Ганглиозные клетки сетчатки лягушки

Содержание:
Описание

Изучение ганглиозных клеток сетчатки лягушки представляет особый интерес не только из-за чрезвычайно выраженных отличий в морфологии всей сетчатки этого позвоночного по сравнению с другими видами, но и потому, что именно эти нейроны послужили основой представления о детекторах [Lettvin et al.,1959].
Дендриты ганглиозных клеток сетчатки лягушки изучались многократно как морфологически [Догель 1888b; Ramon у Cajal, 1972; Rozemeyer, Stolte, 1930; Шибкова, 1970, 1972; Калинина, 1975, 1976; Kalinina, 1974, 1976; Венслаускас, Гутаускас, 1972; Школьник-Яррос, 1971; Школьник-Яррос, Подуголышкова, Дюбина, 1975], так и морфофизиологически [Maturana et al., 1960; Lettvin et al., 1959, 1961; Pomcranz, 1972; Grusser, Grusser-Cornehls, 1976; Мантейфель, Школьник-Яррос, 1980].
В этом разделе, несколько более подробном, чем другие, собственные данные излагаются в сопоставлении с классическими данными Ramon у Cajal [1972], классификацию которого мы принимаем за основу (рис. 52),

а также классификациями ганглиозных клеток сетчатки лягушки, предложенными Lettvin et al. [1961] и Grusser, Grussеr-Cornehls [1976]. Сетчатка лягушки исследована на срезах, импрегнированных серебром по Гольджи [Ramon у Cajal, 1972; Мантейфель, Школьник-Яррос, 1980], окрашенных прижизненно метиленовым синим [Maturana et al.,1960],а также на тотальных препаратах сетчатки, импрегнированиых серебром по Грос- Бильшовскому - Лаврентьеву [Kalinina, 1974, 1976]. В этих работах были изучены сетчатки нескольких видов лягушек: Rana temporaria L., R. esculenta L., R. pipiens Shr., R. ridibunda Pall. Отличительные особенности ганлиозных клеток сетчаток лягушек различных видов рода Rana не велики, что позволяет обсуждать эти данные в одном разделе.
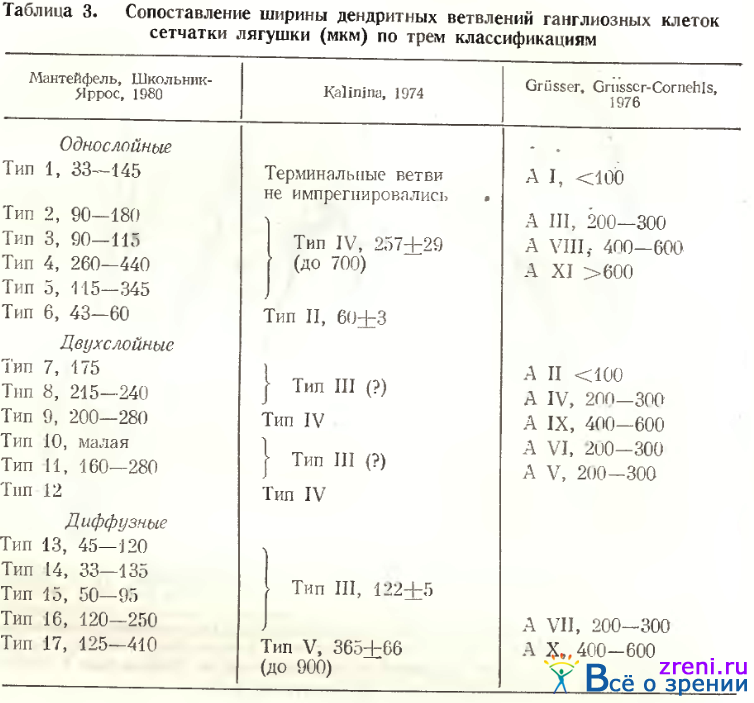
Все авторы использовали в качестве классификационных признаков размер сомы и дендритных ветвлений, а также характер ветвления дендритов во внутреннем плексиформиом слое (слое 7); окончание дендритов на разных уровнях (в подслоях) енутреннего плексиформного слоя; направление, мощность и число основных дендритов; степень разветвленности промежуточных и конечных дендритных ветвей, наличие пуговок, шипиков и варикозиостей на них. В табл. 1

сопоставлены типы ганглиозных клеток сетчатки лягушки пяти существующих ныне классификаций. Одному типу клеток классификации А. В. Калининой могут соответствовать несколько типов клеток более дробной классификации Е Г. Школьник-Яррос. В этом случае в таблице фигурной скобкой связаны входящие в этот тип разновидности ганглиозных клеток. В двух случаях рядом с указанным типом поставлен в скобках "вопросительный знак, что означает неуверенность авторов в таком сопоставлении.
Внутренний плексиформный слой сетчатки лягушки, по нашим данным, может быть подразделен на несколько подслоев, не менее 10 (см. сноску1, рис. 1; 53; A; 54).


Рис.54. Схема подслоев внутреннего плексиформного слоя (7) сетчатки лягушки Rana temporariа.
Метод Гольджи; am — тела амакриновых клеток; g — тела Ганглиозных клеток
Метод Гольджи; am — тела амакриновых клеток; g — тела Ганглиозных клеток
Однако для удобства сопоставления с другими классификациями мы условно вслед за Ramon у Cajal разделяем внутренний плексиформный слой на 5 подслоев, нумеруя их от внутреннего ядерного слоя к слою ганглиозных клеток, и классифицируем нейроны в соответствии с размещением дендритных ветвлений в этих слоях.
Переходим к описанию семнадцати типов ганглиозных клеток сетчатки лягушки. По характеру ветвления дендритов нейроны ганглиозного слоя могут быть разделены на две группы: слоистые (одно-, двух- и трехслойные) и диффузные.
