Зрение человека


Содержание:
Описание
↑ Введение
Зрительная система человека достигла высочайшего совершенства. Ученые, работающие над созданием электронных или химических систем со сравнимыми характеристиками, могут лишь восхищаться ее чувствительностью, компактностью, долговечностью, высокой степенью воспроизводимости и изящной приспособляемостью к потребностям человеческого организма. Справедливости ради следует, конечно, отметить, что попытки создания соответствующих искусственных систем были начаты менее столетия назад, тогда как зрительная система человека формировалась в течение миллионов лет. Она возникла из некоего «космического» набора элементов — отбираемых, отбираемых и отбираемых до тех пор, пока не выпадала удачная комбинация. Мало кто сомневается, что эволюция человека носила «слепой», вероятностный характер, и проследить ее шаг за шагом совершенно невозможно. Издержки эволюции давно канули в Лету, не оставив следа.
Зрение занимает почти уникальное место в схеме эволюции. Можно допустить, например, что в дальнейшем эволюционное развитие приведет к увеличению объема мозга, усложнению нервной системы или к различным усовершенствованиям существующих функций. Однако невозможно представить, что чувствительность зрительного процесса заметно возрастет. Зрительный процесс представляет собой абсолютную конечную веху в цепи эволюции. Если принять во внимание, что в зрительном процессе «считается» каждый поглощенный фотон, то дальнейшее увеличение чувствительности маловероятно, если не увеличится поглощение. Законы квантовой физики устанавливают жесткий предел, к которому наша зрительная система приблизилась почти вплотную.
Мы сделали оговорку, что зрение занимает почти уникальное место, поскольку согласно определенным данным, некоторые другие процессы восприятия также достигли абсолютного предела в своей эволюции. Способность ряда насекомых (например, моли) «детектировать» отдельные молекулы служит свидетельством того, что и обоняние в иных случаях достигло квантового предела. Аналогично наш слух в пределе ограничен тепловым шумом окружающей среды.
Высокая чувствительность зрительного процесса не является привилегией только человека. Имеются явные свидетельства того, что менее совершенные виды животных и ночные птицы достигли здесь подобного уровня. По-видимому, рыбы, обитающие в темных глубинах океана, также должны использовать до предела ту скудную информацию, которая проникает к ним со случайными лучами света. Наконец, мы можем указать на фотосинтез как свидетельство того, что различные формы растительной жизни издавна научились использовать почти каждый падающий фотон, во всяком случае в пределах определенной спектральной области.
Основная цель этой главы — продемонстрировать высокую квантовую эффективность человеческого глаза в широкой области интенсивностей света. Для того чтобы выразить исходные данные о зрении человека через плотность фотонов на единицу площади сетчатки, необходимо знать» оптические параметры человеческого глаза. Их мы рассмотрим в следующем разделе.
↑ Оптические параметры
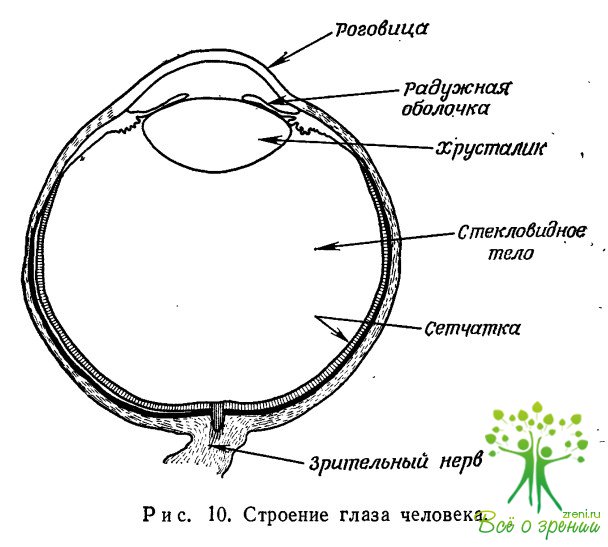
На рис. 10 показано строение человеческого глаза.
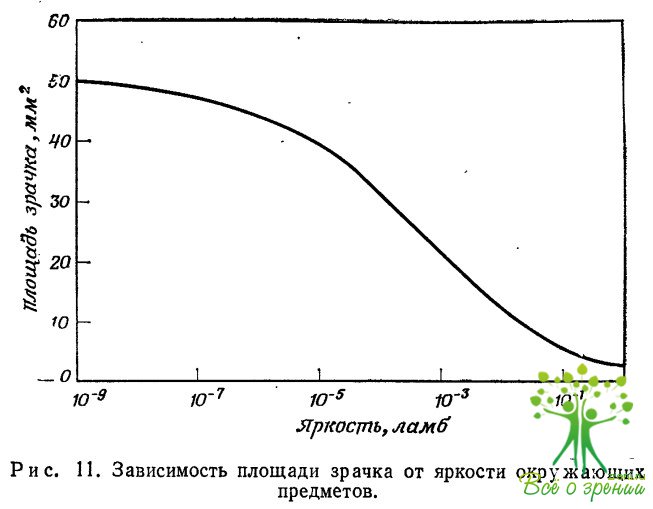
Отверстие зрачка хрусталика меняется от 2 мм при большой освещенности до примерно 8 мм вблизи порога зрительного восприятия. Эти изменения происходят за время порядка десятых долей секунды. Фокусное расстояние хрусталика равно 16 мм. Это означает, что относительное отверстие оптической системы меняется от 1:2 при малых освещенностях до 1:8 при высоких освещенностях. Примерная зависимость площади зрачка от уровня освещенности показана на рис. 11.
