Об особенностях различных цветореагирующих аппаратов зрительного анализатора. Часть 2.


Содержание:
Описание
↑ Об участии ацетилхолинового звена в реакциях колбочкового аппарата глаза на непрямые раздражители
Ряд физиологических работ (см. Михельсон, 1948) свидетельствует далее о том, что одновалентные катионы К и Na, с одной стороны, и двухвалентные катионы Са и Mg, с другой,— помимо валентности, обладают и еще некоторыми общими свойствами, которые могут иметь значение для функционирования цветоощущающих аппаратов глаза. Этим свойством является характер их действия на холинэстеразу—фермент, который разрушает гидролизует) ацетилхолин в тканях. Ацетилхолин же, по современным представлениям физиологии, есть вещество, выделяемое нервной тканью при возбуждении, и существующее в ней в связанной форме, в виде «запаса». Хотя роль и место ацетилхолина в нервных процессах и не всеми физиологами понимается одинаково, несомненное значение его для интимных процессов функционирования нервной клетки едва ли может быть оспариваемо.
Опыты Бабского, Альперна и др. (см. Михельсон, 1948) показали, что двухвалентные ионы кальция и магния действуют на холинэстеразу активирующим образом. Тем самым они создают условия, неблагоприятные для наличия ацетилхолина, содержание которого от них уменьшается. Ионы же калия и натрия, напротив, угнетают холинэстеразу и тем самым увеличивают содержание ацетилхолина в ткани.
Из описанных выше опытов над изменением цветовой чувствительности нашего зрительного анализатора можно сделать и тот вывод, что зелено- и синеощущающие аппараты глаза лучше всего функционируют (оказываются более чувствительными) при сравнительно меньшем содержании ацетилхолина в глазе; аппарат же красноощущающий, напротив, от увеличения ацетилхолина в глазе в своей возбудимости выигрывает.
В пользу того, что в найденном нами действии ионов на чувствительность колбочкового аппарата нашего органа зрения играет какую-то роль и ацетилхолиновое звено, говорят недавние опыты Поликаниной (1950). Подвергая глаз действию гоматропина и испытывая затем действие ионофореза натрия, Поликанина нашла, что гоматропинизированный глаз не обнаруживает сколько-нибудь заметных сдвигов в своей цветовой чувствительности в ответ на такой ионофорез. Обнаружилось, следовательно, что гоматропинизация глаза устраняет его реактивность на данное непрямое раздражение. Рис. 59
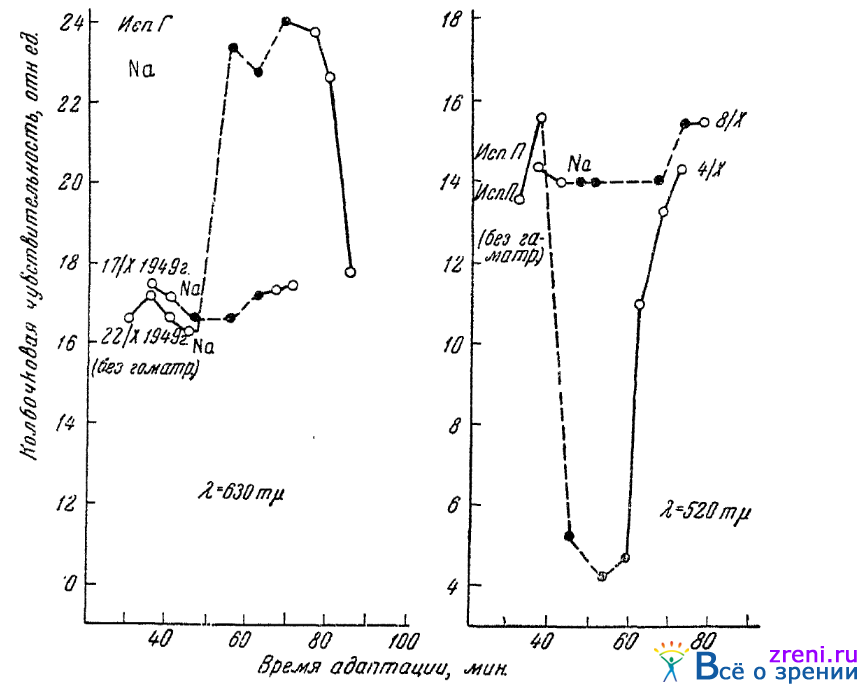
Рис. 59. Отсутствие реакции на ионофорез колбочковой чувствительности гоматропинизированного глаза (по Поликаниной); черными кружками обозначены величины, найденные во время ионофореза
воспроизводит типичные результаты описываемых опытов. Из фармакологии же известно, что гоматропин препятствует соединению ацетилхолина с химическими веществами рецептивной субстанции эффекторной клетки. Описанные выше результаты опытов, проведенных над реакциями чувствующих, цветоощущающих клеток, позволяют считать вероятным, что и по отношению к ним изменение в содержании ацетилхолина перестает иметь значение, если в глаз введен гоматропин.
Итак, в итоге рассмотренных выше опытных данных мы вправе сказать, что «ионная» и «ацетилхолиновая» природа цветоощущающих аппаратов, обслуживающих ощущение нами коротковолновой и длинноволновой частей спектра, различна в выше упомянутом смысле.
