Функциональная организация фиксационных поворотов глаз в процессе зрительного восприятия


Содержание:
Описание
В предыдущем разделе фиксационный поворот глаз рассматривался довольно абстрактно—как двигательный автоматизм (окуломоторный навык), характеристики которого определяются локализацией, конфигуративными особенностями поверхности объекта восприятия, решаемой задачей и социокультурной принадлежностью наблюдателя. В действительности же он имеет собственную организацию и подчиняется действию не только внешних, но и внутренних детерминант. К их числу относятся прогнозирование конечного и/или промежуточного результата, способ управления движениями глаз, ведущий уровень, на котором они строятся, сопряженность окуломоторики с другими двигательными актами наблюдателя и т. п. Действие внешних детерминант фиксационного поворота глаз всегда опосредствовано констелляцией его внутренних условий. Вез их учета анализ окулограмм оказывается неполным или недостаточно корректным. Именно внутренние условия целенаправленных движений являются, источником дисперсии амплитуды саккад, нелинейности и ограниченности влияния внешних детерминант, а также сложных по составу поворотов глаз. Анализу этих условий посвящен третий Раздел книги. Мы рассмотрим закономерности функционирования ГДС с измененным направлением зрительной обратной связи и каналы ее включения в процесс зрительного восприятия. Излагаемые здесь исследования конкретизируют возможности еще одного—электромагнитного—метода регистрации движений глаз.↑ Архитектоника фиксационного поворота
Фиксационный поворот глаз представляет собой целостный поведенческий акт, реализующий познавательное (перцептивное) отношение индивида к среде. Его результатом становится новое относительно устойчивое направление глаз, которое обеспечивает оптимальные условия восприятия значимого элемента или отношения среды. Этому направлению соответствует локализация (или тенденция локализации) проекции объекта в центральной области сетчатки. Визуальная данность субъекту значимых свойств действительности оказывается здесь и побуждением, и полезным эффектом движения, и условием его завершения.
Как и любой поведенческий акт фиксационный поворот глаз имеет сложную архитектонику (Рис. 3.1),
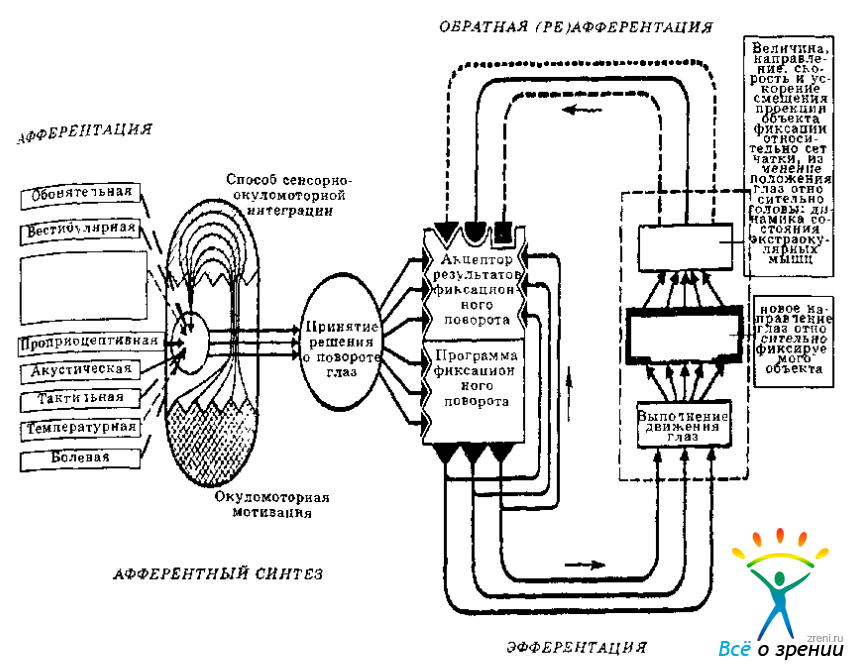
Рис. 3.1. Функциональная архитектоника фиксационного поворота глаз.
которая неплохо описывается в понятиях теории функциональных систем (Анохии, 1978, 1980). Он начинается с афферентного синтеза —интеграции исходных предпосылок движения: мотивации (потребности в визуальном выделении некоторого элемента или отношения среды), опыта выполнения окуломоторных актов в прошлом и афферентации, информирующей об актуальном состоянии организма (позе, положении головы, актуальной направленности глаз, открытости-закрытости век, перемещении наблюдателя и т. д.) и среды (расположении значимых элементов ситуации, их конфигурации, величине, удаленности, направлении, скорости движения и др.). Последнее непосредственно связано с действием внешних детерминант.