↑ Однослойные ганглиозные клетки
Тип 1. Описание однослойных клеток сетчатки лягушки Ramon у Cajal [1972] начинает с клеток 4-го подслоя (рис. 52, 4с, 6е. Они отличаются малым размером тела, имеют один восходящий дендрит, делящийся на 2—4 ветви. Многочисленные терминальные веточки сильно изогнуты, снабжены коллатеральными Шипиками. Среди однослойных ганглиозных клеток этот тип (рис. 55, 56)

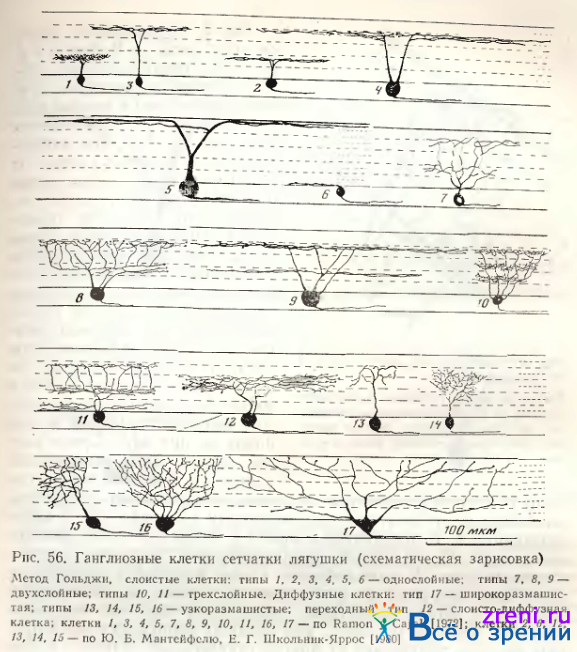
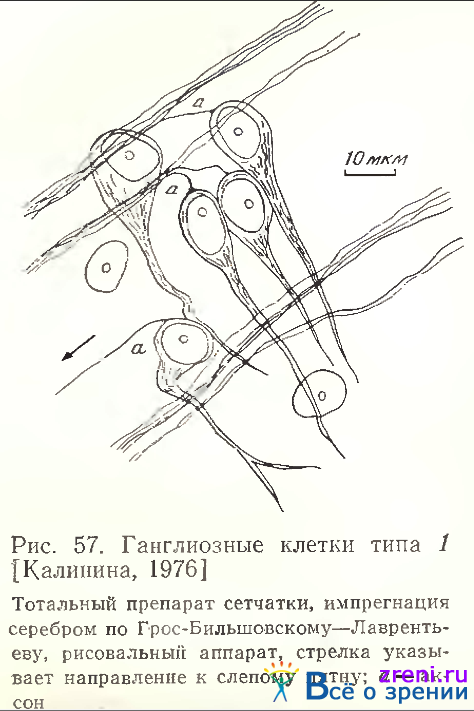
занимает особое положение в связи с тем, что, во- первых, все авторы единодушны в выделении его в самостоятельную классификационную единицу, а во-вторых, это самые малые и самые многочисленные среди всех типов клетки, составляющие 94—96% от общего числа нейронов ганглиозного слоя [Шибкова 1970; Kalinina, 1976]. Аксоны этих кустовидных клеток импрегнируются по Гольджи с трудом, однако нам удалось с помощью нейрофибриллярной методики многократно наблюдать отхождение аксона (рис. 57).
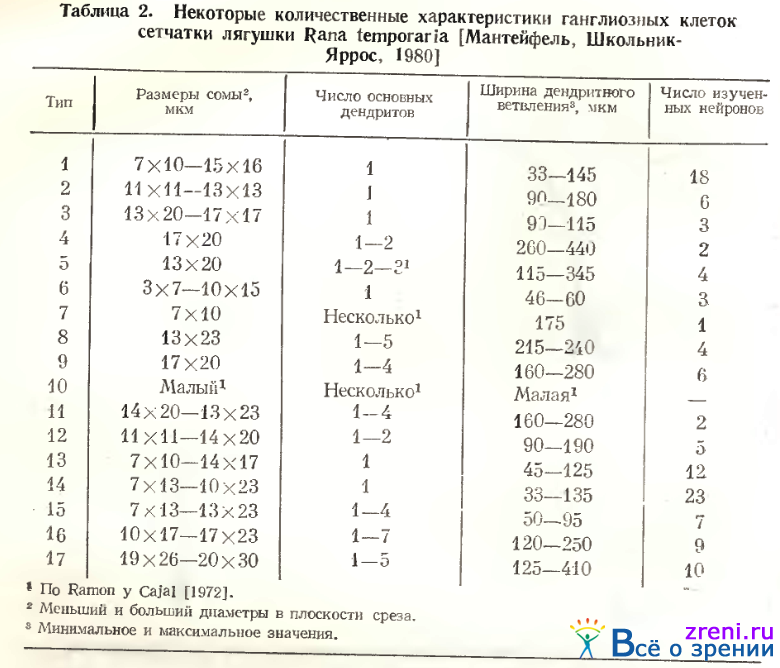
В то же время слоистое компактное ветвление их дендритов в 4-м подслое не обнаруживается с помощью этой методики, зато четко импрегнируется по Гольджи. Неудачи в окраске аксонов заставили Ramon у Cajal предположить, что это смещенные в ганглиозный слой амакриновые клетки. Кроме шипиков с типичными синаптическими головками, можно отметить у нейронов типа 1 многочисленные пуговки по ходу ветвящихся извилистых дендритов (см. рис. 55). Дендритные терминали часто образуют вилообразные ветвления, иногда подобие небольшой нежной корзинки. Размеры сомы и ширина дендритного ветвления нейронов типа 1 приведены в табл. 2.
Тип. 2. Кроме однослойных клеток — слоистых густоветвистых однодендритных (тип 1), наблюдаются типичные однослойные, однодендритные, но редковетвистые клетки с небольшим количеством синаптических образований и несколько большим размахом ветвлений (см. табл. 2), которые мы выделяем в тип 2 (рис. 56, 58, Б).
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Ветвления этих клеток иногда простираются кроме 4-го подслоя и в 3-й подслой внутреннего плексиформного слоя. Для упрощения номенклатуры и облегчения сравнения с сетчаткой других позвоночных мы подобные клетки называем кустовидными, густо- (тип 1) и редковетвистыми (тип 2). Однослойные клетки второго подслоя в соответствии с классификацией Ramon у Cajal мы также разделяем на два типа: 3 и 4 (см. рис. 56).
Тип 3. Тело клеток этого типа средней величины (см. рис. 58, В табл. 2). Восходящий дендритный ствол делится на 2—4 ветви (чаще всего на уровне 2-го подслоя), которые распространяются на большие расстояния в радиальном направлении, разветвляясь в узкой зоне внутреннего синаптического слоя. Вторичные дендриты гладкие, слабо извитые, с весьма малым количеством еле различимых пуговок. В некоторых случаях у нейронов типа 3 выявляются утолщения на уровне деления основных дендритных восходящих стволов. Ramon у Cajal описывает нейроны этого типа крупных и средних размеров (см. рис. 52, 4Ь, f).
Тип 4. Размеры перикариона несколько больше, чем у клеток предыдущего типа (см. рис. 52, 6а; 56; табл. 2), в то время как характер ветвления дендритов аналогичен, хотя дендриты более извилисты. В соответствии с описанием Lettvin et al. [1961] и Grusser, Grusser-Cornehls [1976] клетки типа 4 нашей классификации (тип II и AXI соответственно у этих авторов) самые крупные нейроны ганглиозного слоя (табл. 3).
Действительно, размах дендритных ветвей может достигать у них 600—700 мкм [Kalinina, 1974; Grusser, Grusser-Cornehls, 1976]. Lettvin et al. выделили нейроны этого вида в тип II, проиллюстрировав рисунком Ramon у Cajal (см. рис. 52, 6а).