↑ О возможности нейтрализации действия непрямых раздражителей ионными изменениями в глазе
Описанные опыты позволяют сделать и еще некоторый, немаловажный, как нам кажется, вывод. Было показано (Поликанина, 1950), что влияние на цветовое зрение непрямых раздражителей, действующих несомненно центрально, может быть нейтрализовано изменением ионного состояния глаза, вызываемым ионофорезом. Так на рис. 60
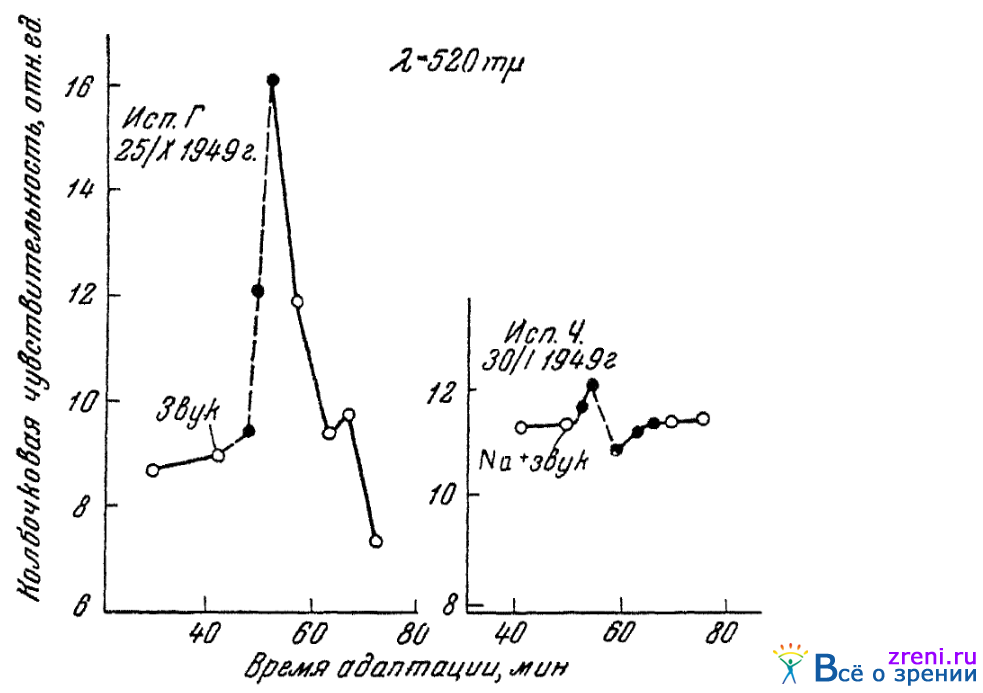
Рис. 60. Одновременное действие слухового раздражения и ионофореза натрия на колбочковую чувствительность глаза; черными кружками обозначены величины, найденные во время применения непрямых раздражителей (по Поликаниной)
воспроизводятся результаты некоторых еще неопубликованных опытов Поликаниной, в которых на подопытного субъекта одновременно воздействовали звуковым раздражителем и на глаз — ионофорезом натрия. Ионофорез натрия, примененный отдельно, обычно вызывает снижение колбочковой чувствительности глаза к зеленому (520 m?), звуковое же раздражение, также примененное отдельно, обычно повышает эту чувствительность. В опытах, когда оба раздражителя применялись одновременно, наблюдалось отсутствие какого-либо заметного сдвига чувствительности. Повышающее чувствительность влияние «с центров» нейтрализовалось понижающим влиянием, идущим от периферического раздражения глаза ионофорезом.
Все опыты с ионофорезом, о которых шла речь выше, по своим результатам чрезвычайно определенны и легко воспроизводимы, можно сказать, «без отказа». Несмотря на это, вопрос о том, каким же путем подводимые ионофорезом ионы оказывают свое действие, еще ждет своего разрешения. Эффект ионофореза сказывается на колбочковой чувствительности глаза, по нашим данным, уже через 3—4 мин. после начала ионофореза. Трудно предполагать, чтобы при применявшемся нами очень слабом токе (не свыше 0,5 mА) ионы могли за это время достигать сетчатки под влиянием имеющейся здесь разности потенциалов. Скорость передвижения ионов под влиянием этого фактора, как известно, очень незначительна. Можно думать о действии ионов через ток крови и лимфы, куда ионы могут проникать вследствие ионофореза. Вероятным является и допущение нервного, рефлекторного действия, вызываемого ионами, проникающими в кожу век и передние отрезки глаза.
Описанные выше опыты с применением ионофореза вызывали эффекты такие же, какие наблюдались нами в результате электротонического воздействия на глаз. Сопоставление этих фактов и заставляет полагать, что в результате ионофореза — каким бы путем ионы не оказывали свое действие — в средах глаза, в сетчатке, наступают изменения в относительной концентрации соответствующих ионов. С подобным представлением, допускающим, что под влиянием ионофореза, в конечном итоге, изменяется ионный состав сред глаза, хорошо согласуются факты однородных действий ионофореза К и Na, с одной стороны, и Са и Mg — с другой. Вызываемые соответствующими ионами изменения однородно влияют на холинэстеразу.
В заключение мы должны сказать, что ионные сдвиги, производимые ионофорезом в средах глаза, носят, видимо, в значительной мере периферический, местный характер. За такое именно понимание говорят результаты тех описанных выше опытов, в которых в оба глаза одновременно производился ионофорез различных ионов и, соответственно с тем, какой ион вводился, изменения колбочковой чувствительности в обоих глазах носили разный характер.
↑ Сопоставление реактивности цветового зрения на непрямые раздражители с кривыми основных цветовых возбуждений
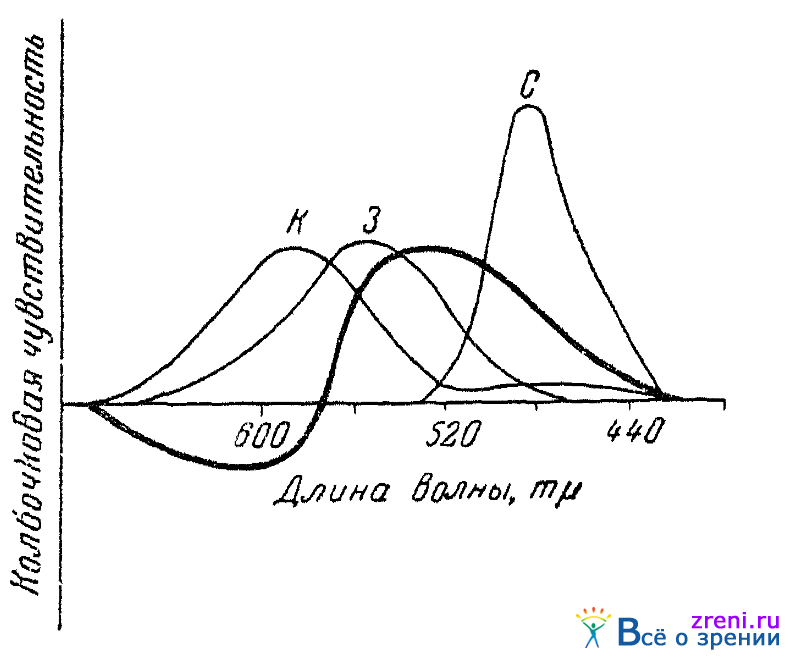
Для объяснения физиологической природы нашего цветового зрения необходимо обратиться к дальнейшему анализу установленной опытами картины действия непрямых раздражителей. Сопоставим для этого эффект действия непрямых раздражителей (например, первой группы их, включающей слуховые раздражители, запахи гераниола и бергамотового масла, анэлектротон, адреналин и т. п.) с тремя кривыми основных цветовых возбуждений глаза. На рис. 61
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Рис. 61. Схематическое сопоставление кривой действия на колбочковую чувствительность симпатикотропных непрямых раздражителей с тремя кривыми основных цветовых возбуждений глаза (по Кравкову)
жирной линией обозначена чувствительность, наблюдаемая при действии указанной группы непрямых раздражителей на темноадаптированный глаз. Горизонтальная линия для этой кривой обозначает уровень чувствительности, имевшийся до применения непрямого раздражителя.