На основе афферентного синтеза принимается решение о перемещения глаз в сторону определенного предмета (формируется цель окуломоторного акта). Оно реализуется посредством двух механизмов. Во-первых, программы фиксационного поворота глаз, которая включает основные диспозиции движения (Becker, Fuch, 1969; Levy-Schoen, Blenc-Garin, 1974; Frost, Poppel, 1976). Благодаря этому механизму складывается динамическая интеграция окуломоторных единиц и их сонастроенность на определенный способ активности; наблюдатель приводится в состояние готовности выполнить окуломоторный акт с заданными характеристиками: направлением, амплитудой или скоростью (Гуревич, 1971; Teylor, 1962; Festinger, 1971). Во-вторых, с помощью аппарата предвидения двигательного и/или перцептивного эффекта — акцептора результатов действия. Благодаря этому механизму формируется модель будущего результата движений (наряду с афферентными «кодами» ожидаемого направления глаза она включает дубликат окуломоторной программы—«эфферентную копию» (Holst, 1954; Holst, Mittelstaedt, 1973)), с помощью которой оценивается обратная афферентация и корректируется неадекватная позиция глаз (Владимиров, Хомская, 1981; Howard, Templeton, 1966; Robinson, 1975).
Совокупность описанных процессов образует содержание латентного периода фиксационного поворота глаз, продолжительность которого зависит от характеристик визуальной информации, требований задачи, выбираемой стратегии, особенностей предшествующих движений глаз и других обстоятельств (Salthous, Ellis, 1980; Findlay, Crowford, 1983).
Исполнение, или собственно поворот глаз, осуществляется посредством саккадических и плавных движений. Они объединеняются в один, либо несколько перекрывающихся циклов «дрейф — саккада—дрейф», образующих окуломоторную структуру поворота. Выполняемые движения сопровождаются обратной (ре-) афферентацией, которая сопоставляется с моделью результатов фиксационного поворота. Существует несколько источников обратной афферентации: а) позиция и смещение проекции воспринимаемого предмета по сетчатке; этот источник считается основным (Андреева, Вергилес, Ломов, 1975; Stark, 1971; Jung, 1973); б) проприоцепция наружных мышц глаза (Becker, Fuchs, 1969; Skavenski, 1972; Weber, Daroff, 1972; Shebilske, 1976); в) раздражение вестибулярного аппарата, обеспечивающего координированность направлений глаз и головы (Курашвили, Бабняк, 1975; Bizzi, 1974; Barnes, 1975); г) проприоцепция скелетной мускулатуры, тактильная и акустическая сенсорные системы (Jones, Kabanoff, 1975; Buchele, Arnold, Brandt, 1977; Steinbach, 1987). Наличие обратной афферентации позволяет уточнять и корректировать программу и акцептор результатов действия в процессе выполнения фиксационного поворота глаз (Becker, Jurgens, 1979; Otts, Van Gisbergen, Eggermont, 1984). Последний совершается до тех пор, пока сохраняется значимое рассогласование Модели результата поворота и реафферентации.
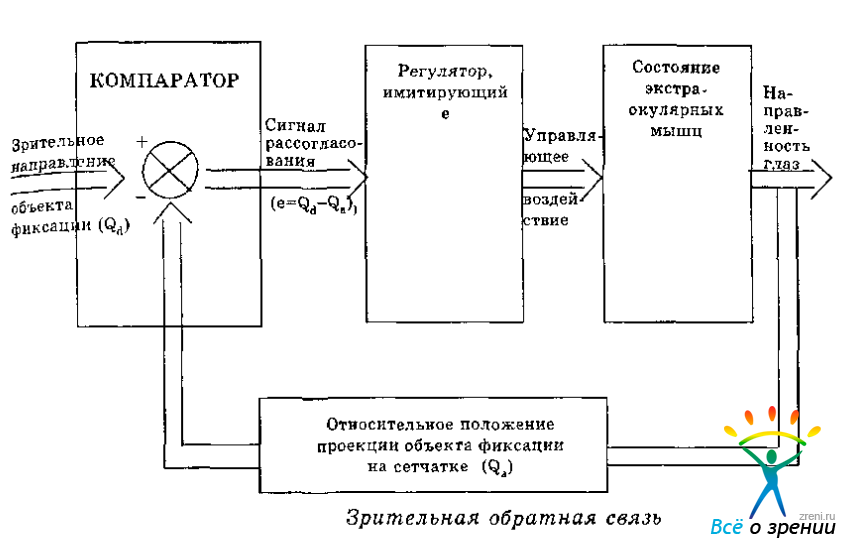
С позиции теории автоматического регулирования, которая привлекается исследователями для описания закономерностей oкуломоторной активности, механизм выполнения фиксационного поворота глаз рассматривается в терминах следящей системы с отрицательной зрительной обратной связью (Милсум, 1968; Андреева, Вергилес, Ломов, 1975; Арбиб, 1976). Подобная система реагирует на рассогласование между заданной и текущей направленностью глаз таким образом, что в результате Движения происходит элиминация входного сигнала (Рис. 3.2).