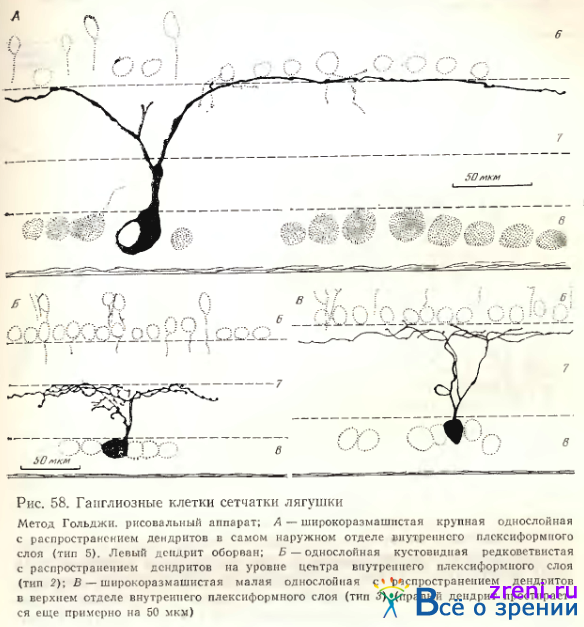
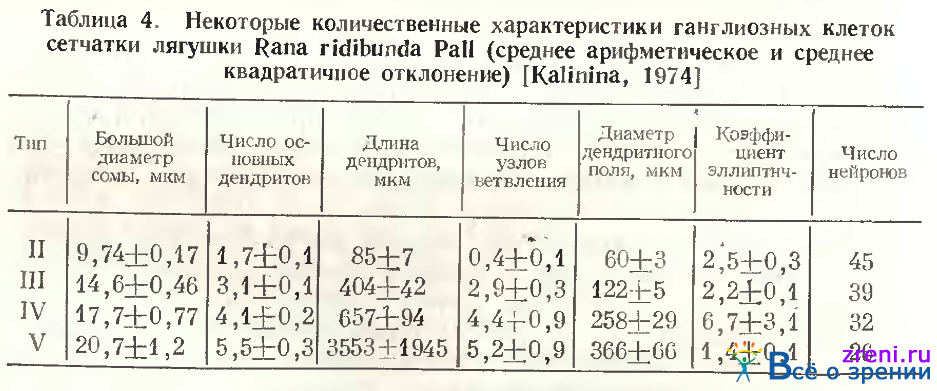
Тип 5. Однослойные клетки этого типа по Ramon у Cajal также очень крупные, с 2— 3 основными деидритами и густым ветвлением в первом подслое, редко встречающиеся (см. рис. 52, 4 d). Мы наблюдали чаще крупные клетки, от которых отходит лишь один основной дендрит, вскоре делящийся на два, либо отдающий крупные вторичные ветви, доходящие до самой границы внутреннего синаптического слоя с внутренним ядерным (см. рис. 58, А), где он расходится, образуя широкоразмашистое, но редковетвистое дендритное поле. Крупные синаптические площадки видны в зонах узлов ветвления дендритов, типичных пуговок и шипиков на них нет. Среди клеток этого типа встречаются нейроны с большим телом, однако с меньшим размахом ветвлений более тонких дендритов, снабженных пуговчатыми образованиями (см. табл. 2). В классификации А. В. Калининой нейроны 2, 3, 4 и 5-го типов объединены в тип IV по способу распространения дендритов в плоскости сетчатки. У этих нейронов самые вытянутые дендритные поля (см. коэффициент эллиптичности в табл. 4).
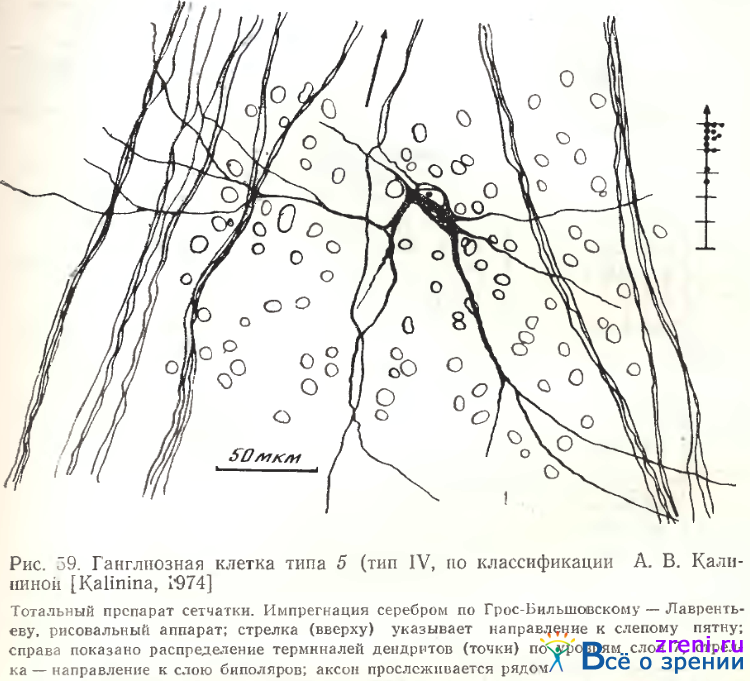
Характерна для них и четкая ориентация дендритного поля на плоскости сетчатки: длинная ось поля, как правило, перпендикулярна пучкам аксонов, тянущимся от других ганглиозных клеток к слепому пятну. Обычно 2—4 наиболее мощные, отходящие от противоположных сторон клетки дендритные ветви тянутся, разветвляясь по ходу на значительные расстояния (рис. 59).
Дендриты ветвятся в основном на расстоянии 30—80 мкм от ядросодержащей части нейрона (рис. 60).
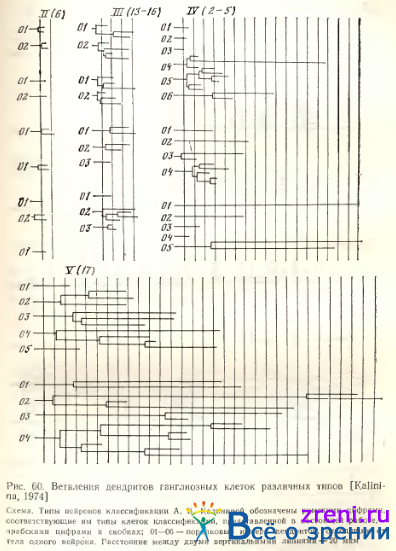
Схема показывает также, что для нейронов этого типа характерны одиночные, очень длинные неветвящиеся дендриты. Нейроны, объединяемые в тип I г, могут посылать дендриты в различные подслои внутреннего синаптического слоя, однако для каждого нейрона характерно ветвление и окончание дендритов в ограниченной по вертикали зоне. Эта зона располагается на различных уровнях плексиформного слоя от его наружной до внутренней границы (рис. 61, Б).

При этом чаще встречаются нейроны, дендриты которых расположены во внутренней половине слоя 7, чем нейроны, чьи дендриты ветвятся в его наружных зонах. Этот факт согласуется с данными других авторов. Некоторые количественные характеристики однослойных нейронов типа IV (по классификации А. В. Калининой) приведены в табл. 4.