Из рис. 61 непосредственно вытекает следующее. Во-первых, изменения чувствительности глаза происходят под влиянием непрямых раздражителей по отношению не ко всем участкам спектра. «Безразличными» точками являются концевые участки спектра и область желтого (около 570 m?). «Безразличие» их требует объяснения. Во-вторых, рис. 61 показывает, что от всех непрямых раздражителей взятой группы чувствительность по отношению к лучам с длиной волны меньше 570 m? повышается, к лучам же с длиной волны больше 570 m?, наоборот, падает.
Попытка согласовать друг с другом оба эти факта приводит к необходимости признать, что изменения колбочковой чувствительности от всех упомянутых непрямых раздражителей наступают лишь там, где затрагивается зеленоощущающий аппарат нашего зрительного анализатора. Таким образом, приходится сделать важный вывод, что зеленоощущающий аппарат занимает в нашем цветовом зрении особое положение. Приемником влияний, идущих со стороны непрямых раздражений на цветовое зрение, является преимущественно именно он.
Но если дело обстоит так и если, как мы знаем, в области спектра, обслуживаемой главным образом зеленоощущающим аппаратом, при воздействии непрямых раздражителей наблюдается повышение колбочковой чувствительности, то как понять снижение чувствительности в тех же условиях по отношению к лучам оранжево-красной части спектра. Если реагирует на непрямые раздражители лишь зеленоощущающий аппарат и его реакция на непрямые раздражители сказывается повышением чувствительности, то почему также не повышается чувтвительность и к оранжево-красным лучам спектра, ведь при воздействии на глаз этих лучей также затрагивается зелено-ощущающий аппарат.
Объяснить это, по нашему мнению, можно, лишь допустив, что повышение возбуждения зеленоощущающего аппарата вызывает понижение возбудимости аппарата красноощущающего. Результирующее изменение чувствительности колбочек к оранжево-красным лучам под влиянием непрямых раздражителей, повышающих возбуждение зеленоощущающего аппарата, при таких условиях как раз и будет снижением чувствительности. В этой части спектра зрительное впечатление определяется преимущественно красноощущающий аппаратом, который, согласно нашей гипотезе, зеленоощущающим аппаратом угнетается. В области «безразличной» точки — желтого цвета (около 570 m?) — повышение возбуждения зеленоощущающего аппарата и обусловливаемое им понижение возбудимости аппарата красноощущающего, видимо, уравновешивают друг друга, благодаря чему результирующая чувствительность глаза, измеряемая по отношению к излучению с длиною волны 570 m?, и оказывается неизменяющейся.
Если повышение возбуждений зеленоощущающего аппарата угнетает красноощущающий, то с синеощущающим аппаратом зеленоощущающий находится уже в иных отношениях — в отношениях содействия. Повышение возбуждения зеленоощущающего аппарата вызывает повышение возбудимости и в аппарате синеощущающем. Такое следствие вытекает из рассмотрения того), что в области синего, где кривые К и 3 вторично пересекаются, под влиянием побочных раздражителей колбочковая чувствительность заметно повышается. Сравним с этим фактом отсутствие какой-либо реакции на те же непрямые раздражители, если речь идет о чувствительности колбочек к более крайним фиолетово-синим лучам спектра: синеощущающий аппарат глаза в этом случае возбуждается, но зеленоощущающий не затрагивается, а потому|не происходит и какого-либо изменения чувствительности.
↑ Особая природа зеленореагирующего аппарата
Таким образом, изучение изменений цветовой чувствительности, происходящих под влиянием непрямых раздражителей, заставляет признать, что зеленоощущающий аппарат в нашем цветовом зрении занимает особое положение в смысле своей способности реагировать на непрямые раздражителя. Кроме того, мы оказываемся вынужденными допустить существование между отдельными аппаратами нашего зрительного анализатора особых функциональных связей как тормозного, так и содействующего характера.
В пользу признания особой анатомо-физиологической природы зеленоощущающего аппарата глаза можно привести и другие факты. Отмечалось, например [Максвелл (Maxwell), 1918], что чувствительность к зеленому наиболее легко уязвима при контузиях мозга; часто наблюдаемое при этом сужение границ полей зрения на цвета для зеленого цвета бывает выражено особенно значительно. В опытах нашей лаборатории [Кравков (ред.), 1939] наблюдалось, что разностный порог яркости для зеленого цвета иначе зависит от условий адаптации, чем для полей других цветов. При исследовании снижения критической частоты слияния мельканий в темноте для фовеально видимого поля было найдено (Кравков, 1938), что это снижение для лучей зеленой области спектра минимальное; снижение критической частоты для участков спектра, возбуждающих сине- и зеленоощущающие аппараты глаза, гораздо сильнее выражено. Упоминавшиеся уже выше опыты Селецкой, касавшиеся индуктивных процессов в колбочковом зрении, показали, что (для темноадаптированного глаза) выраженность отрицательного индуктивного действия тем больше, чем больше возбуждение зеленоощущающего аппарата. В опытах Шварц (1946), наконец, было показано, что раздражение слабым красным светом повышает чувствительность к зеленому как глаза, освещавшегося красным, так и глаза не освещавшегося. В случае же, если мы наблюдаем сенсибилизацию к красному после освещения зеленым, таковая наблюдается лишь на освещавшемся глазе; на другой глаз сенсибилизирующее действие зеленого не передается. Все эти факты ждут еще своего объяснения. Здесь же мы считали нужным их привести как факты, говорящие о своеобразии зеленоощущающего аппарата, что вытекает и из сделанного выше анализа данных, полученных в результате применения метода непрямых раздражений.