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Рис. 3.2. Оптический контур регулирования направленности глаз.
В этом случае отношение поворота глаз к соответствующему смещению проекции предмета по сетчатке выступает как характеристика канала зрительной обратной связи ГДС. Количествен, но ее можно выразить с помощью трех параметров: величины, знака и направления (Барабанщиков, Белопольский, Вергилес, 1978, 1980). За величину зрительной обратной связи принимают отношение амплитуды смещения проекции объекта фиксации по сетчатке к углу поворота глаз. Поскольку в нормальных условиях они совпадают, величина зрительной обратной связи равна единице. Однако в принципе, например, при патологии окуломоторного аппарата, она может быть как больше (визуальный угол, на который смещается проекция объекта, больше угла поворота глаз), так и меньше единицы (отношение углов обратное). Случаю, когда величина зрительной обратной связи равна нулю соответствуют условия стабилизации изображения объектов относительно сетчатки (Ярбус, 1965; Зинченко, Вергилес, 1969; Балонов, 1970), Под знаком зрительной обратной связи ГДС понимают вектор изменения входного сигнала. Если при повороте глаз ретинальный образ объекта фиксации приближается к fovea centralis (как это имеет место в норме), говорят об отрицательной, если удаляется в противоположную сторону—о положительной зрительной обратной связи. Так как регулирование позиции глаза осуществляется по крайней мере в двух координатах, вектор изменения входного сигнала (?) по разным причинам может варьировать в пределах -180° ? ? ? 180°, а адекватной характеристикой зрительной обратной связи выступает не ее знак, а направление.
Зрительная обратная связь ГДС—функциональное образование, имеющее смысл только по отношению к прямой связи. Если последняя задает цель регулирования, или желаемое направление глаз, то первая позволяет достигать его оптимальным образом и удерживать необходимое время. Несмотря на то, что с изменением предмета восприятия содержание прямой и обратной связи ГДС также изменяется, способ их функционирования остается неизменным.
Вертикальная организация фиксационного поворота глаз может быть представлена в виде иерархии разнородных процессов трех уровней.
1) Уровень интенции и произвольного контроля движений выражает отнесенность окуломоторной активности к субъекту восприятия (наблюдателю). Здесь складываются намерение и «первичный проект» движений, «сформулированные» на языке зрительно данных предметных отношений действительности: куда смотреть, на что обратить внимание, что контролировать, в какой последовательности, как долго и т. п. «Первичный проект» непосредственно увязан со схемой ситуации и планом выполняемого наблюдателем действия и соотносится с системой координат внешнего пространства (среды). На этом уровне ставятся или принимаются зрительные (окуломоторные) задачи и осуществляется произвольный контроль их исполнения.
На (2) уровне механизмов организации движений «первичный проект» переводится в цепочки исполнительных команд и критерии адекватности их выполнения. События этого уровня описываются на языке информационных потоков или эфферентноафферентных процессов ГДС, соотнесенных с ретинальной и окуломоторной системами координат. Здесь учитываются либо устанавливаются взаимосвязи окуломоторики с другими двигательными актами (поворотами головы, локомоциями и т. п.).
(3)Уровень феноменов движений характеризует способ выполнения поворота глаз. Здесь доминирует язык моторных единиц, сокращений и растяжений экстраокулярных мышц, развертывается действие активных и реактивных сил, влияющих на глазное яблоко и т. п. Выполняемые движения соотносимы как с внешней, так и с ретинальной(окуломоторной) системами координат.