Светочувствительный слой, называемый сетчаткой, состоит из дискретных светочувствительных клеток, палочек и колбочек, отстоящих друг от друга примерно на 2 мкм. Вся сетчатка — ее площадь близка к 10 см2 — содержит 108 таких элементов. Колбочки, расположенные преимущественно в области центральной ямки, имеющей угловые размеры около 1°, работают при средних и высоких освещенностях и передают цветовые ощущения. Палочки, занимающие большую часть площади сетчатки, действуют вплоть до самых малых освещенностей и не обладают цветовой чувствительностью. Колбочки определяют предел разрешающей способности при высоких уровнях освещенности, который составляет 1—2', что близко к размерам дифракционного диска, соответствующего диаметру зрачка хрусталика, равному 2 мм. Исследование работы глаза и анатомическое изучение его строения показывают, что по мере удаления от центра сетчатки палочки объединяются во все большие и большие группы вплоть до нескольких тысяч элементов каждая. Свет, попадающий на сетчатку, проходит через слой нервных волокон, расходящихся от зрительного нерва к клеткам сетчатки.
Пространство между хрусталиком и сетчаткой заполнено водянистой средой, так называемым стекловидным телом, показатель преломления которого равен 1,5. Согласно различным оценкам, до сетчатки доходит лишь половина падающего на глаз света. Остальной свет отражается или поглощается.
Физическое время накопления фотонов глазом лежит в пределах от 0,1 до 0,2 с и, вероятно, ближе к последней цифре. Физическое время накопления эквивалентно времени экспозиции в фотографии. При переходе от высоких освещенностей к порогу зрительного восприятия время накопления возрастает не более чем в два раза. «Работе» глаза подчиняется закону взаимозаменяемости: при времени экспозиции меньше 0,1—0,2 с его реакция зависит только от произведения интенсивности света на время воздействия последнего.
↑ Качественные показателиВ течение последних ста лет происходило непрерывное накопление данных, касающихся человеческого зрения. Блэкуэлл опубликовал результаты последних и наиболее полных измерений способности глаза различать отдельные пятна различного размера и контраста при изменении освещенности в широких пределах. На рис. 12 приведены данные, полученные Блзкузллом в интервалах освещенностей 10-9 — 10-1 ламб, контраста 1 — 100% и углового разрешения 3—100'. Мы опустили данные, относящиеся к контрастам, меньшим 1%, и угловому разрешению ниже 3поскольку очевидно, что характеристики глаза в этой области ограничены не шумовыми факторами, а другими причинами; последние устанавливают абсолютный предел различимости контраста 0,5%, а углового разрешения 1—2'. Геометрический предел разрешающей способности определяется конечным размером палочек и колбочек.На рис. 13 представлены аналогичные данные, по-лученные ранее Коннером и Ганунгом (1935), а также Коббом и Моссом (1928). Как видно, данные, приведенные на рис. 12 и 13, в общих чертах согласуются друг с другом. Однако существенное различие заключается в том, что, согласно данным Блэкуэлла, характеристики не улучшаются при изменении яркости в пределах 10-2-10-1 ламб, тогда как, по данным Кобба и Мосса, такое улучшение имеет место.На рисунках линии, идущие под углом 45°, представляют собой характеристики, которые следовало бы ожидать в том случае, если бы свойства системы были ограничены шумом, согласно соотношению (1.2). На рис. 13 экспериментальные точки довольно хорошо укладываются на прямые, соответствующие шумовым ограничениям и идущие под углом 45°. На рис. 12 экспериментальные кривые имеют вид изогнутых линий, которые касаются указанных прямых лишь на ограниченных участках. Такие отклонения можно, по-видимому, объяснить влиянием ограничений, не связанных с фотонным шумом.? Квантовая эффективность зрения человека
Для оценки квантовой эффективности глаза данные, приведенные на рис. 12 и 13, следует выразить через число фотонов, падающих на 1 см2 сетчатки. Для этого предположим, что время накопления равно 0,2 с, пропускание хрусталика 0,5, а пределы раскрытия зрачка определяются данными Рива, представленными на рис. 11. Проделав такое преобразование, подставляем плотность фотонов в соотношение (1.3), записанное в виде
C2*d2*?*n=k2=25,
где ? — квантовый выход глаза (квантовая эффективность ? 100*?%)- Пороговое отношение сигнал/шум k принимается равным 5.
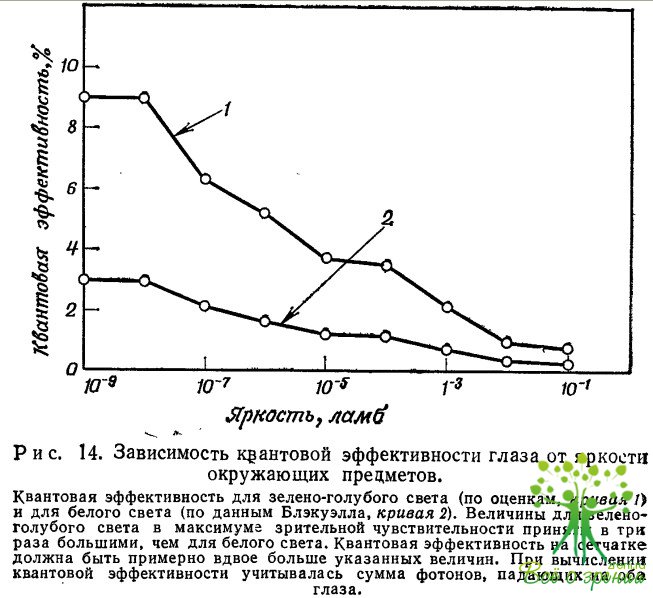
На рис. 14 показана зависимость квантовой эффективности глаза (вычисленной по данным Блэкуэлла) от яркости объектов. В этих результатах более всего поражает относительно малое изменение квантовой эффективности при изменении интенсивности света на 8 порядков. Квантовая эффективность составляет 3% при предельно низких яркостях вблизи абсолютного порога (примерно 10-10 ламб) и медленно убывает до примерно 0,5% при 0,1 ламб.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Конечно, это десятикратное изменение эффективности. Однако следует вспомнить, что в ранних работах для объяснения явления темнотой адаптации в подобных случаях предполагалось 1000- или 10000-кратное изменение квантовой эффективности. (Ниже мы рассмотрим этот вопрос подробнее.) Более того, даже это десятикратное изменение на самом деле может оказаться сильно завышенным. При расчете квантовой эффективности мы предполагали, что время экспозиции и множитель k постоянны, но, по некоторым данным, при низких освещенностях время экспозиции может быть вдвое большим, чем при высоких. Если это так, то квантовая эффективность изменяется лишь в пять раз. Далее, не исключено, что множитель k при низких освещенностях меньше, чем при высоких. Такое изменение k (точнее, k2) может легко привести к появлению еще одного множителя 2, в результате оказывается, что квантовая эффективность изменяемся всего лишь в 2 раза при изменении интенсивности света в 108 раз.