↑ Особенности краснореагирующего аппарата
Особая природа, очевидно, присуща и красно- и синеощущающим аппаратам, поскольку они сами по себе не реагируют на ряд воздействий, вызывающих реакции аппарата зеленоощущающего. Они стоят, как мы пытались показать выше, в различной зависимости от аппарата зеленоощущающего.
Укажем здесь на некоторые особенности красноощущающего аппарата, обзор которых отчасти уже дан в специальной статье Селецкой (1950). Раздражитель крайнего красного цвета (706 m?) не оказывает никакого индуктивного действия на видность близких к нему световых точек других цветов в условиях темновой адаптации. Вместе с тем и другие световые раздражители никак не меняют видность красной точки, возбуждающей лишь красноощущающий аппарат глаза. В условиях световой адаптации подобная «безразличность» крайнего красного раздражителя, как наблюдал Федоров (1949), уже не имеет места. Так именно и должно было быть с точки зрения высказанной нами выше гипотезы. В условиях световой адаптации к белому свету возбужденным оказывается, очевидно, уже не только красноощущающий аппарат, но и прочие два аппарата, в том числе и зеленоощущающий. Последнее же обстоятельство делает крайний красный раздражитель уже подобным не крайнему красному, а оранжевато-красному, поскольку одновременно с красноощущающим прибором возбужден и прибор зеленоощущающий. При таких же условиях, конечно, крайний красный раздражитель и обнаруживает те индуктивные реакции, какие обнаруживает и более коротковолновый красный цветовой раздражитель.
В работе Колесниковой (1947) при исследовании действия блеских раздражителей на видность цветовых полей разного цвета найдено, что видность красного поля (670 m?) в широких пределах не зависит ни от близости, ни от яркости блеского источника, между тем как в тех же опытах видность полей других цветов заметно изменяется при изменении упомянутых условий.
Жанделиз и Друэ (Jeandelize, Drouet, 1938) нашли, что световое раздражение глаз у лягушки влияет на секреторную деятельность гипофиза; последний выделяет особый меланофорный гормон — интермедин. В отличие от всех прочих цветных лучей, красный световой раздражитель никак не сказывается на выделении этого гормона.
В пользу того, что многие изменения чувствительности глаза к красному цвету осуществляются лишь посредством возбуждения зеленоощущающего аппарата, говорят и данные опытов Селецкой с применением обонятельных раздражителей, о которых мы уже упоминали выше. Наблюдая над изменениями границ цветовых полей зрения, этот автор отметил, что границы полей зрения для красного цвета реагируют на непрямые раздражители лишь в том случае, когда они исследуются маленькими объектами, не выходящими из зоны сетчатки, в которой объект одновременно может возбуждать и зеленоощущающие аппараты сетчатки.
↑ Значение возбуждения зеленореагирующего аппарата для реактивности цветового зрения но отношению к непрямым раздражителям
Согласно нашей гипотезе, изменения в красно- и синеощущающих аппаратах глаза под влиянием разного рода непрямых раздражителей вызываются изменениями возбужденного зеленоощущающего аппарата, а не самими лишь условиями, способными менять возбудимость этого последнего. Если зеленоощущающий аппарат не возбужден (как это бывает при воздействии на глаз крайних красных и крайних фиолетовых лучей спектра), то никаких изменений в возбудимости красно-или синеощущающих аппаратов не наблюдается, хотя бы условия для изменения возбудимости зеленоощущающего аппарата и были налицо (например, глаз подвергался бы действию ан-или катэлектротона).
Мы должны поэтому сформулировать найденную выше закономерность так: изменения возбужденного зеленоощущающего аппарата, связанные с повышением или понижением его возбудимости, оказывают влияние на возбудимость остальных двух цветоощущающих аппаратов нашего зрительного анализатора.
Оставалась все же потребность на опыте, более прямым путем проверить наш вывод относительно того, что возбуждение зеленоощущающего аппарата есть необходимое условие изменяемости цветовой чувствительности глаза под влиянием тех или иных непрямых раздражителей.
Такая прямая экспериментальная проверка нашей гипотезы и была осуществлена в нашей лаборатории Медведевой (1948). Она исследовала, как ведет себя колбочковая чувствительность темноадаптированного глаза по отношению к крайним красным и фиолетовым лучам спектра при пропускании через глаз постоянного тока в обычных условиях монокулярного смотрения на световой раздражитель (при затемненном другом глазе) и при одновременном освещении другого глаза зеленым светом. В последнем случае, очевидно, вместе с возбуждением красно- и синеощущающего аппаратов (вызываемым соответетвующим световым раздражителем исследуемого глаза) в возбуждение приводился в центрах и зеленоощущающий аппарат нашего зрения (благодаря зеленому свету, воздействующему па другой глаз). Следовательно, непрямые раздражители действовали на субъекта, когда зеленоощущающий аппарат был возбужден, и под влиянием этих непрямых раздражителей мог претерпевать те или иные изменения в своей возбудимости. Последнее же, согласно нашим выводам, изложенным выше, должно было уже сказаться и на возбудимости красно- и синеощущающего аппаратов. То обстоятельство, что зеленое раздражение действует на один глаз, в то время как измерения порога производятся другим глазом, не должно менять дела по существу, если допускать, что процессы взаимодействия цветоощущающих аппаратов протекают в нашем зрительном анализаторе более центрально, чем в самой сетчатке.