Хотя содержание, функции и язык событий, происходящих на каждом из уровней, существенно отличаются друг от друга, они подчинены выполнению общей задачи и выступают как одно целое.
Таким образом, с точки зрения внутреннего строения целенаправленный поворот глаз слабо напоминает окуломоторную реакцию на оптический стимул, или «фиксационный рефлекс». Это—сложноорганизованное многомерное целое, которое складывается и функционирует в акте зрительного восприятия в Качестве и его предпосылки, и его результата(продукта). Фиксационный поворот глаз включает моменты прогнозирования (цель, акцептор результатов действия, модель потребного будущего, эфферентная и афферентная копни и т. п.), эфферентной готовности (установка, эфферентный синтез и т. п.)» двигательных синергий (содружественность движений обоих глаз и г. и,), полисенсорности и многоуровневости организации процессов управления. Соответственно любые детерминанты движений глаз оказываются опосредствованными состоянием функциональной системы фиксационного поворота в целом. При необходимости наблюдатель способен усилить эффект одной из детерминант, ослабляя действие остальных.
↑ Механизм стабилизации взора
Целенаправленный поворот глаз завершается фиксацией наблюдателем значимых элементов или отношений среды. Речь идет о краткосрочной (200-400 мс) стабилизации зрительной направленности (взора), которая проявляется в соответствующей (а) направленности внимания, (б) ориентации оптических осей глаз и (в) установке, или состоянии готовности наблюдателя. Опосредованные установкой зрительно-аттенциональный и окуломоторный компоненты взора оказываются относительно независимыми.
Стабилизация взора предполагает участие всех звеньев функциональной системы окуломоторного акта. На основании афферентного синтеза задаются предмет (область) фиксации и критерии ее адекватности, складываются соответствующие установка и программа окуломоторной активности наблюдателя, наконец, актуализируются собственно двигательные средства: дрейф и микросаккады, контролируемые обратными связями (зрительной, вестибулярной, проприоцептивной). По существу, фиксация предмета означает окуломоторное отслеживание его зрительного направления.
При описании механизма стабилизации взора исследователи нередко обращаются к ретинотопической модели следящей системы. Предполагается, что во время фиксации дрейф, отражающий нестабильность экстраокулярных мышц, уводит изображение неподвижного предмета от fovea centralis (оптическая ось глаза уходит в сторону от центра фиксируемого объекта, в то время как зрительное направление последнего остается неизменным); благодаря обратной связи позиционное рассогласование регистрируется зрительной системой и инициирует коррекционную микросаккаду, возвращающую линию взора в центр предмета восприятия (Глезер, 1959; Глезер, Леушина, 1975; Cornsweet, 1956; Nachmias, 1959; Boyce, 1967). Данная модель построена на допущении неуправляемости дрейфа (случайности его направления) и опирается на ряд экспериментально установленных фактов; 1) зависимость вероятности появления микросаккад от амплитуды смещения ретинального изображения; 2) наличие «зоны нечувствительности» сетчатки; 3) соответствие амплитуды микросаккад диаметру «зоны нечувствительности» и др. Накопленный массив экспериментальных данных показывает, однако, что предлагаемое описание — лишь первое приближение к действительности.
Фиксационный дрейф. Несмотря на то, что дрейф является выражением собственных «шумов» ГДС, а также «люфта» взаимодействующих систем (окуломоторной и вестибулярной, правой и левой образующих вергентной субсистемы и т. п.), он подвержен влиянию различных переменных , а значит, управляем.
Скорость дрейфа, или скольжения глаза, увеличивается с увеличением расстояния до зрительного стимула (до 6°), его яркости и относительного контраста (Андреева, Вергилес, Ломов, 1975) (Рис. 3.3).