Тип 6. Эта разновидность (всегда асимметричных) однослойных клеток, расположенных в 5-м подслое внутреннего плексиформного слоя или слое ганглиозных клеток (см. рис. 56, 62, 63, 4-я слева).
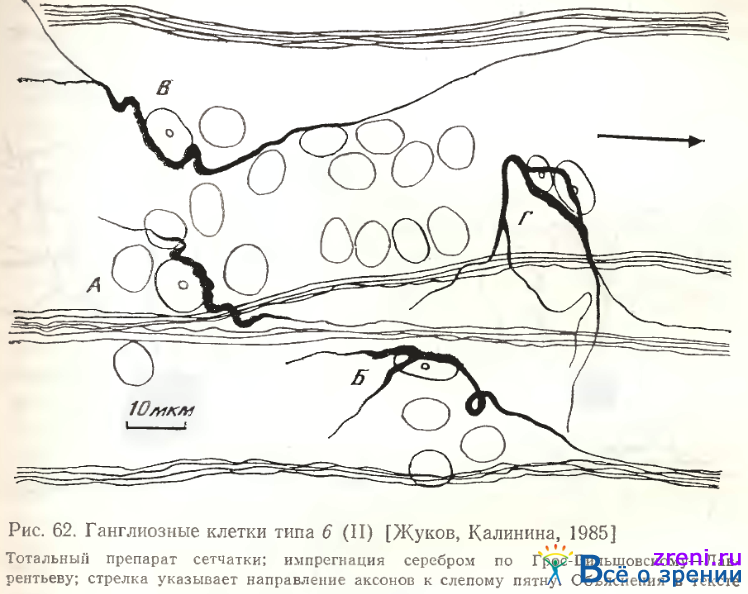
Впервые подобные нейроны описаны С. А. Шибковой (1970, 1972), исследовавшей нейронное строение внутреннего отдела сетчатки в эволюционном ряду позвоночных животных и обнаружившей этот тип ганглиозных клеток в сетчатке лягушек. Для нейронов этого типа характерна интенсивная импрегнация серебром дендритов и аксонов. При серебрении нейрофибриллярным методом в дендритах почти неразличимы нейрофибриллы, сливающиеся в сплошную аргирофильиую массу [Kalinina,1974].
Около 55% этих клеток биполярны (рис. 62, А). Однако аксон никогда пе отходит от тела клетки, почти всегда он является продолжением дендрита. Исключительно редко он отходит от базального участка дендрита, как это видно у клетки «Г» на этом же рисунке. Такое отхождение мы наблюдали у двуядерной разновидности этих нейронов, имеющих, как правило, более разветвленные дендриты. Дендритный «жгут» из слившихся нейрофибрилл, выявляемых по методике Грос-Бильшовского-Лаврентьева, проходит от дендрита через сому клетки, иногда разделяется в соме на две ветви (рис. 62, Г) и продолжается в противоположном от сомы направлении, постепенно истончаясь, в аксон. У всех исследованных 65-ти клеток аксон прослеживается вплоть до вхождения в пучок аксонов, направляющихся к слепому пятну.
Околоядерная часть нейрофибриллярного «жгута» часто имеет варикозности и неровные края. Отличительной чертой строения является большой диаметр проксимальной части дендрита, продолжающейся в аксон, по сравнению с противоположным ему дендритом. Длина «аксонного» утолщения дендрита, как правило, также больше, чем у противоположного дендрита. Некоторые дендриты нейронов типа 6 ветвятся один раз (см. рис. 62, В табл. 4). Количественные характеристики дендритов нейронов этого типа представлены в табл. 2, 3, 4. Дихотомия дендритов происходит на расстоянии 20 мкм от перикариона (см. рис. 60). Хотя большая часть дендритов нейронов 6-го типа оканчивается в 5-ом подслое, часть из них проникает до 3-го подслоя (см. рис. 61, А). В описаниях нейронов ганглиозного слоя у Ramon у Cajal и других авторов клетки этого типа не встречаются.
Таким образом, однослойные клетки могут быть как крупными, широковетвистыми, так и малыми, узковетвистыми. Зона их ветвлений не ограничивается двумя склеральными подслоями, как считал Ramon у Cajal [1972], однослойные клетки ветвятся в любом подслое внутреннего плексиформного слоя сетчатки лягушки (см. рис. 55, 56, 58, 61 и др.). Особенно убедительно это положение доказано по отношению к крупным широкоразмашистым клеткам [Kalinina, 1974]. Необходимо также отметить, что размеры клеток одного и того же типа существенно изменяются от центра к периферии сетчатки. Например, диаметр дендритного поля однослойных широкоразмашпетых клеток увеличивается от центра к периферии от 250 до 700 мкм, увеличивается и размер их перикариона.
↑ Двухслойные ганглиозные клетки
Ramon у Cajal различал три разновидности двухслойных ганглиозных клеток в сетчатке лягушки (см. табл. 1; рис. 52, 6f, g, с). Большинство из них мультиполярны, два горизонтальных сплетения дендритов размещаются в основном во 2-м и 4-м подслоях внутреннего синаптического слоя. При этом размах верхнего сплетения преобладает, как правило, по величине над размахом нижнего. Нижнее сплетение может занимать, помимо подслоя 4-го, и 3-й, на отростках дендритов наблюдаются пуговки и шипики. Двухслойные ганглиозные клетки (см. рис. 56) представлены нейронами разного калибра: малыми (тип 7), средними (тип 8) и крупными (тип 9), (см. табл. 2, 3). Чем больше тело клетки, тем сильнее развиты ее дендриты и меньше тонких восходящих веточек от нижнего сплетения к верхнему. Гигантскую двухслойную клетку Ramon у Cajal (см. рис. 52, 6с) Lettvin и соавторы обозначили как тип III с Н-образным ветвлением дендритов. По рисункам Ramon у Cajal Е-образные клетки соответствуют двухслойным клеткам Ramon у Cajal. Однако и тексте Lettvin et al. обозначают эту же клетку как многослойную (в табл. 1 это соответствие указано стрелкой).
Мы лишь кратко опишем здесь полученный нами материал по двухслойным нейронам. Из всех импрегнированных по Гольджи клеток, 11 относились к этой группе. По размерам тел выделялись крупные клетки, средине и малые (см. табл. 2). Наибольший размах дендритов у крупных клеток доходил до 280 мкм. Размах верхнего сплетения преобладал по величине над размахом нижнего сплетения в слое 3 и 4 (см. рис. 53, Б, клетка справа). У нейрона, также отнесенного нами к двухслойным (рис. 64, Б),

верхнее сплетение на границе с внутренним ядерным слоем обширное, состоит из множества тонких, длинных (до 208 мкм), извилистых дендритных ветвлений. Они образованы продолжением более крупных дендритов, отходящих либо от тела, либо от вторичных ветвей в подслоях 3; 4. На отростках наблюдаются пуговки и шипики. Нижнее сплетение редковствистое. Тело клетки, хотя частично и скрыто скоплением серебра, с уверенностью можно отнести к небольшим. По Grusser, Grusser-Cornehls [1976], трем типам двухслойных нейронов (по признаку возрастания размеров тела и дендритных ветвлений) соответствуют типы AII, AIV и AIX (см. табл. 1).