Опыты, проведенные на четырех лицах, согласно показали, что колбочковая чувствительность к крайнему красному цвету (700 m?) и к фиолетовому (425 m?) при воздействии непрямых раздражителей не остается уже неизменной, если во время определения порога исследуемым глазом другой глаз подвергается освещению зеленым светом. Так, при воздействии анэлектротона, чувствительность к крайнему красному снижается, при воздействии катэлектротона повышается. В случае применения звука в качестве непрямого раздражителя чувствительность к фиолетовому при освещении другого глаза зеленым повышается, чувствительность же к крайнему красному падает. Это наблюденное в опытах направление изменения чувствительности — до того «безразличных» точек — хорошо согласуется с высказанным нами предположением об антагонистической связи зеленоощущающего аппарата с красноощущающим и синэргической связи зеленоощущающего с синеощущающим. Рис. 62
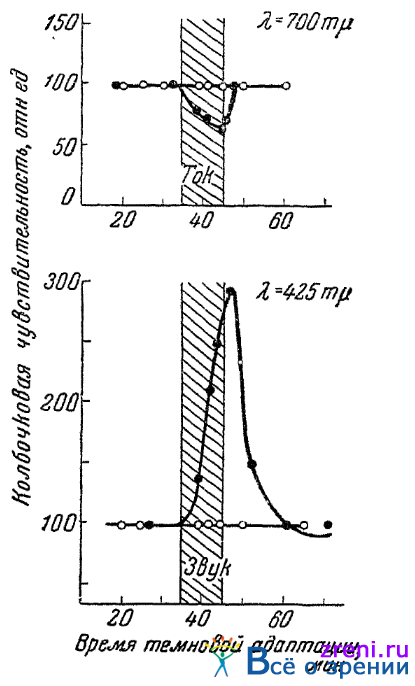
Рис. 62. Влияние освещения другого глаза зеленым светом на реактивность колбочковой чувствительности исследуемого глаза к 425 и 700 m? в ответ на непрямое раздражение eгo (по Медведевой)
иллюстрирует сказанное, воспроизводя результаты опытов Медведевой, в которых испытывалась чувствительность к красному цвету (700 m?) при воздействии на глаз анодом постоянного тока и чувствительность к фиолетовому (425 m?) при звуке.
Следует здесь заметить, что нами позже ставились и контрольные опыты (неопубликованные), в которых второй глаз освещался не зеленым, а крайним красным светом приблизительно того же уровня яркости. При таком добавочном возбуждении не зелено-, а красноощущающего аппарата никаких сдвигов чувствительности по отношению к 700 m? под влиянием анэлектротона нами не наблюдалось. Таким образом, решающее значение возбужденности именно зеленоощущающего аппарата в реакциях цветовой чувствительности на непрямые раздражители можно считать доказанным.
↑ Вероятное место взаимодействия цветореагирующих аппаратов
В опытах Медведевой возбуждение зеленоощущающего аппарата достигалось путем освещения зеленым светом второго глаза. Очевидно, что возникающее здесь возбуждение зеленоощущающего аппарата локализовалось в нервной системе, где-то выше, центральнее сетчатки исследуемого глаза. Это добавочное возбуждение зеленоощущающего аппарата сказывалось на реактивности «безразличных» точек в спектре и проявляло свои антагонистические и содействующие влияния по отношению к аппаратам красно- и синеощущающим. Таким образом, бесспорно, что взаимодействие цветоощущающих аппаратов происходит не в сетчатке, а выше.
Позволительно поставить дальнейший вопрос, где же выше сетчатки — в коре или в подкорке — по преимуществу протекают эти физиологические процессы взаимодействия цветоощущающих аппаратов? Попытку подойти к ответу на этот вопрос мы находим в работе Шварц (1948). Она испытывала действие на цветовую чувствительность глаза разного рода фармакологических веществ. Чувствительность испытывалась (в условиях темповой адаптации) монохроматическим излучениям с длинами волн 435, 460, 520, 570, 625 и 700 m?. Фармакологические же вещества были взяты с тем расчетом, чтобы одни были возбудителями, а другие угнетателями и одни действовали преимущественно на кору, а другие преимущественно на подкорку. Применялись кордиамин (как стимулятор подкорки), веронал (как депрессор подкорки), кофеин (как стимулятор коры) п хлоралгидрат (как депрессор коры). Результаты опытов согласно показали, что медикаменты, действующие преимущественно на кору (кофеин, хлоралгидрат) влияют на чувствительность ко всем лучам спектра одинаково, т. е. или повышают чувствительность ко всем лучам спектра (кофеин), или же ко всему спектру понижают чувствительность (хлоралгидрат). В отличие от этого, вещества, влияющие преимущественно на подкорку (кордиамин, веронал), всегда на цветовую чувствительность к длинноволновым и коротковолновым лучам спектра действуют противоположным образом. Так, кордиамин повышает чувствительность к лучам зелено-синим и понижает ее по отношению к оранжево-красным лучам. Веронал понижает чувствительность к зелено-синим лучам и повышает чувствительность к оранжево-красным. Таким образом, те противоположные изменения чувствительности к зелено-синему и оранжево-красному цветам, которые мы наблюдали в результате воздействия на субъекта ряда непрямых раздражителей и которые мы объясняли как проявление определенного рода взаимодействий, существующих между различными цветоощущающими аппаратами нашего зрительного анализатора, наблюдаются нами при медикаментозном воздействии на