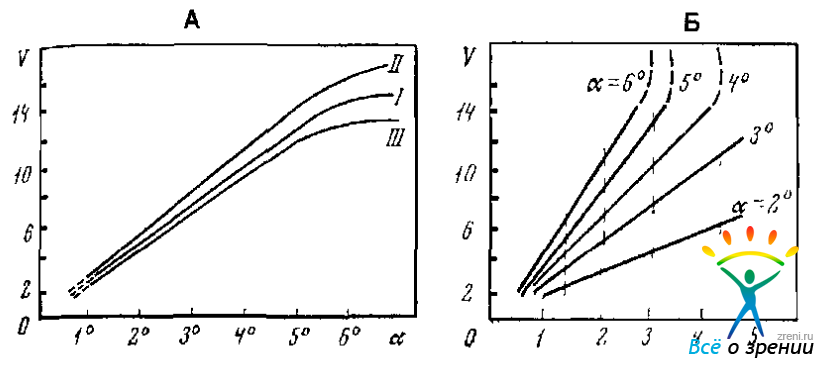
Рис. 3.3. Зависимость скорости «скольжения» глаза ( в угл. град./с) от рассогласования направлений глаз и стимульного объекта (А) и относительной яркости стимула (Б) (Андреева, Вергилес, Ломов, 1975).
При ограничении поля зрения до 1° зрительные оценки, пересчет объектов, восприятие и опознание контурных изображений осуществляются посредством плавных Движений, а не саккад (Зинченко, Вергилес, 1969; Андреева, 1972; Андреева, Вергилес, Ломов, 1975). В условиях «ганцфельда» (равноосвещенного незаполненного пространства) и стабилизации ретинального образа относительно сетчатки скорость и амплитуда дрейфа возрастают примерно на порядок. Во время наблюдения последовательных образов ускоренный дрейф направлен в сторону объекта фиксации и чувствителен к его размеру (площади) и локализации в поле зрения. В определенных ситуациях скользящие движения глаз могут принимать вид периодических колебаний (Белопольский, 1989; Doesschate, 1954). Рост скорости и амплитуды дрейфа наблюдается и с уменьшением коэффициента зрительной обратной связи ГДС; чем дальше от объекта фиксации удалена оптическая ось глаза, тем выше значения параметров дрейфа (нередко эта зависимость имеет вид экспоненты) (Белопольский, Вергилес, 1979; Вергилес, Андреева, 1990).
Предъявление точечного объекта на фоне медленно движущихся (десятки угл. сек./с) черно-белых полос допускает как сохранение фиксационной направленности глаза, так и его дрейф в сторону движения полос, но с гораздо меньшей скоростью. Возможность произвольного выбора той или иной стратегии окуломоторной активности сохраняется при сверхмедленной скорости смещения полос—пять угл. сек./с (Murphy, Kowler, Steinman, 1975). Если объект фиксируется на фоне быстро движущихся (десятки угл. град./с) черно-белых полос, дрейф получает соответствующую направленность, «растягивается») (возрастают его скорость и амплитуда) и становится плавным компонентом фиксационного оптокинетического нистагма—ФОКН (Гиппенрейтер, Романов, 1970; Stark, 1971). Гальванический нистагм, возникающий при фиксации объекта во время стимуляции вестибулярной системы электрическим током, напротив, «свертывается» (скорость его медленной фазы уменьшается в 10-15 раз), превращаясь в гальванический микронистагм—ГМН (Дормашев, Романов, 1983). Обе разновидности фиксационного нистагма зависят от физических характеристик объекта (яркости, размера и др.), степени сложности задачи и напряженности зрительного внимания. В частности, расширение площади настройки зрительного внимания на реальный или визуализированный предмет от 0,5 до 6° сопровождается значительным увеличением амплитуды нистагма и увеличением скорости его дрейфового компонента. Продолжительность отдельных дрейфов ФОКН и ГМН совпадают с выполнением и произвольным контролем микроэтапов текущего действия (Гиппенрейтер, 1978; Гиппенрейтер, Романов, 1990).
Требование зрительной фиксации неподвижного точечного объекта сопровождается увеличением, а требование сохранения взора в определенном положении—уменьшением частоты микросаккад (Steinman, Haddad, Skavenski, Wyman, 1973). Число микросаккад сокращается и во время активной интеллектуальной работы, в частности, при решении сложных арифметических задач (Филин, Сидоров, 1972), и с увеличением углового размера фиксируемого объекта (Steinman, 1976). В последнем случае амплитуда дрейфов также возрастает. Опытные испытуемые способны произвольно подавлять микросаккады, причем целевой стимул может располагаться в стороне от центральной точки фиксации (Филин, 1975; Steinman, 1976). В принципе, дрейф оптических осей достаточен как для достижения точной фиксации миниатюрного элемента среды, так и для дестабилизации ретинального образа (Ярбус, 1965; Ditchburn, 1973). Стабильность параметров дрейфа при нарушении проводимости глазодвигательного нерва (офтальмоплегии) демонстрирует его более тесную связь с эфферентными влияниями, чем с изменением состояния экстраокулярных мышц (Филин, 1975).