Сопоставление двухслойных нейронов, импрегнированных по Гольджи, с данными, полученными нейрофибриллярной методикой, затруднено, так как последняя выявляет не полностью мелкие дендритные веточки и терминали дендритов. Можно лишь предполагать, что среди нейронов типа III [Kalinina, 1974] могут находиться нейроны 7-го и 8-го типов, а среди гигантских нейронов типа IV — клетки 9-го типа классификации Ю. Б. Мантейфеля и Е. Г. Школышк-Яррос [1980]. Некоторые ганглиозные клетки было вообще трудно отнести с определенностью к какой-либо из описанных групп, поскольку сложно оценить степень слоистости и диффузности в ее структуре. Такая клетка представлена, например, на рис. 64, В. По-видимому, правильнее считать такие клетки переходными многослойно-диффузными формами.
↑ Трехслойные ганглиозные клетки
По описанию Ramon у Cajal, многослойные клетки в действительности трехслойны: малые (тип 10 нашей классификации) занимают своими ветвлениями 2, 3 и 4-й подслои (см. рис. 52, 4а), а более крупные (тип И) и 5-й подслой (см. рис. 50, 52, 6d), образуя два сплетения во 2-м и 4-м подслоях. В нашем материале типично трехслойпой являлась лишь одна клетка (рис. 65).

Довольно густое сплетение образуется у нее во 2-м подслое, где ветви снабжены многочисленными пуговками, реже шипиками. Другие многослойные клетки существенно отличаются от описанных Ramon v Cajal. Дендритные ветвления соответствуют не трем подслоям, а большему их количеству, в некоторых случаях удается видеть горизонтальное их расположение во всех подслоях. Grusser, Grusser-Cornehls эти клетки обозначили как тип AV — среднего размера клетки с тремя уровнями разветвления дендритов.
Некоторые нейроны сетчатки лягушки имеют характер диффузных, т. е. дендритные ветвления занимают почти всю ширину внутреннего плексиформного слоя (или 2—3 подслоя), однако при полном обзоре вссх отростков удается видеть тенденцию многих из них располагаться параллельно поверхности сетчатки. Если у слоистых нейронов, особенно у однослойных, эта тенденция выражена с предельной четкостью, то у описываемой группы она лишь намечена. Из 115 ганглиозных клеток, импрегнированных серебром по Гольджи, 5 можно было отнести к таким переходным диффузно-слоистым (см. табл. 1, 2), мы выделяем их в тип 12. Все клетки были однодендритными, кроме одной, с большим количеством (8) отходящих от тела дендритов. Пример диффузно-слоистого нейрона с распространением отросткоз по всему внутреннему плексиформному слою приведен на рис. 64, В, с ограниченным распространением — на рис. 56 (12).
↑ Диффузные ганглиозные клетки
Ramon у Caial наблюдал две разновидности диффузных клеток. Первая отличалась малыми размерами и весьма тонкими, густыми н сложными дендритными ветвлениями (см. рис. 52, 4е); вторая — большей величины, с более редкими, но обширными ветвлениями (см. рис. 52, 6Ь). Клетки мультиполярны, а их ветвления занимают всю ширину внутреннего плексиформного слоя. В тексте синаптические образования на дендритах не отмечены, но на рисунках можно видеть типичные пуговки на конечных ветвлениях у второго варианта клеток. Нам удалось выявить обе разновидности, наблюдавшиеся Ramon у Cajal, и еще несколько вариантов густо- и редковетвистых ганглиозных клеток.
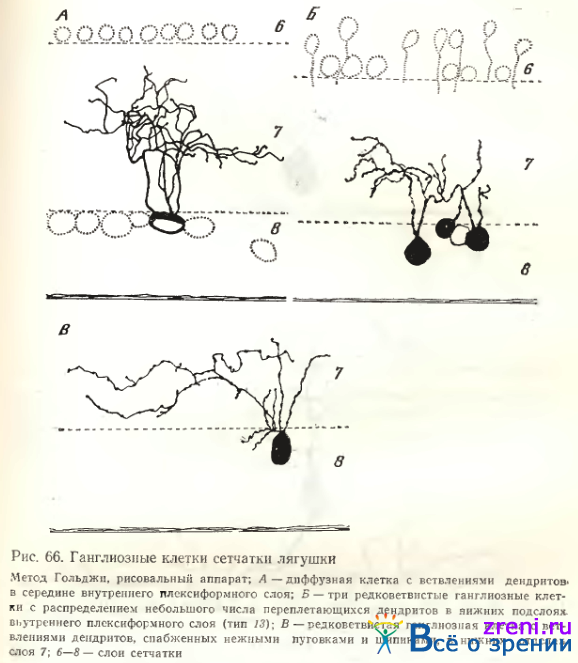
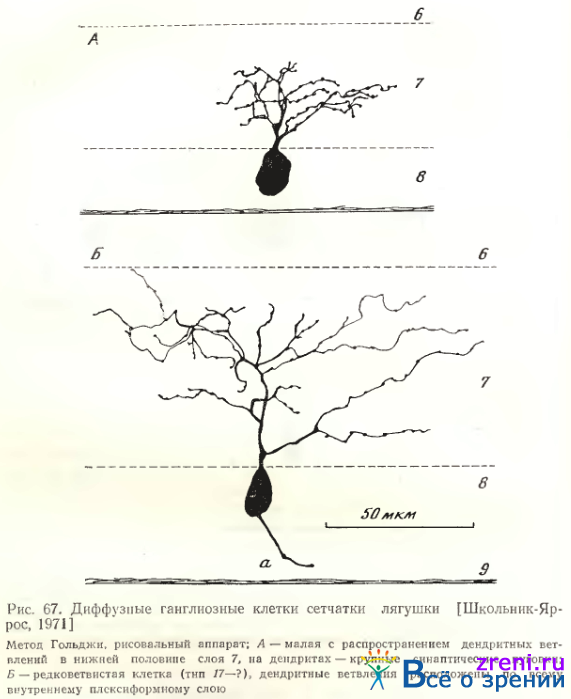
Тип 13. Среди редковетвистых диффузных клеток выделяется группа, которую мы отнесли к узкоразмашистым (тип 13). Их разветвления, ограниченные 2—3-мя подслоями внутреннего синаптического слоя, занимают небольшую зону: 45—120 мкм (см. рис. 56; 66, Б; 67, Л; табл. 2).
У некоторых из них очень нежные дендриты с небольшими пуговками и шипиками (рис. 66, В).
Тип 14. Среди густоветвистых диффузных нейронов, исследованных нами, выделяется группа ганглиозных клеток среднего* размера (см. табл. 2) с очень нежным ветвлением дендритов и множеством синапсов на них (рис. 68).