Второй важный момент, который следует отметить, анализируя рис. 14, — это относительно большая величина квантовой эффективности.
Согласно имеющимся в литературе оценкам, чувствительное вещество сетчатки (родопсин) поглощает только 10% падающего света. Если это так, то квантовая эффективность (для белого света) по отношению к поглощенному свету составляет около 60% при низких освещенностях. Таким образом, остается весьма мало возможностей для улучшения самого механизма счета фотонов.
Однако трудно понять, чем обусловлено столь низкое поглощение (лишь 10%) падающего света, которое сформировалось в процессе эволюции. Возможно, что причиной этого послужил ограниченный выбор биологических материалов.
Некоторое уменьшение квантовой эффективности при высоких освещенностях можно приписать специфическим требованиям, которые предъявляются к системе, способной различать цвета. Если, как показывают последние данные, имеется 3 типа колбочек с различными спектральными характеристиками, то площадь, чувствительная к свету с данной длиной волны, при высоких освещенностях уменьшается вдвое.
Значения квантовой эффективности, представленные на рис. 14 нижней кривой, относятся к белому свету. Известно, что зрительная реакция на зеленый свет примерно в три раза выше, чем на то же самое полное число «белых» фотонов, то есть фотонов, распределенных по всему видимому спектру. Использование зеленого света (или зелено-голубого при низких освещенностях) должно было бы привести к увеличению квантовой эффективности втрое, как это показано на рис. 14. В таком случае квантовая эффективность при малых освещенностях составляла бы примерно 10%, и мы должны были бы предположить, что сетчатка поглощает не 10%, а по крайней мере 20% падающего света.
Следует вновь подчеркнуть, что значения квантовых эффективностей, приведенные на рис. 14, зависят от выбора параметров: времени накопления (0,2 с) и порогового отношения сигнал/шум {k = 5). Значения этих параметров определены недостаточно точно, особенно это касается данных Блэкуэлла.
Возможно, соответствующие уточнения приведут к более высоким значениям квантовой эффективности. Например, если предположить, что время накопления составляет 0,1 с, то квантовые эффективности окажутся вдвое больше, чем приведенные на рис. 14. Однако вряд ли следует затрачивать усилия на уточнение этих пара-метров; не лучше ли заняться разработкой усовершенствованной, не зависящей от этих параметров экспериментальной методики измерения квантовой эффективности.
↑ Предпочтительный метод определения квантовой эффективности
В настоящее время существует исключительно простая методика довольно надежного определения квантовой эффективности глаза. Недавно разработанная телевизионная камера с кремниевым усилителем способна передавать изображения при низких уровнях освещенности, когда эти изображения явно ограничены шумами, точнее шумом, обусловленным частью падающих фотонов, которые создают фотоэлектроны на фотокатоде.
Существенно, что такие изображения, ограниченные только шумами, позволяют надежно определять квантовую эффективность фотокатода. Процедура заключается в том, что наблюдатель и телевизионная камера «разглядывают» один и тот же слабо освещенный объект с одинакового расстояния. Диафрагма на оптике камеры устанавливается в соответствии с раскрытием зрачка глаза наблюдателя. Затем наблюдатель сравнивает непосредственно видимый им слабо освещенный объект с изображением на кинескопе телевизионной системы. Если информация одинакова, то квантовая эффективность глаза наблюдателя равна измеренной эффективности фотокатода передающей трубки. Если наблюдатель видит больше или меньше, чем камера, то диафрагму регулируют до тех пор, пока различие не исчезнет, после чего квантовая эффективность глаза наблюдателя вычисляется по отношению диафрагм линз.
Основное достоинство метода сравнения «бок о бок» состоит в том, что он не зависит от времени зрительной экспозиции или выбора соответствующего порогового отношения сигнал/шум. Эти параметры, каковы бы ни были их точные значения, по существу остаются одинаковыми при рассмотрении наблюдателем самого объекта и его изображения на телевизионном экране, следовательно, при сравнении они исключаются. Более того, влияние памяти на эффективное время экспозиции в этих двух случаях, по-видимому, будет одинаковым.
Мы остановились на этом методе, поскольку сейчас он легкодоступен экспериментаторам, искушенным в исследовании зрительного процесса. Различные устройства, пригодные для сравнения, использовались как автором данной книги, так и другими исследователями для предварительных оценок квантовой эффективности при малых освещенностях. В одном из экспериментов использовалось устройство для сканирования движущимся световым пятном (рис. 15); Дж. Е. Рюди применил суперортикон усилителем изображения, а Т. Д. Рейнольде воспользовался многоступенчатым усилителем изображения. Все эти устройства давали изображения, ограниченные фотонным шумом, и всех случаях по предварительным оценкам квантовая эффективность составляла около 10% для низких уровней освещенности.