подкорковую область нашего мозга. Если под влиянием примененного медикамента возбудимость зелено-ощущающего аппарата в подкорке повышается, то цветовая чувствительность по отношению к оранжево-красным лучам при этих условиях обязательно понижается. Если же чувствительность зеленоощущающего аппарата в подкорке падает, чувствительность к оранжево-красной части спектра обязательно повышается. Одновременного повышения или понижения чувствительности и к коротковолновым и к длинноволновым лучам спектра при медикаментозном воздействии на подкорку никогда не наблюдалось. Эти, установленные в опытах Шварц, факты позволяют считать вероятным, что физиологические механизмы, обусловливающие антагонистическую связь зеленоощущающего аппарата с красноощущающим лежат, по-видимому, в подкорке. Рис. 63
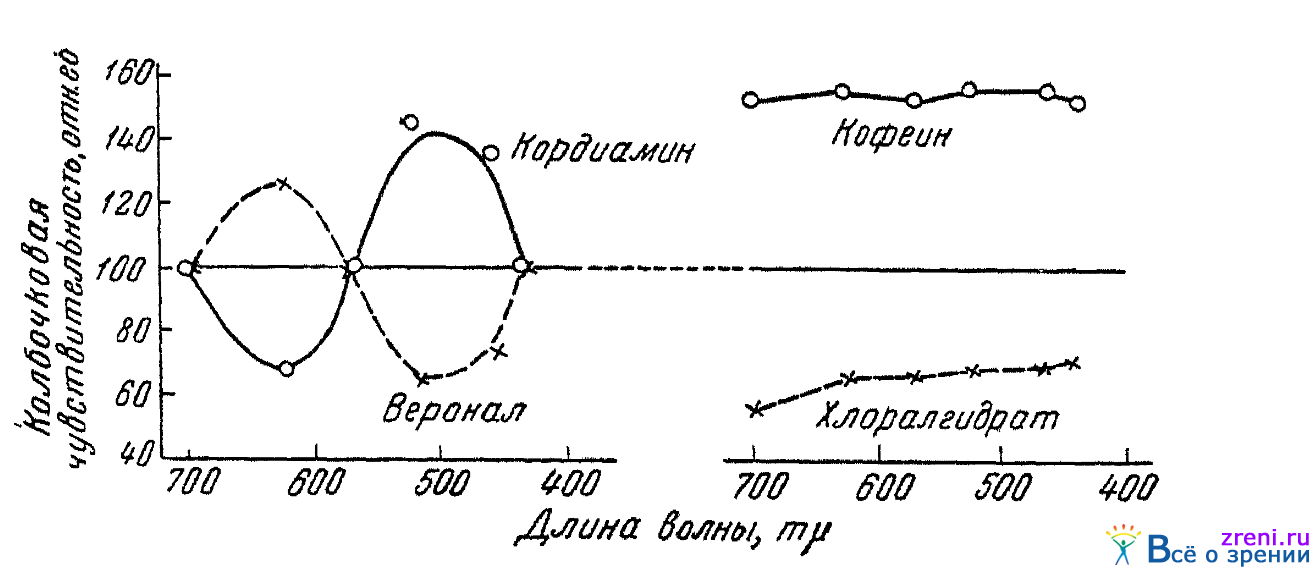
Рис. 63. Изменение колбочковой чувствительности глаза к спектральным лучам в зависимости от различных медикаментозных воздействий (по Шварц)
воспроизводит данные одного из испытуемых, типичные для всех опытов. По абсциссе отложены длины волны спектральных лучей, по ординате — относительное значение цветовой чувствительности на 40-й минуте после приема того или иного медикамента. За 100 принят установившийся уровень чувствительности, имевшийся до приема медикамента.
↑ Действие цветного света на состояние глаза и организм вообще
Описанными выше опытами с применением непрямых раздражителей была вскрыта различная природа зелено - и красноощущающих аппаратов глаза. Это различие в их физиологической природе носит, как мы видели, характер известной противоположности в том смысле, что ионные сдвиги, благоприятные для чувствительности одного цветоощущающего аппарата, оказываются неблагоприятными для чувствительности другого. Зеленоощущающий аппарат делается более возбудимым в условиях симпатикотонии; аппарат же красноощущающий становится чувствительнее в условиях ваготонии. Симпатикотропность (в этом смысле) первого и ваготропность второго требуют, конечно, объяснения с общебиологической точки зрения, с точки зрения биологической целесообразности подобного рода связи цветоощущающих аппаратов с различными отделами вегетативной нервной системы. Пока, к сожалению, мы такого объяснения дать еще не можем.
Здесь, в заключительной части нашей книги, мы хотели бы указать еще на то, что цветовое зрение не только расширяет наше познание окружающего объективного мира, но и позволяет входить в более тесную связь с изменяющимися факторами окружающей среды, различным образом перестраивая физиологическое состояние организма в зависимости от этих факторов. Мы имеем в виду вскрытые в последнее время работами нашей лаборатории факты изменений в состоянии органа зрения и организма вообще, происходящих в зависимости от цветности света, действующего на наш зрительный анализатор. Описанная выше противоположность зелено- и красноощущающего аппаратов весьма резко сказывается на характере реакций, вызываемых в организме зеленым и красным цветом.
Зарецкая (1941), изучая внутриглазное давление в различных условиях освещения, нашла, что освещение одного глаза зеленым светом вызывает понижение внутриглазного давления в другом глазе. Освещение же глаза светом красным (той же яркости), наоборот, повышает внутриглазное давление в другом глазе (исследуемом). Насколько различны возникавшие здесь реакции внутриглазного давления, можно видеть из рис. 64,

Рис. 64. Влияние цветового раздражения на внутриглазное давление (по Зарецкой)
воспроизводящего типичные результаты опытов Зарецкой.
Позже, Зарецкой же (1948), было найдено, что физиологические границы слепого пятна в зависимости от адаптации глаз к зеленому или к красному свету меняются в обратных направлениях. При адаптации к зеленому слепое пятно уменьшается, при адаптации к красному, напротив, становится больше. Факт этот был подтвержден затем в работе Рославцева (1949). Ий было, кроме того, показано, что адаптация глаз к зеленому и красному свету обратным образом сказывается и на ширине ангиоскотом, т, е. тех мелких слепых мест в поле зрения, которые обусловливаются тенями кровеносных сосудов, находящихся перед светочувствительным слоем сетчатки. В красном свете ангиоскотомы расширяются, в зеленом суживаются. Семеновская, Зарецкая и Гуртовой установили, далее, что электрическая чувствительность глаза при освещении глаз зеленым светом бывает заметно более высокой, чем при раздражении глаз красным светом той же яркости. В условиях красного освещения электрическая чувствительность глаза в ряде случаев оказывается даже более низкой, чем в условиях темноты.
