи с цветностью раздражителя только лишь начинают изучаться. Некоторыми авторами (Donner, 1949) отмечается, например, что импульсы, возникающие в нерве при раздражении глаза световыми раздражителями, действующими на зрительный пурпур одинаково, дают наибольшую частоту в различные периоды времени после своего начала, и это различие зависит от цветности раздражителя. По названному автору, раздражение красным светом (600 m?) дает максимальную частоту импульсов в ближайший момент после начала разрядов, синее же раздражение (460 m?.) — заметно позже, зеленое (520 m?) занимает промежуточное место.
Начальная стадия зрительного возбуждения сказывается на электрическом потенциале сетчатки. Изменения этого потенциала могут быть записаны струнным гальванометром или осциллографом в виде электроретинограммы. Такие записи производились многими учеными. При этом один электрод соприкасался с роговицею исследуемого глаза, а другой прикладывался вне глаза, к какому-нибудь месту поверхности тела (например, к мочке уха, нёбу и т. п.). В связи с проблемами цветового зрения было интересно выяснить, как зависят электрические процессы в сетчатке специально от цветности (длины волны) светового раздражителя. Следует, однако, признать, что на пути такого электрофизиологического изучения закономерностей цветового зрения сделано пока еще очень мало. Изучить в достаточной мере различия в форме ретинограммы, которые зависели бы от цветности раздражителя, еще не удалось. Сакс (Sachs, 1929) отмечает лишь, что у цветнослепых типа протанопов (или, как их еще называют, «краснослепых») красные лучи спектра при объективно одинаковой интенсивности вызывают заметно меньший электродвигательный эффект в сетчатке, чем у лиц с нормальным цветоощущением.
В еще не опубликованных опытах Зарецкой и Стручкова, выполненных в нашей лаборатории, было найдено, что раздражение красным светом (небольшого участка сетчатки, лежащего на 2° к периферии от центральной ямки) дает ретинограмму, отличную от получающейся при раздражении того же места сетчатки зеленым или белым светом той же яркости. Сказанное иллюстрирует рис. 28,
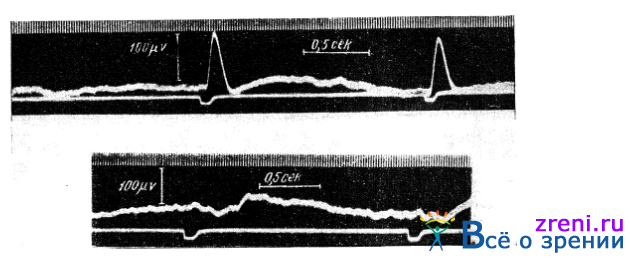
Рис. 28. Ретинограммы при раздражении глаза зеленым (верхняя ретинограмма) и красным (нижняя) светом (по Зарецкой и Стручкову). Время раздражения отмечено на нижней линии. Интервалы времени в 0,02 сек. отмечаются сверху
где приведены ретинограммы, полученные названными выше авторами при раздражении глаза испытуемого зеленым (а) и красным (б) светом. Обе ретинограммы записаны во время одного сеанса в тождественных условиях (кривая б через 1 мин. после кривой а).
↑ О центральных частях цветореагирующих аппаратов зрительного анализатора
Не подлежит сомнению, что различные цветоощущающие аппараты зрительного анализатора, периферической частью которых являются три вида колбочек в сетчатке, должны иметь свою особую анатомическую локализацию и в выше лежащих участках нервной системы. Такими участками для нашего зрительного анализатора являются в промежуточном и среднем мозге подушка зрительного бугра, наружное коленчатое тело и переднее четверохолмие, а в коре головного мозга — затылочная область, охватывающая так называемую борозду птичьей шпоры (fissura calcarina). Из промежуточных центров с функцией чувствительности связано, по-видимому, лишь наружное коленчатое тело.
Были попытки [Кларк (Clark), 1940] связать слоистое строение наружного коленчатого тела с тройственностью цветоощущающих аппаратов глаза. Именно в наружных коленчатых телах у человека и обезьян Кларк наблюдал по три слоя, связанных со зрительными волокнами каждого глаза. Гипотеза цитируемого автора, полагавшего, что каждый из этих слоев является анатомическим субстратом одного из трех цветоощущающих аппаратов глаза, не нашла широкого признания. Указывалось [Уоллс (Walls), 1942] между прочим и на то, что подобное же слоистое строение наружного коленчатого тела имеется и у некоторых животных, ведущих ночной образ жизни и цветовым зрением, по-видимому, не обладающих.
Что касается коры головного мозга, то гистологическими исследованиями Хеншена (Henschen, 1929), установлено в IV слое зрительной области коры головного мозга наличие двух видов ганглиозных клеток: клеток большего размера, с круглыми ядрами и хуже прокрашивающихся, и клеток меньшего размера, с ядрами овальной формы и лучше прокрашивающихся. Сравнительное гистологическое изучение срезов мозга дневных и ночных обезьян показало, что с дневным зрением и цветоощущением связан именно последний вид ганглиозных клеток.
Поскольку цветовое зрение есть функция преимущественно центральной области сетчатки, последняя же представлена в задних частях затылочной области коры головного мозга, можно думать, что там же главным образом локализован и корковый субстрат цветового зрения.
Несомненно, что анатомический субстрат цветового зрения в мозге в известной мере обособлен от субстрата нашей способности видеть яркостные свойства предметов, т. е. ощущать светлоту. Описаны случаи, когда, в силу тех или иных нарушений мозга, у людей утрачивалось цветоощущение, между тем как способность видеть посредством колбочек белый цвет сохранялась. Потеря цветоощущения при сохранности светоощущения порою захватывает лишь половину поля зрения и носит название гемиахроматопсии.
В годы войны нам пришлось обследовать двух черепномозговых раненых, у которых после ранения было полностью потеряно цветовое зрение, зрение же посредством палочек, зрение сумеречное, осталось нормальным. Относительно обоих этих раненых было достоверно известно, что до ранения они обладали цветовым зрением и лишились его лишь после мозговой травмы.
---
Статья из книги: Цветовое зрение | Кравков С. В.

Комментариев 0