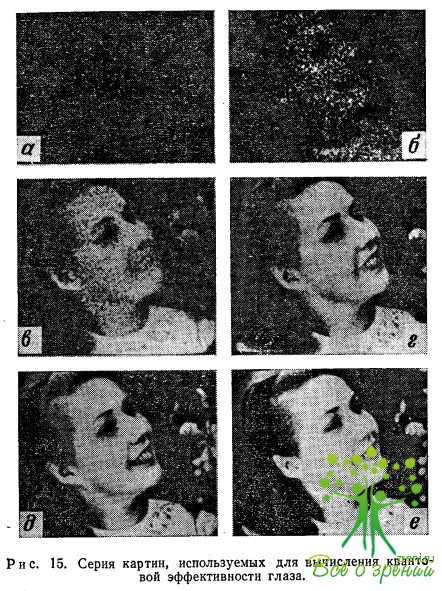

Серия изображений, представленных на рис. 15, показывает, какое максимальное количество информации может быть передано , различным заданным числом фотонов. Каждый фотон регистрируется как дискретная видимая точка. Информация, которую мы получаем, ограничена только статистическими флуктуациями, неизбежно проявляющимися при регистрации потока фотонов. В таблице дано полное число фотонов N. которое содержалось бы в изображении, если бы все оно равномерно освещалось с интенсивностью, соответствующей его наиболее ярким участкам.
Указанные в таблице яркости рассчитаны в предположении, что глаз использует один из каждых десяти падающих фотонов. При расчете учитывались и другие параметры: время накопления — 0,2 с, диаметр зрачка — примерно 6 мм. Иными словами, если объект заменить белым листом с указанной яркостью, вычислить число фотонов, попадающих в глаз за 0,2 с, и поделить это число на 10, то в результате мы получим число фотонов N. соответствующее данной величине яркости. Следовательно, приведенная серия изображений показывает, какое максимальное количество информации может в действительности воспринять наблюдатель при указанных яркостях, если квантовая эффективность его зрительного процесса равна 10%, а расстояние от объекта до наблюдателя составляет 120 см.
↑ Сравнение различных оценок квантовой эффективности
Уже более столетия назад стало известно, что на абсолютном пороге видимости едва различима вспышка от маленького источника, при которой в глаз попадает примерно 100 фотонов. Так был установлен нижний предел квантовой эффективности, равный примерно 1%. Затем несколькими исследовательскими группами была выполнена серия экспериментов с целью выяснить, сколько из этих 100 фотонов на самом деле использует глаз. Если, например, глаз использовал бы все 100 фотонов, то переход от невидения к видению был бы достаточно резким и совершался бы при увеличении потока фотонов до 100. Если бы глаз использовал только несколько фотонов, то переход был бы размыт вследствие хаотического характера эмиссии фотонов. Таким образом, резкость перехода может служить мерой числа используемых фотонов и, следовательно, квантовой эффективности глаза
Идея такого эксперимента была не лишена определенной простоты и элегантности. К сожалению, в результате проведения подобных экспериментов оказалось, что число фотонов, которое используется глазом при пороговом восприятии, меняется в широком интервале от 2 до 50. Таким образом, вопрос о квантовой эффективности оставался открытым. Разброс полученных результатов, по-видимому, не удивит инженера— специалиста в области электроники или физики. Измерения были выполнены вблизи абсолютного порога видимости, когда к шуму фотонного потока легко примешивается шум от посторонних источников внутри самого глаза. Например, если произвести аналогичные измерения с фотоумножителем, то подобный разброс был бы обусловлен влиянием шума, связанного с термоэлектронной эмиссией с фотокатода или со случайным электрическим пробоем, возникающим между электродами. Все это справедливо для измерений вблизи абсолютного порога. Если же производить измерения отношения сигнал/шум при освещенности значительно выше пороговой, когда фотонный шум превышает шум, связанный с посторонними источниками, такая процедура дает надежное значение квантовой эффективности. Именно поэтому результаты измерений зрительной квантовой эффективности, выполненные при освещенностях, значительно превышающих абсолютный зрительный порог, более надежны.
Р. Кларк Джонс провел анализ тех же данных, на основании которых была получена кривая квантовой эффективности, представленная на рис. 14. Определенные им эффективности, в общем, примерно в десять раз меньше, чем показано на рис. 14; при расчетах он исходил из более короткого времени накопления (0,1 с) и гораздо меньшей величины k (1,2). Джонс считает, что поскольку наблюдатель должен лишь выбрать одно из восьми возможных положений тестового объекта, то такая величина k обеспечивает надежность 50%. В количественном отношении это утверждение, разумеется, правильно.
Основной вопрос состоит в том, в действительности ли наблюдатели делают свои заключения о видимом именно таким образом. Если мы обратимся к рис. 4, а, то обнаружим, что k = 1,2 означает, что наблюдатель может заметить, с какой из восьми возможных площадок оператор удалил один или два фотона. Простое рассмотрение рис. 4, а показывает, что это невозможно. Подобные вопросы подчеркивают необходимость разработки такого метода измерений, который позволил бы избежать неоднозначностей, связанных с выбором правильных значений k или времени накопления. Описанный выше способ сравнения «бок о бок» человеческого глаза и электронного устройства, ограниченного фотонным шумом, представляет собой именно такую процедуру и заслуживает самого широкого применения.
В своих ранних оценках зрительной квантовой эффективности Де Врис также исходил из величины k = 1, и его результаты были значительно ниже значений, приведенных на рис. 14. Де Врис, однако, одним из первых указал на то, что наблюдаемая разрешающая способность глаза и его контрастная чувствительность могут быть объяснены фотонным шумом. Кроме того, он, как и автор этой книги, обратил внимание на то, что флуктуирующий, зернистый характер изображений, полученных при слабой освещенности, служит свидетельством дискретности света.