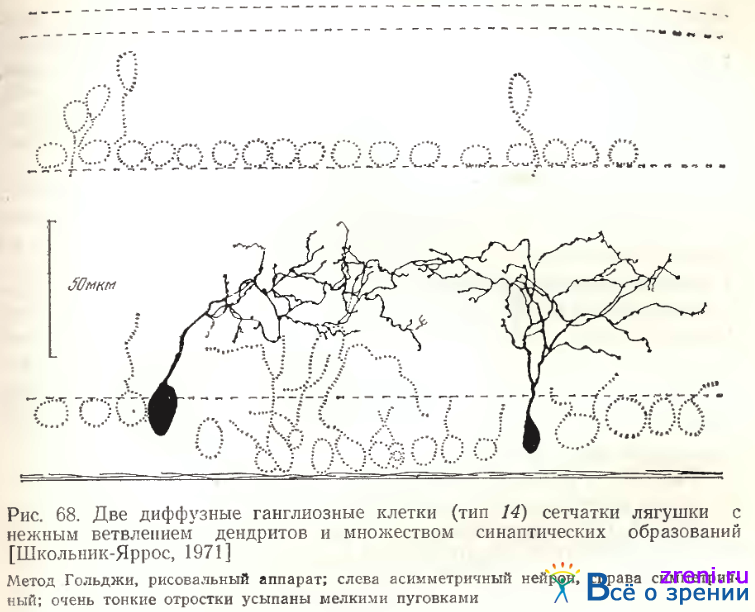
Тонкая сеть разветвлений дендритов явилась основанием назвать их паутинообразными (тип 14). От тела клетки, как правило, отходит довольно тонкий (основной) дендрит, отдающий букет ветвлений примерно на уровне 4-го подслоя. Разветвления занимают 2—3 подслоя на вертикали внутреннего плексиформного слоя. Весьма извилистые, очень тонкие отростки дендритов с трудом прослеживаются под микроскопом. Они усыпаны отчетливыми пуговками и часто многочисленными небольшими шипиками. Размах дендритных ветвлений чаще всего не превышает 100 мкм (см. табл. 2, 3). Лишь в одном случае на крайней периферии сетчатки тело клетки было крупным, а размах дендритов достигал 135 мкм.
Тип 15. Другая разновидность густоветвистых диффузных клеток представлена на рис. 66, А. У многодендритной густоветвистой клетки отростки ветвятся в центре внутреннего плексиформного слоя, образуя подобие сети, занимающей 2—3 его подслоя. Диффузные густоветвистые клетки с ветвлением, распространяющимся в 2—3 подслоях внутреннего плексиформного слоя„ могут быть либо однодендритными (см. рис. 56, тип 15), либо многодепдритпыми (см. рис. 66. А). У таких клеток наблюдаются пуговки разных размеров па извилистых дендритных ветвлениях и редкие шипики с головками.
У многодендритных клеток этой группы также весьма четко выражены синаптические образования. Один из подобных нейронов находился на крайней периферии сетчатки и, несмотря на такую локализацию, имел небольшие размеры тела (13X7 мкм), а размах дендритных ветвлений лишь 96 мкм. Уровень его ветвления, включающий слои 2—3—4, отличает его от описанной Ramon у Cajal группы 1-го типа, где ветвления занимали весь поперечник внутреннего плексиформного слоя. Нахождение таких весьма небольших клеток на крайней периферии сетчатки позволяет думать, что мы имеем дело с растущим нейроном (как известно, рост и развитие сетчатки происходит у амфибий в течение всей жизни).
Тип 16. Густоветвистыс диффузные нейроны с ветвлениями по всему внутреннему плексиформному слою в нашем материале сходны с описанием Ramon у Cajal (см. рис. 52, 4с). К этому типу мы отнесли 9 нейронов из 115, импрегнированных по Гольджи. От тела отходят 2—3, либо много первичных дендритов, образующих весьма густые извилистые ветвления (рис. 69).
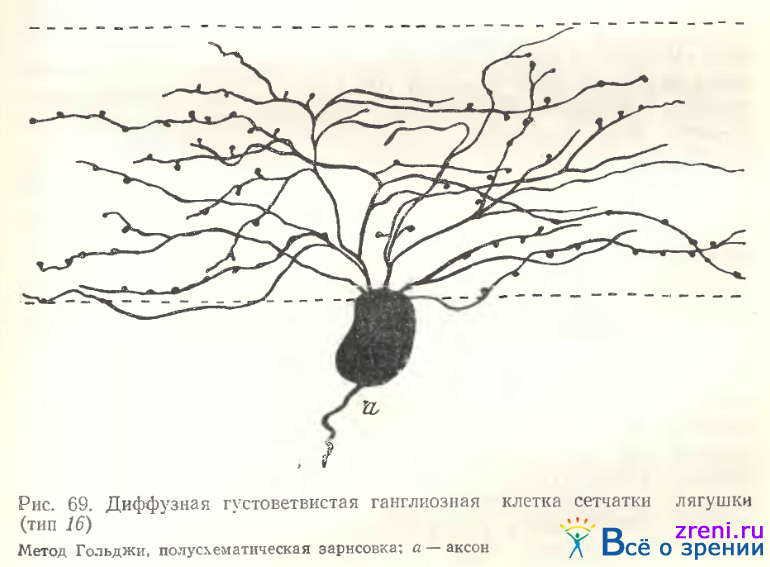
Размах дендритов от 120 мкм до 250 мкм (см. табл. 1, 2, 3). Все отростки покрыты многочисленными пуговками, иногда шипиками с головками. С описанием и иллюстрацией Ramon у Cajal не совпадала лишь величина тела нейронов, она была либо средней (16 мкм), либо даже крупной (23 мкм). Другой вариант диффузного густоветвистого нейрона, ветвления которого также занимали весь поперечник внутреннего плексиформного слоя, представлен на рис. 70.