Красный и зеленый свет различно действуют и на некоторые «незрительные» функции организма. Так, укажем, например, что, по данным опытов Шварц (1949), адаптация глаз к зеленому свету повышает слуховую чувствительность человека, пребывание же в условиях красного освещения (при той же яркости) слуховую чувствительность снижает. В опытах Шеваревой (1950) было показано, что зеленый и красный свет по-разному сказываются на мускульно-двигательной работоспособности пальца руки. Устанавливаемая эргографическим методом работоспособность эта при зеленом освещении у всех испытуемых оказалась больше, чем при освещении красным светом той же яркости.
↑ Цветное освещение как фактор среды, меняющий физиологический «фон» организма
Световые раздражения являются естественными адекватными раздражителями нашего зрительного анализатора. Посредством периферических и центральных (подкорковых и корковых) частей его мы различаем цвета окружающего нас мира и в соответствии с этим реагируем на изменения внешней среды теми или иными переменами как в самом нашем зрительном приборе, так и в поведении нашего организма вообще.
Внутриглазное давление, равно как ангиоскотомы и физиологические границы слепого пятна, безусловно зависят от сосудодвигательных реакций, изменений в проницаемости сосудистых стенок, т. е. от процессов, регулируемых вегетативной нервной системой. Состояние вегетативной нервной системы, несомненно, сказывается и на мышечной работоспособности, а также и на слуховой чувствительности. Различное и часто обратное действие зеленого и красного света можно, поэтому, отнести за счет различного действия этих цветовых раздражителей именно на вегетативную нервную систему. Такой вывод, как нетрудно видеть, хорошо согласуется с теми характеристиками, которые по современным опытным данным следует давать зелено- и красноощущающему аппаратам глаза.
Действие же вегетативной нервной системы, как мы видели выше, часто бывает подобно определенным ионным сдвигам в ткани. Мы приводили уже взгляды (например, Цондека), по которым ионные изменения, главным образом изменения в соотношении ионов калия и кальция, необходимы для реализации действия симпатического или блуждающего нервов. Определенные же ионные сдвиги бывают связаны и с определенными изменениями в содержании ацетилхолина в ткани. Если всю эту цепь зависимости иметь в виду, то можно полагать, что действие зеленого и красного света на глаз, вызывая через центральную нервную систему (кору и подкорку) определенные вегетативные сдвиги, бывает связано с определенными изменениями ионного состояния ткани и изменениями в ее ацетилхолиновом хозяйстве. В частности, зеленый раздражитель, как симпатикотропный в указанном смысле, должен изменять ионное отношение ткани реагирующего органа в направлении относительного увеличения ионов кальция (и соответственно —уменьшения содержания ацетилхолина). Красный же раздражитель, как ваготропный, должен производить в ионном показателе перемену в сторону относительного увеличения ионов калия (и соответственно — увеличения ацетилхолина).
Подтверждение правильности подобных умозаключений можно видеть в опытных данных, полученных недавно в нашей лаборатории Буниным (1949) и Рославцевым (1949). Бунин на биологических объектах (сердце лягушки и мышце пиявки) прямым образом показал, что при одинаковой освещенности глаз в сетчатках лягушачьих глаз больше ацетилхолина при адаптации глаз к красному свету и меньше при адаптации к зеленому. На рис. 65
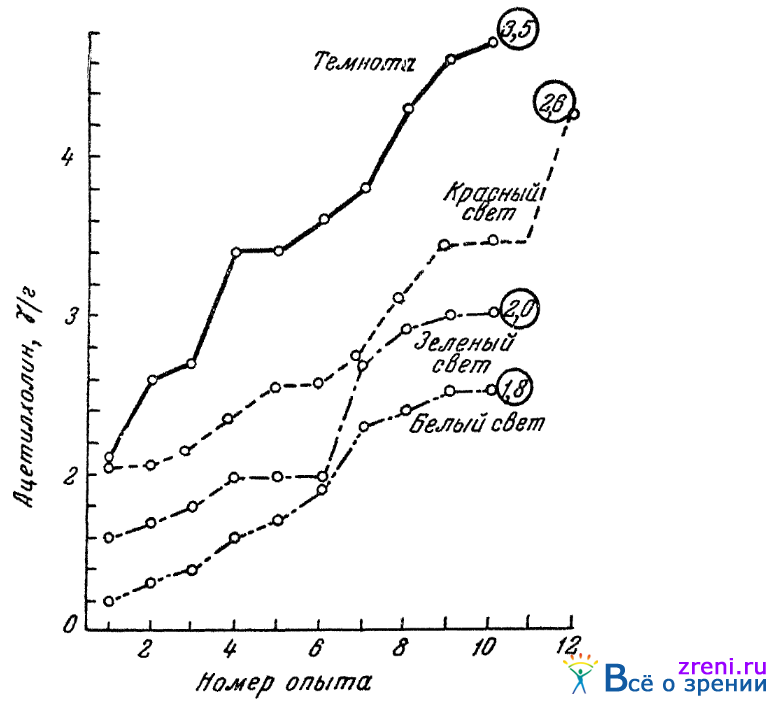
Рис. 65. Содержание ацетилхолина в сетчатке в зависимости от условий освещения (по Бунину). В кружках указаны средние значения (медианы); красный и зеленый свет подравнены по яркости
показаны результаты опытов Бунина по влиянию цветности освещения на количество ацетилхолина в сетчатках. Значения количества ацетилхолина (в «гаммах», т. е. в 0,001 мг на грамм ткани — ?/г) для каждых условий освещения расположены в порядке от минимальных до максимальных, наблюдавшихся в опытах.
Рославцев же наблюдал, что действие цветности зеленого и красного освещения на границы слепого пятна и на ширину ангиоскотом не сказывается в глазах, подвергшихся атропинизации. Этот факт говорит за то, что в реакциях ангиоскотом п слепого пятна на цветное освещение участвует ацетилхолиновое звено.
Вспомним теперь, что по нашему анализу всех экспериментальных данных, добытых нашими сотрудниками и нами, следовало, что сдвиги цветовой чувствительности под влиянием разного рода непрямых раздражителей объясняются тем, что в конечном результате всех нервных изменений исходное ионное состояние среды реагирующей ткани, характеризуемое отношением [К] + [Na]/[Ca] + [Mg] изменяется. При этом, в зависимости от того, произойдет ли этот сдвиг в направлении к оптимальной для возбудимости величине ионного отношения, или же в обратную сторону, мы будем наблюдать или повышение, или понижение цветовой чувствительности.