Барлоу в значительной степени избежал неоднозначности в выборе k, проводя измерения с помощью двух расположенных рядом тестовых световых пятен. ЕГО цель заключалась в том, чтобы установить, какое пятно ярче, причем относительные интенсивности пятен менялись. Как показал статистический анализ результатов, проведенный в предположении, что способность различать яркости- ограничена фотонным шумом, значения квантовой эффективности глаза лежат в интервале 5—10% при изменении яркости вплоть до величины, в 100 раз превышающей абсолютный зрительный порог. Барлоу ссылается на работы Баумгардта и Хехта, которые из анализа кривой вероятности обнаружения вблизи абсолютного порога получили квантовую эффективность, близкую к 7%.
Резюмируя, можно сказать, что большинство исследователей считают, что квантовая эффективность человеческого глаза лежит в пределах 5—10% при изменении интенсивности света от абсолютного порога до величины, в 100 раз его превышающей. Эта эффективность определена для длин волн вблизи максимума кривой чувствительности глаза (зелено-голубая область) и относится к свету, падающему на роговицу глаза. Если предположить, что до сетчатки доходит только половина этого света, то эффективность на сетчатке составит 10—20%. Поскольку, согласно имеющимся оценкам, доля света, поглощаемого сетчаткой, также лежит в этих пределах, то эффективность глаза, отнесенная к поглощенному свету, близка к 100%. Иначе говоря, глаз способен считать каждый поглощенный фотон.
Данные, приведенные на рис. 14, указывают еще на одно в высшей степени существенное обстоятельство: в области от абсолютного порога чувствительности до 0,1 ламб, то есть при изменении интенсивности, в 10 раз, квантовая эффективность уменьшается не более чем в 10 раз. В дальнейшем может оказаться, что этот фактор не превышает 2—3. Таким образом, глаз поддерживает высокий уровень квантовой эффективности при изменении интенсивности света в 108 раз! Мы используем это заключение при интерпретации явления темновой адаптации и появления зрительного шума.
↑ Темновая адаптация
Одним из наиболее известных и вместе с тем удивительных аспектов зрительного процесса является темновая адаптация. Человек, входящий в темный зрительный зал с залитой светом улицы города, оказывается буквально слепым в течение нескольких секунд или даже минут. Затем постепенно он начинает видеть все больше и больше и через полчаса полностью привыкает к темноте. Теперь он может видеть объекты более чем в тысячу раз темнее тех, которые он едва различал в первый момент.
Эти факты свидетельствуют о том, что в процессе темновой адаптации чувствительность глаза увеличивается более чем в тысячу раз. Подобные наблюдения направили исследователей на поиски механизма или химической модели, которые объяснили бы столь сильные изменения чувствительности. Например, Хехт уделил особое внимание явлению обратимого выцветания чувствительного материала сетчатки, так называемого зрительного пурпура. Он утверждал, что при низких освещенностях зрительный пурпур совершенно не затрагивается и таким образом обладает максимальным поглощением. С увеличением освещенности он все более обесцвечивается и соответственно поглощает все меньше и меньше падающего света. Считалось, что длительное время темновой адаптации обусловлено большой продолжительностью процесса восстановления высокой плотности зрительного пурпура. Таким путем глаз восстанавливает свою чувствительность.
Однако подобные выводы противоречили результатам шумового анализа чувствительности глаза, который показал, что собственная чувствительность глаза не может изменяться более чем в 10 раз чем переходе от темноты к яркому свету. Достоинство метода шумового анализа состояло в том, что его результаты не зависит от конкретных физических или химических моделей самого зрительного процесса. Чувствительность измерялась по абсолютной шкале, при этом постулировались лишь квантовая природа света и хаотический характер распределения фотонов.
Как же тогда объяснить тысячекратное и даже большее усиление способности видеть, наблюдаемое в процессе темновой адаптации? Напрашивалась определенная аналогия между этим процессом и работой таких устройств, как радио- и телевизионные приемники. Если при перестройке приемника с сильной станции на слабую звук оказывается почти неслышимым, слушатель берется за ручку регулировки громкости и доводит уровень звучания слабой станции до удобной величины. Существенно, что чувствительность радиоприемника остается постоянной как при переходе от сильной станции к слабой, так и при регулировке громкости. Она определяется только характеристиками антенны и первой лампы усилителя. Процесс «поворота ручки громкости» меняет не чувствительность приемника, а лишь «уровень представления» слушателю. Вся операция перестройки с сильной на слабую станцию, включая и продолжительность процесса регулировки громкости, полностью аналогична весьма длительному процессу зрительной темновой адаптации.
За то время, пока осуществляется темновая адаптация, коэффициент усиления «усилителя» возрастает в результате химических реакций до нужного «уровня представления». Собственная чувствительность глаза остается почти постоянной в период темновой адаптации. Нам не остается ничего иного, как предположить, что в зрительном процессе участвует некий усилитель, действующий между сетчаткой и мозгом, и что коэффициент его усиления изменяется в зависимости от освещенности: при больших освещенностях он мал, а при низких — велик.
↑ Автоматический контроль усиления
Вывод о том, что зрительный процесс обязательно включает в себя автоматический контроль усиления, мы сделали в предыдущем разделе на основании сильных изменений в кажущейся чувствительности, с которыми мы сталкиваемся при темновой адапта¬ции, и относительного постоянства собственной чув¬ствительности, которое следует из шумового анализа зрительного процесса.
К подобному же заключению мы придем, если рассмотрим другие, более прямые данные, встречаю¬щиеся в литературе. Известно, что энергия нервного импульса на много порядков больше энергии тех не¬скольких фотонов, которые требуются для запуска нервного импульса на абсолютном пороге чувствительности. Поэтому для генерации нервных импульсов нужен механизм с соответственно высоким коэффициентом усиления непосредственно на сетчатке. Из ранней работы Хартлайна по электрической регистрации зрительных нервных импульсов мечехвоста было известно, что частота нервных импульсов возрастает с увеличением интенсивности света не линейно, а лишь логарифмически. Это означает, что при высоких освещенностях коэффициент усиления меньше, чем при низких.