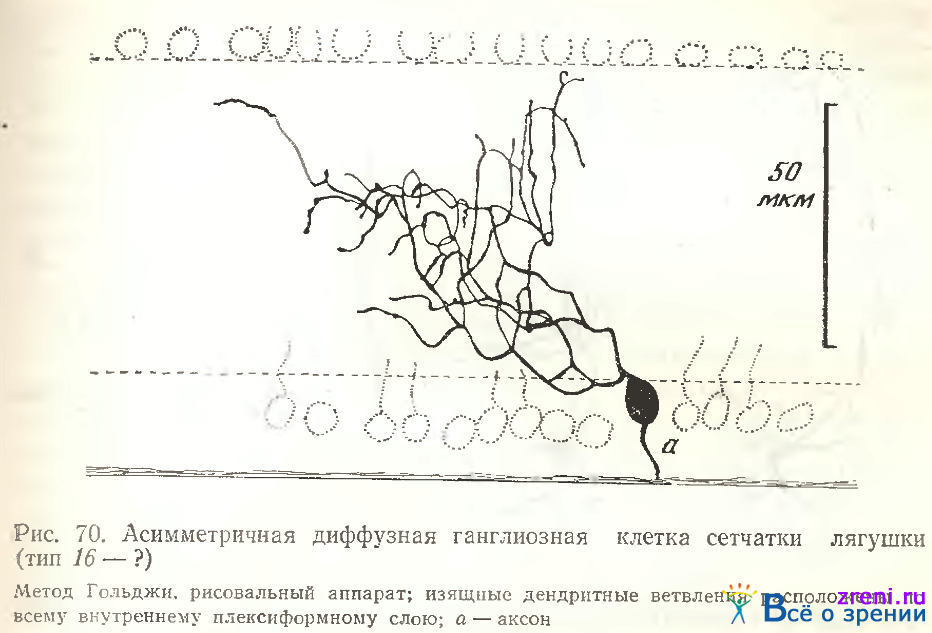
Сложный переплет извилистых отростков отличается, однако, от описанных выше клеток отсутствием явных пуговок и шипиков. От тела отходит лишь один отросток, сейчас же повторно делящийся. Ветвление у данной клетки полностью асимметрично.
Четыре типа (13—16) объединены А. В. Калининой в одну группу крупных и среднего размера диффузных нейронов — тип III. На тотальных препаратах сетчатки были изучены количественные характеристики их дендритных ветвлений (см. табл. 4, рис. 60). Дендритные терминали сконцентрированы в основном в витрсальной части внутреннего синаптического слоя, ближе к его середине, меньшая часть достигает более высоких уровней (см. рис. 61, А). Дендритные поля расположены, как правило, эксцентрично относительно перикариона и ориентированы на плоскости сетчатки так, что направление аксона, идущего к слепому пятну, противоположно направлению ветвлений первичных дендритов (рис. 71).
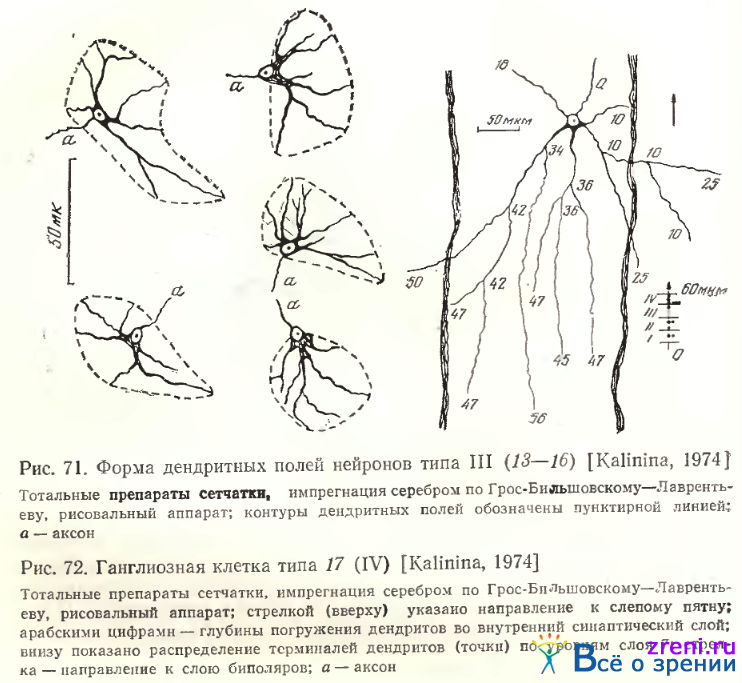
Чаще всего дендритные поля имеют овальную форму, но встречаются и яйцевидной, и бобовидной конфигурации. Размеры этих диффузных нейронов больше в периферической зоне сетчатки по сравнению с центральной, хотя различия эти в смежных зонах не всегда достоверны. Площадь дендритных полей увеличивается к периферии сетчатки в 2 раза.
Тип 17. Многодендритные редковетвистые нейроны (см. рис. 67, Б), относящиеся к этому типу, отличаются очень крупной сомой (см. табл. 2). Это второй вариант диффузной ганглиозной клетки сетчатки лягушки по Ramon у Cajal (см. рис. 52, 6Ь, табл. 1). Данный тип широковетвистого нейрона соответствует клеткам типа V А. В. Калининой. Дендриты этих диффузных нейронов ветвятся по всему внутреннему плексиформному слою (см. рис. 56, тип 17; 61; 72).
Каждый дендрит дихотомически ветвится на расстоянии 40—260 мкм от перикариона, окончания их распределены по всей толще слоя 7 (см. рис. 60; 61, А; табл. 4). Вторичные и третичные отростки значительно более гладкие, чем у клеток 1-го варианта. Размах дендритных ветвлений может быть весьма значительным, до 400 мкм в центре и до 900 мкм на периферии (см. табл. 2, 3, 4). В некоторых случаях выявляются пуговки по ходу и на концах ветвлений (по Гольджи), но более характерны довольно гладкие отростки с очень небольшим количеством варикозных утолщений.
Таким образом, диффузные ганглиозные клетки сетчатки лягушки представлены следующими разновидностями:
- густоветвистые с ветвлением по всему внутреннему плексиформному слою (тип 16);
- густоветвистые с ветвлением в ограниченных зонах плексиформного слоя (тип 15), с нежным ветвлением (тип 14);
- редковетвистые широкоразмашистые с ветвлением по всему внутреннему плексиформному слою (тип 17);
- редковетвистые узкоразмашнстые с ветвлением в ограниченных зонах плексиформного слоя (тип 13);
- переходные диффузно-слоистые (тип 12).
Lettvin et al., объединяют диффузные нейроны различных размеров в один тип V, приводя в качестве иллюстрации рисунок самого крупного нейрона этой разновидности из работы Ramon у Cajal. Grusser, Grusser-Cornehls в соответствии с классификацией Ramon у Cajal диффузные нейроны средних размеров и крупные выделяют в два типа: AVII и АХ (см. табл. 1).
Численное соотношение разновидностей ганглиозных клеток сетчатки лягушки исследовано в нескольких работах. Поскольку классификация каждого автора имеет свои особенности, сведения эти в каждом случае относительны. В исследованиях озерной лягушки С. А. Шибковой [1970] на одну крупную ганглиозную клетку в среднем приходится 5 нейронов среднего размера и 100 мелких. Указаны размерные вариации для этих клеток: 7—10 мкм —мелкие; 10—12 мкм —средние; 18,6—31,8 мкм — крупные. Количественные исследования численности и топографии па тотальных препаратах озерной лягушки показали те же соотношения [Kalinina, 1976].