Дрейф не только отклоняет оптические оси глаз от центра тяжести поверхности объекта, но и приближает к нему, реализуя коррекционные функции. В свою очередь, микросаккады способны не только возвращать ретинальный образ в fovea centralis, но и выводить его из этой области (Шахнович, 1974; Nachmias, 1959, 1961). У разных испытуемых соотношение направлений дрейфов и саккад широко варьирует, конституируя тот или иной индивидуальный тип зрительной фиксации (Шахнович, 1974).
Микросаккады. Объем коррекционных фазических движений не превышает трети всех выполняемых микросаккад (Проскурякова, Шахнович, 1968; Воусе, 1967). Большая их часть обеспечивает обследование (сканирование) поверхностей миниатюрных объектов, особенно пограничных областей. В этой функции микросаккады тождественны макросаккадам и, вероятно, являются их редуцированной формой. Подобные движения глаз имеют место, например, при восприятии игольного ушка, в которое пытаются вдеть нитку.
В пользу данного предположения говорят факты совпадения средней частоты микросаккад во время фиксации отдельной буквы и макросаккад при чтении текста (Cunitz, Steinman, 1969), постоянство отношения амплитудно-скоростных характеристик саккад разных уровней (Zuber, Stark, Cook, 1965), а также возможность последовательного уменьшения («свертывания») амплитуды саккад в процессе зрительного научения (Steinman, Haddad, Skavenski, Wiman,1973). Существенно, что наблюдатели могут произвольно выполнять саккады амплитудой в несколько угловых минут, в том числе и в направлении, противоположном расположению предмета восприятия (это говорит о том, что для выполнения фазических движений необходим не столько сигнал рассогласования, сколько структурирование зрительного поля, которое задает соответствующий масштаб оптической системы координат; если при выполнении саккады предмет исчезает, амплитуда произвольных фазических движений возрастает в 3-4 раза (Haddad, Steinman, 1973)), осознают момент выполнения микросаккад и достаточно точно отслеживают микроступенчатые стимулы (Рис. 3.4)

Рис. 3.4. Движения глаз при предъявлении пары точек и инструкции «переводить взгляд с точки на точку» (Гиппенрейтер, 1978). А— расстояние между точками 2'; В — расстояние между точками 5'; В — расстояние между точками 8'. По обеим сторонам записей отметки испытуемого о переводе взора в соответствующую сторону.
(Гиппенрейтер, 1973, 1978; Haddad, Steinman, 1973). На «центральное» происхождение фазических микродвижений указывает их наличие у слепых и зрячих в полной темноте (Проскурякова, Шахнович, 1968; Филин, 1975).
В процессе фиксации сканирующие микросаккады (иногда их называют фликами) организуются в более или менее сложные структуры—«автономные фликовые комплексы» (Карпов, Карпова, 1978). Как правило, первый либо второй скачек, входящий в комплекс, уводит взор в сторону, а последующие—возвращают его в центр фиксируемого объекта; на временной развертке эта разновидность микродвиженнй имеет выраженную П-образную форму (Карпов, Карпова, Зеленкин, 1982; Feldon, Langston, 1977; Herishanu, Sharpe, 1981). Момент появления и направление фликов не зависят от направления предшествующего дрейфа. Частоты микросаккад и сканирующих макродвижений глаз примерно совпадают и изменяются в сходных обстоятельствах. Так, усиление познавательной активности (состояние напряженности, гиперактивация) ведет к увеличению, а ее снижение (состояние релаксации, отрешенности)—к снижению частоты саккад (Карпов, 1975; Карпов, Карпова, 1978). При выполнении задачи воспроизведения материала, предъявленного «на слух», фликовые комплексы соответствуют моментам ошибочного или неуверенного воспроизведения (Дормашев, Романов, 1989).
Несмотря на сходство параметров и условий появления, полное, или абсолютное тождество микро- и макросаккад отсутствует. Микросаккады более независимы от характеристик стимуляции (формы, цвета, размера фиксируемого элемента среды), не подчиняются действию «центра тяжести» миниатюрного контурного объекта и более «привязаны» к центральной точке фиксации. Поэтому, например, цепочки однонаправленных саккад, характерные для чтения текста или рассматривания изображений, на микроуровне встречаются редко (Steinman, 1965; Boyce, 1967; Ruttle, 1969; Murphy, Haddad, Steinman, 1974).