Хотя энергия нервного импульса точно не известна, приближенно ее можно оценить, предполагая, что запасенная энергия импульса соответствует напряжению 0,1В на емкости 10-9[/sup]Ф (это емкость 1см внешней оболочки нервного волокна). Тогда электрическая энергия составляет 10-11 Дж, что в 108 раз больше энергии фотона видимого света. Мы, конечно, можем ошибиться в оценке энергии нервного импульса на несколько порядков величины, но это не ставит под сомнение сделанный нами вывод о том, что непосредственно на сетчатке должен происходить процесс чрезвычайно большого усиления, и только благодаря этому энергия нескольких фотонов может вызвать нервный импульс.
Прогрессирующее уменьшение усиления по мере возрастания интенсивности света отчетливо наблюдается в данных Хартлайна, согласно которым частота нервных импульсов при увеличении интенсивности света медленно возрастает по логарифмическому закону. В частности, при увеличении интенсивности света в 104 раз частота возрастает лишь в 10 раз. Это означает, что коэффициент усиления уменьшается в 103 раз.
Хотя конкретные химические реакции, лежащие в основе процесса усиления, не известны, по-видимому, вряд ли что можно предположить, кроме некой формы катализа. Фотон, поглощенный молекулой чувствительного материала (родопсина), вызывает изменение ее конфигурации. Последующие этапы процесса, в ходе которого возбужденный родопсин оказывает каталитическое действие на окружающий биохимический материал, пока не ясны. Однако разумно предположить, что каталитическое усиление будет уменьшаться при увеличении интенсивности света или числа возбужденных молекул, так как при этом должно уменьшаться количество катализируемого материала, приходящееся на одну возбужденную молекулу. Можно также предположить, что скорость истощения катализируемого материала (световая адаптация) велика по сравнению со скоростью его регенерации (темновая адаптация). Известно, что световая адаптация происходит в течение доли секунды, тогда как темновая адаптация может продолжаться до 30 мин.
↑ Зрительный шум
Как мы неоднократно подчеркивали, наша зрительная информация ограничена случайными флуктуациями в распределении падающих фотонов. Следовательно, эти флуктуации должны быть видимыми. Однако мы замечаем это не всегда, во всяком случае, при нормальных освещенностях. Отсюда вытекает, что при каждом уровне освещенности коэффициент усиления оказывается в точности таким, чтобы фотонный шум был едва различим или, лучше сказать, почти неразличим. Если бы коэффициент усиления был больше, то это не дало бы дополнительной информации, а лишь способствовало бы увеличению шума. Если бы коэффициент усиления был меньше, то это привело бы к потере информации. Аналогично усиление телевизионного приемника следует выбирать так, чтобы шум находился на пороге видимости.
Хотя фотонный шум нелегко заметить при обычных освещенностях, автор на основании своих собственных наблюдений убедился, что при яркостях примерно 10-8—10-7 ламб однородно освещенная стена приобретает такой же флуктуирующий, зернистый вид, как и изображение на экране телевизора при наличии сильных шумов. Более того, степень видимости этого шума сильно зависит от степени возбужденности самого наблюдателя. Такие наблюдения удобно производить непосредственно перед сном. Если во время наблюдений в доме раздается какой-то звук, предвещающий появление неожиданного или нежелательного посетителя, то поток адреналина мгновенно увеличивается и одновременно «видимость» шумов заметно возрастает. В этих условиях механизмы самосохранения вызывают увеличение коэффициента усиления зрительного процесса (точнее амплитуды сигналов, поступающих от всех органов чувств) до такого уровня, который гарантирует полное восприятие информации, то есть до уровня, когда шум легко наблюдаем.
Конечно, такие наблюдения субъективны. Де Врис — один из немногих, кто, помимо автора данной книги, отважился опубликовать свои сравнительные наблюдения. Однако многие исследователи в частных беседах рассказывали автору о подобных результатах.
Очевидно, что вышеописанные картины шумов обусловлены падающим потоком фотонов, поскольку они отсутствуют в «совершенно черных» участках изображения. Наличие лишь нескольких освещенных участков достаточно для того, чтобы величина коэффициента усиления установилась на таком уровне, при котором другие, гораздо более темные участки кажутся совершенно черными.
С другой стороны, если наблюдатель находится в совершенно темной комнате или его глаза закрыты, у него не создается зрительного ощущения однородного черного поля. Скорее он видит серию слабых, движущихся серых изображений, которые часто упоминались в более ранней литературе под названием «е15епНсЫ;» , то есть как нечто возникающее в пределах самой зрительной системы. Опять-таки появляется искушение рационально истолковать эти наблюдения, предположив, что ввиду отсутствия реального светового изображения, которое привело бы к установлению определенной величины коэффициента усиления, последний достигает максимальной величины в поисках объективной зрительной информации. При таком усилении обнаруживаются шумы самой системы, которые, по-видимому, связаны с процессами теплового возбуждения в сетчатке или возникают в каком-либо удаленном от нее участке нервной си« СТеМБ1.
Последнее замечание касается, в частности, процесса усиления зрительных ощущений, который, как говорят, происходит в результате приема различных веществ, вызывающих галлюцинации. Представляется весьма вероятным, что эффекты, вызываемые этими веществами, обусловлены увеличением коэффициента усиления мощного усилителя, находящегося в самой сетчатке.
Как мы уже отмечали, эмоциональное состояние, связанное с каким-то напряжением или повышенным вниманием наблюдателя, приводит к значительному увеличению усиления.