Наиболее многочисленными в ганглиозном слое являются малые нейроны (тип 1). Их распределение по сетчатке определяет общую плотность ганглиозных клеток, которая уменьшается от центра сетчатки к периферии в 2—3 раза. Клеток средних размеров, к которым мы относим нейроны типа 6 (тип II по А. В. Калининой) почти в 60 раз меньше, чем мелких. Процентное содержание их несколько увеличивается к периферии сетчатки. Диффузные нейроны средней и крупной популяции (тип III) также чаще встречаются на периферии сетчатки, чем в центре. В целом их несколько больше, чем нейронов типа 6. Самые крупные нейроны, дендритные поля которых захватывают большие пространства плоскости сетчатки — самые малочисленные (0,5—1% от общего числа нейронов). Широкоразмашистые крупные однослойные нейроны типа IV представлены несколько большим числом (1,5—2%). Они чаще встречаются на периферии сетчатки и в средней зоне.
Исследования численности ганглиозных клеток в сетчатке травяной лягушки показали, что группа клеток с диаметром сомы 9—14 мкм составляет 67% всех изученных нейронов, с диаметром сомы 15—18 мкм — 29% и 23—25 мкм — 6% [Киселева, 1975; Маитейфель, Школьник-Яррос, 1980] (рис. 73).
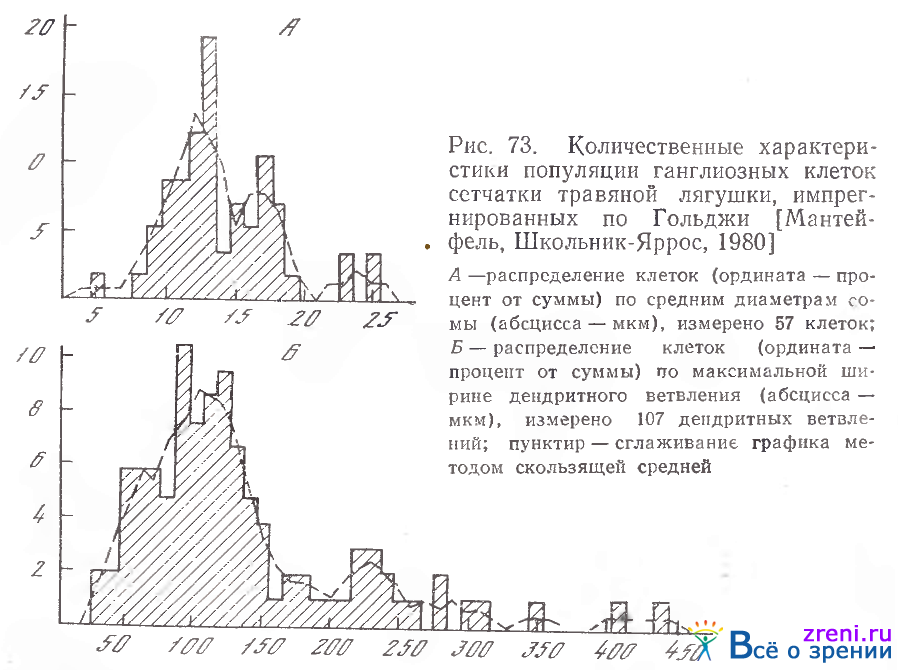
Авторами было установлено, что 71% всех изученных ганглиозных клеток сетчатки лягушки имеют максимальную ширину дендритных ветвлений от 40 до 150 мкм; общее количество ганглиозных клеток с размахом дендритных ветвлений до 200 мкм равно 83%- Клеток с шириной ветвления 215—246 мкм — 6%.
Таким образом, основополагающая классификация ганглиозных клеток амфибий Ramon у Cajal получила подтверждение и развитие в дальнейших исследованиях сетчатки лягушки. Она дополнена ранее не описанной разновидностью всегда асимметричных однослойных нейронов 5-го подслоя (тип 6). Среди однослойных небольшого размера нейронов выделен тип 2, характеризующийся рсдковетвистостью по сравнению с кустовидным густым и компактным ветвлением нейронов типа 1. Установлено, что существуют переходные формы между слоистыми и диффузными нейронами, которые отнесены к новому типу 12. В группе диффузных нейронов также выделены новые типы клеток, характеризующиеся своеобразием ветвления дендритов (13, 14 и 15).
Рассмотрение деталей строения, а также анализ количественных характеристик ганглиозных клеток сетчатки лягушки показывают, что разнообразие признаков и их вариабельность у этих нейронов сетчатки весьма велики.
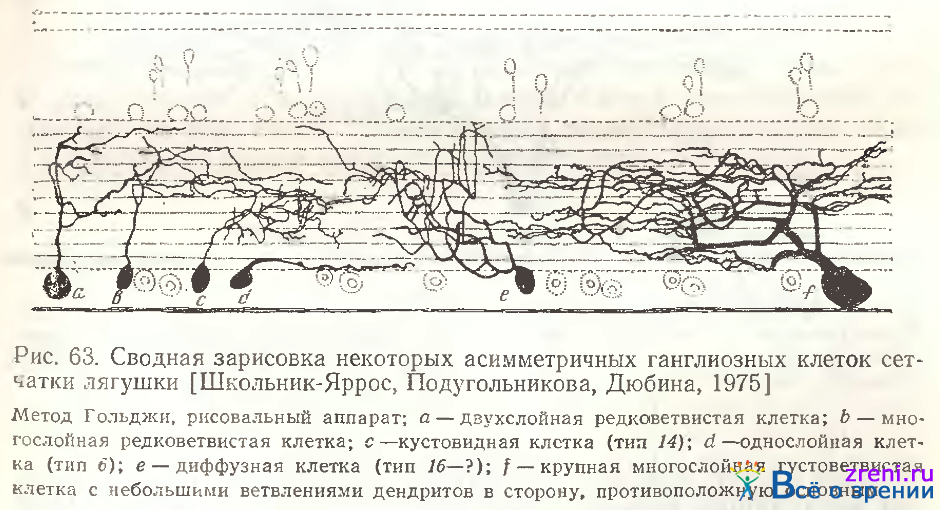
Попытки создания классификации ганглиозных клеток сетчатки лягушки оказались достаточно сложными в связи с почти непрерывным спектром переходных форм нейронов. Слоистые формы незаметно Слоистые формы незаметно переходят к диффузным, малые к большим. На сводной зарисовке (см. рис. 63) лишь некоторых из разновидностей асимметричных ганглиозных клеток хорошо видны как различные размеры тел, так и переход от чисто однослойного нейрона к редко ветвистым, а затем все более густоветвистым формам. Такое же затруднение при классификации встретилось нам при изучении биполярных клеток сетчатки лягушки, тоже весьма многообразных. Naka, Carraway [1975] при исследовании нейронов проксимальных слоев сетчатки Ictalurus punctatus отметили сходное положение, т. е. непрерывный спектр многообразных типов клеток, что затрудняет их классификацию.
Многообразие типов ганглиозных клеток в нашей работе установлено методиками импрегнации серебром. В дальнейшем желательно исследовать связи каждого типа нейрона с помощью Гольджи-электронной микроскопии либо пероксидазой хрена.
----
Статья из книги: Нейроны сетчатки | Е.Г. Школьник-Яррос, А.В. Калинина

Комментариев 0