Не трудно понять, что один и тот же агент, действующий всегда в одном и том же направлении (например, увеличивающий в конечном итоге относительную концентрацию ионов кальция), может влиять на чувствительность и положительно и отрицательно, в зависимости от величины наличного ионного отношения в ткани. Если это отношение по своей величине больше оптимального, то, очевидно, наш агент будет приближать его к этой оптимальной величине и тем самым повышать чувствительность исследуемого субстрата. Если же величина ионного отношения ткани меньше оптимальной, тот же агент повлияет на чувствительность ухудшающим образом. Если мы признали, что цветной свет может изменять величину ионного отношения в нашем зрительном анализаторе, мы должны допустить, что адаптация глаз к тому или другому цветному раздражителю может менять направление реакций колбочковой чувствительности глаза в ответ на различные непрямые раздражители.
Этот вывод также нашел себе экспериментальное подтверждение. Опыты показали, что в зависимости от того, адаптирован ли глаз к зеленому или красному свету, реакция колбочковой чувствительности в ответ на одно и то же непрямое раздражение может быть противоположной. Мы (1948) испытывали изменение видности цветной полосы, находящейся на зеленом или красном фоне, к цвету которого и был адаптирован глаз, и смогли установить, что видность этой полоски при воздействии непрямых раздражителей (таких, как анэлектротон и звук) меняется в обратном направлении при адаптации глаза к зеленому или же к красному.
В этих наших опытах, правда, не было исключено возможное влияние контраста со стороны цветного фона, который мог меняться под влиянием непрямого раздражителя. Особенно показательны в рассматриваемой связи, поэтому, здесь, как нам кажется, результаты более поздних опытов Гуртового, в которых он изучал влияние цветового раздражения второго (не испытываемого) глаза; испытуемый же глаз был все время темноадаптирован. Ему удалось наблюдать, что и при подобной методике, когда возможное влияние контраста с фоном исключено и цветовая адаптация действует уже не на периферию, а на центральные части зрительного анализатора, ее влияние все же может изменять реакции колбочковой чувствительности глаза на обратные. Так при адаптации одного глаза к красному непрямое звуковое раздражение вызывает сдвиг чувствительности колбочкового аппарата другого глаза в сторону ее повышения (как это происходит обычно и в темноте). При адаптации же одного глаза к зеленому цвету, наступающее при воздействии того же звука, изменение чувствительности другого глаза идет уже в противоположном направлении мы наблюдаем снижение чувствительности. На рис. 66
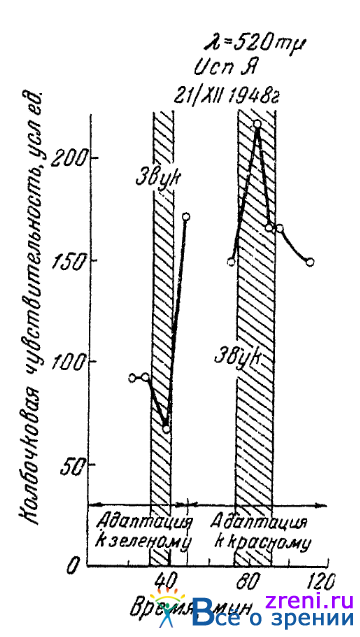
Рис. 66. Влияние цветовой адаптации одного глаза на направление изменений колбочковой чувствительности другого в ответ на слуховое раздражение (по Гуртовому)
приводятся результаты одного из подобных опытов. Результаты опытов опубликованы в статье Гуртового, написанной совместно с нами (1951).
Можно, таким образом, сказать, что цветовые воздействия на глаз влекут за собой перестройку нервной системы организма и тем самым меняют «физиологический фон», на который падают те или иные применяемые нами воздействия. Это с несомненностью показано и недавними опытами Семеновской и Кондорской (1950), которые изучали электрическую чувствительность глаза и его лабильность (функциональную подвижность) и установили, что оба эти показателя возбудимости нервного прибора глаза меняются по-разному в условиях адаптации к красному или к зеленому свету.
В какой мере исходное состояние, физиологический «фон», может определять реактивность нашего органа зрения, показывают, наконец, еще и недавние эксперименты Рославцева (1950). Им было установлено, что в обычных условиях освещения глаза зеленым светом ангиоскотомы суживаются, от освещения же светом красным — расширяются. Эти обычные, «нормальные» реакции на цветные раздражители, однако, извращаются, если человек потерял более или менее значительное количество крови. Специальные опыты цитируемо го автора показали, что у доноров, после гемоэксфузий, равно как и у женщин в менструальный период, ширина ангиоскотом суживается и в ответ на красный свет, что в нормальных условиях происходит лишь в ответ на цвет зеленый. В зависимости от потери организмом некоторого количества крови реакции глаза на цветные раздражители, таким образом, «извращаются».
В самое последнее время исследования Рославцева и Семеновской, выполненные в нашей лаборатории и направленные на изучение функциональных особенностей зрительного анализатора глаукоматозных больных, установили наличие подобных «извращенных» реакций на красный свет у этих больных. В ответ на освещение их глаз красным светом, размеры слепого пятна и ангиоскотом, а также и функциональная подвижность органа зрения у больных глаукомой изменяются так же, как они изменяются у здоровых в ответ на освещение их глаз зеленым. В связи с тем, что сказано выше о «симпатикотропности» зеленого раздражителя и «парасимпатикотропности» красного, описанное извращение реакций на красный свет можно рассматривать как показатель известной симпатикотонии при глаукоме.
Конечно все описанные выше процессы, лежащие в основе изменений цветовой чувствительности глаза, находятся в зависимости и от высшего регулятора жизни всего организма — коры головного мозга.
Вскрыть ближе эти зависимости и их конкретные проявления в изменениях цветовой чувствительности глаза составляет одну из очередных задач дальнейших исследований в области психофизиологии цветового зрения.
---
Статья из книги: Цветовое зрение | Кравков С. В.

Комментариев 0