↑ Послеобразы
Существование механизма контроля усиления на сетчатке позволяет дать очевидное объяснение различным наблюдениям, при которых человек смотрит на яркий объект, а затем переводит взгляд на нейтральную серую стену. При этом в первый момент человек еще видит некий переходной образ, который затем постепенно исчезает. Например, яркий черно- белый объект дает переходный дополнительный образ (послеобраз) в виде фотографического негатива оригинала. Яркий красный объект дает дополнительный цвет — зеленый. В любом случае в той части сетчатки, куда попадает изображение яркого объекта, усиление уменьшается, так что, когда на сетчатке отображается однородная поверхность, прежде яркие участки сетчатки дают в мозг сигнал меньшей величины и появляющиеся на них изображения кажутся темнее, чем окружающий фон. Зеленый цвет послеобраза ярко- красного предмета показывает, что механизм усиления не только локально меняется на различных участках сетчатки, но и на одном и том же участке действует независимо для трех цветовых каналов. В нашем случае усиление для красного канала моментально уменьшилось, что привело к появлению на нейтральной серой стене изображения в дополнительном цвете.
Стоит отметить, что послеобразы не обязательно всегда отрицательны. Если, глядя на ярко освещенное окно, закрыть глаза, затем мгновенно на некоторое время открыть' их, как бы пользуясь фотографическим затвором, и потом снова плотно закрыть, то в течение нескольких секунд или даже минут после- образ будет положительным (по крайней мере вначале). Это вполне естественно, так как время затухания любого процесса фотовозбуждения в твердом теле конечно. Известно, что глаз накапливает свет в течение 0,1—0,2 с, поэтому среднее время его фотовозбуждения также должно составлять 0,1—0,2 с, и за время порядка секунд фотовозбуждение спадает до все меньшего уровня; послеобраз остается видимым, поскольку усиление продолжает возрастать после того, как мы закроем глаза. Если в процессе наблюдения положительного образа в глаз попадает небольшое количество постороннего света, то этот образ немедленно превращается в отрицательный по причинам, указанным в предыдущем разделе. По мере того как посторонний свет возникает или исчезает, мы можем переходить от положительного послеобраза к отрицательному и обратно. Если в темной комнате смотреть на конец зажженной сигареты, движущейся по кругу, то зажженный конец будет восприниматься как полоска света конечной длины благодаря инерции зрительного восприятия (положительный послеобраз). При этом наблюдаемое изображение, подобно комете, имеет ярко-красную голову и голубоватый хвост. Очевидно, голубые компоненты света сигареты обладают большей инерцией, чем красные. Подобный эффект мы можем наблюдать, глядя на стену красноватого цвета: по мере уменьшения яркости до уровня ниже примерно 10-6 ламб она приобретает синий оттенок. Обе серии наблюдений можно объяснить, если предположить, что коэффициент усиления для голубого цвета достигает больших величин, чем для красного; в результате восприятие голубого цвета сохраняется до меньших уровней возбуждения сетчатки, чем красного.
↑ Видимость излучений высоких энергий
Зрительное восприятие инициируется электронным возбуждением молекул. Поэтому можно предположить существование определенного энергетического порога, но, вообще говоря, не исключено, что излучения высокой энергии будут также вызывать электронные переходы и окажутся видимыми. Если переход, вызывающий зрительное возбуждение, представляет собой резкий резонанс между двумя электронными энергетическими уровнями, то фотоны с большей энергией не будут эффективно возбуждать этот переход. С другой стороны, электроны или ионы высокой энергии могут возбуждать переходы в широком интервале энергий, и тогда они должны быть видимыми, поскольку на своем пути они оставляют плотные области возбуждений и ионизаций. В опубликованной ранее работе, где обсуждались проблемы видимости излучения с высокими энергиями, автор выразил некоторое удивление по поводу того, что до сих пор никто не сообщал о прямых визуальных наблюдениях космических лучей.
В настоящее время имеются некоторые данные, касающиеся проблемы видимости излучений в широком интервале высоких энергий. Прежде всего, уже известно, что ультрафиолетовая граница обусловлена поглощением в роговице. Люди, у которых по тем или иным причинам роговица была либо удалена, либо заменена более прозрачным веществом, действительно могут видеть ультрафиолетовое излучение.
О возможности видеть рентгеновское излучение немало говорилось на ранних стадиях исследования рентгеновских лучей. Публикации в этой области прекратились, когда стало известно о вредном воздействии рентгеновского излучения. Эти ранние наблюдения были неоднозначны, поскольку оставалось неясным, возбуждают ли рентгеновские лучи сетчатку непосредственно или посредством возбуждения флуоресценции в стекловидном теле. Некоторые более поздние и точные эксперименты говорят о том, что происходит прямое возбуждение сетчатки; об этом, в частности, свидетельствует восприятие резких теней от непрозрачных объектов.
Возможность визуального наблюдения космических лучей в настоящее время подтверждена рассказами космонавтов о том, что они видели полосы и вспышки света, когда кабина корабля находилась в темноте. Однако до сих пор остается неясным, связано ли это непосредственно с возбуждением сетчатки или с генерацией рентгеновского излучения в стекловидном теле. Космические лучи создают в любом твердом теле плотный след возбуждения, поэтому было бы странно, если бы они не могли вызвать непосредственное возбуждение сетчатки.
↑ Зрение и эволюция
Способность живых клеток считать фотоны или, по меньшей мере, реагировать на каждый фотон возникла на ранних этапах развития растительной жизни. Квантовая эффективность фотосинтеза оценивается для красного света примерно в 30%. В процессе фотосинтеза энергия фотонов используется непосредственно в определенных химических реакциях. Она не усиливается. Растение использует свет для питания, но не для получения информации, если исключить гелиотропные эффекты и синхронизацию биологических часов.
Использование же света для получения информации означает, что непосредственно на рецепторе должен быть создан в высшей степени сложный усилитель, благодаря которому ничтожно малая энергия фотонов превращается в значительно большую энергию нервных импульсов. ТОЛЬКО таким путем глаз способен передавать информацию в мышцы или мозг. Такой усилитель, по-видимому, появился на ранних этапах развития животной жизни, поскольку многие простейшие животные обитают в темноте. Следовательно, искусство счета фотонов было освоено задолго до появления человека.
Счет фотонов, безусловно, явился существенным достижением эволюционного процесса. Он оказался также наиболее сложным шагом в развитии зрительной системы. Для выживания нужна была гарантия, что вся доступная информация может быть зарегистрирована. При наличии такой гарантии адаптация зрительной системы в зависимости от конкретных потребностей определенного животного представляется уже более легким и вторичным успехом.
Такая адаптация приобрела большое разнообразие форм. Большинство из них, по-видимому, обусловлено очевидными причинами. Мы приведем здесь лишь несколько примеров для того, чтобы подтвердить тесную связь между оптическими параметрами и условиями жизни животного.
Структура сетчатки дневных птиц, таких, как ястреб, в несколько раз тоньше, чем у ночных животных, например у лемура. Очевидно, что у высоколетающего ястреба более высокое разрешение зрительной системы и соответственно более тонкая структура сетчатки оправданы высокой яркостью освещения в середине дня. Более того, в поисках полевой мыши ястребу безусловно необходима большая детальность зрительного образа. С другой стороны, лемур при его ночном образе жизни имеет дело со столь низкими уровнями освещенности, что его зрительные образы, качество которых ограничено фотонным шумом, крупнозернисты и не требуют большего, чем крупнозернистая структура сетчатки. В самом деле, при столь малой интенсивности света выгодно иметь линзы с большой апертурой (f/D) = 1,0), хотя эти линзы неизбежно должны давать плохое качество оптического изображения (рис. 16).

Кривая спектральной чувствительности человеческого глаза хорошо соответствует максимуму распределения дневного солнечного света (5500А). В сумерках максимум чувствительности глаза смещается к 5100 А, что соответствует голубоватому оттенку света, рассеянного небом после захода солнца. Можно было бы ожидать, что чувствительность глаза должна простираться в красную область по крайней мере до той длины волны, где тепловое возбуждение сетчатки начинает конкурировать с фотонами, попадающими извне. Например, на абсолютном зрительном пороге 10-9 ламб спектральная чувствительность глаза могла бы простираться примерно до 1,4 мкм, где такая конкуренция уже становится существенной. Остается не ясным, почему в действительности граница чувствительности глаза находится при 0,7 мкм, если только это ограничение не связано с отсутствием подходящего биологического материала.
Время накопления информации глазом (0,2 с) хорошо согласуется с временем нервной и мышечной реакции человеческой системы в целом. Наличие такой согласованности подтверждается тем, что специально сконструированные телевизионные камеры с временем релаксации 0,5 с или более оказываются явно неудобными и раздражающими в эксплуатации. Возможно, что у птиц время накопления зрительной информации меньше вследствие их большей подвижности. Косвенным подтверждением этому может служить тот факт, что некоторые трели или серии нот птицы «пропевают» столь быстро, что человеческое ухо воспринимает их как хор.
Наблюдается строгое соответствие между диаметром палочек и колбочек человеческого глаза и диаметром дифракционного диска в тот момент, когда отверстие зрачка близко к его минимальной величине (примерно 2 мм), которая устанавливается при высоких интенсивностях света. У многих животных зрачки имеют не круглую, а щелеподобную форму и ориентированы в вертикальном (например, у змей, аллигаторов) или в горизонтальном (например, у коз, лошадей) направлениях. Вертикальная щель обеспечивает высокую резкость изображения, ограниченную для вертикальных линий аберрациями линз, а для горизонтальных — дифракционными эффектами.
Вполне оправданы попытки убедительно объяснить приспособленность этих оптических параметров к образу жизни определенных животных. .
Зрительная система лягушки представляет собой поразительный пример адаптации в соответствии с ее образом жизни. Ее нервные связи устроены так, чтобы выделить движения привлекательных для лягушек мух и игнорировать постороннюю зрительную информацию. Даже в зрительной системе человека мы замечаем несколько усиленную чувствительность периферического зрения к мерцающему свету, что, очевидно, можно интерпретировать как охранную систему для предупреждения о надвигающейся опасности.
Наши рассуждения мы закончим несколько «домашним» замечанием. С одной стороны, мы подчеркивали, что человеческий глаз близко подошел к пределу, обусловленному квантовой природой света. С другой стороны, существует, например, выражение «видит как кошка», которое означает, что зрительная чувствительность домашней кошки в ее ночных похождениях значительно превышает нашу собственную. По-видимому, следует примирить эти два утверждения, заметив, что если бы мы решили разгуливать по ночам на четвереньках, то приобрели бы такую же способность ориентироваться в темноте, какой обладает кошка.
Итак, квантовая эффективность человеческого глаза меняется примерно от 10% при низких освещенностях до нескольких процентов при высоких освещенностях. Полный интервал освещенностей, в котором работает наша зрительная система, простирается от 10-10 ламб при абсолютном пороге до 10 ламб при ярком солнечном свете.
Непосредственно на сетчатке имеется биохимический усилитель с коэффициентом усиления, вероятно, более 106, который превращает малую энергию падающих фотонов в значительно большую энергию зрительных нервных импульсов. Коэффициент усиления этого усилителя изменяется в зависимости от освещенности, уменьшаясь при высоких освещенностях. Этими изменениями объясняется явление тем- новой адаптации и ряд эффектов, связанных с возникновением послеобразов. Зрительная система человека и животных служит свидетельством их эволюции и адаптации к внешним условиям.
----
Статья из книги: Зрение человека и электронное зрение А.Роуз 1977
.

Комментариев 0