Зоны нечувствительности ГДС. ГДС отвечает далеко не на все оптические изменения среды. Область смещений стимула (около нулевого значения), не влияющих на окуломоторную активность, получила название «мертвого пространства» (Fender, Nye, 1961) или «зоны нечувствительности» сетчатки (Глезер, 1959). Согласно С. Rashbass (1961), саккадическая ветвь ГДС не способна отвечать на смещение точечного источника света меньше 25-50 угл. мин. H.Bennet-Clark (1964) указывает несколько меньшую величину—15-30 угл. мин., добавляя, что некоторые испытуемые реагируют на смещение цели менее 10 Угл.мин. По данным В. Д. Глезера, зона нечувствительности соответствует размеру рецептивного поля в центре fovea—около 5 угл.мин. (Глезер, 1959; Глезер, Леушина, 1975), но в принципе, может быть и менее 2 угл. мин. (Timberlake, Wyman, Skavenski, Steinman, 1972). По крайней мере, в 65% случаев ступенчатое смещение стимула на 1,7 угл. мин. вызывает саккаду соответствующей амплитуды; данный эффект не зависит от опыта и предварительной практики испытуемого (Wyman, Steinman, 1973).
Широкий разброс значений казалось бы одного и того же явления позволяет предположить, что зона нечувствительности носит функциональный характер и имеет переменный размер. Определяющими в данном случае становятся не столько анатомо-физиологические свойства ГДС, сколько способ схематизации пространства и установка наблюдателя, его готовность отслеживать микросмещения стимула. В зависимости от поставленной задачи и индивидуальных особенностей наблюдатель настраивается на определенный пространственно-временной масштаб восприятия элементов и отношений среды, пропуская изменения стимула меньшие функционально необходимых. Чем крупнее избираемый масштаб, тем шире зона нечувствительности. Данное предположение объясняет возможность переноса структуры «мертвого пространства» на уровень макродвижений глаз.
Выражением нечувствительности ГДС к непроизвольному смещению оптических осей является порог «позиционного чувства» глаза, или зона блуждания взора во время фиксации (Cornsweet, 1956). Ее величина не выходит за пределы 1 угл. град., варьируя в зависимости от стимульных условий, решаемой наблюдателем задачи, его состояния и других обстоятельств. В частности, зона блуждания взора расширяется с увеличением продолжительности фиксации, в условиях безориентирного поля зрения, при стабилизации изображения фиксируемого объекта относительно сетчатки, в результате поражения центрального и/или цветового зрения (Ярбус,1965; Шахнович, 1974; Ditchburn, 1973). Существенно, что непроизвольные микродвижения глаз могут не только содействовать, но и противостоять выполнению произвольных окуломоторных актов, т. е. выступают в роли «собственных шумов» ГДС. При исключении или нарушении зрительной афферентации их влияние на окуломоторную систему становится особенно наглядным. Согласно данным Ю. Б. Гиппенрейтер (1964; 1978), порог позиционного (мышечного) чувства глаз в условиях выполнения зрительной задачи имеет величину около 40 угл. мин.
Безориентирное поле зрения и элиминация оптической стимуляции. При исчезновении стимула в безориентирном поле зрения или при попытке фиксировать объект в темноте амплитуда и скорость дрейфа возрастают в 4-6 раз, а параметры незрительных видов нистагма (калорического, гальванического и др.)—более чем на порядок. Усиливается коррекционная зависимость между дрейфом и микросаккадами, причем частота фазических движений и их корректирующие возможности уменьшаются (Шахнович, 1974; Гиппеирейтер, 1978; Курашвили, Бабияк, 1975; Matin, Matin, Pearce, 1970; Skavenski, Steinman, 1970; Ditchburn, 1973).
В этих условиях натуральные функции дрейфа (стабилизация взора, дезадаптация сетчатки) остаются нереализованными, а сам он выражает лишь «шум» окуломоторной системы (Skavenski, Steinman, 1970). Ведущими афферентаторами ГДС становятся органы равновесия. Дисбаланс сигналов, поступающих от правого и левого отолитовых аппаратов, инициирует плавные смещения глаз в каком-либо одном из направлений. Именно в темноте тонические микродвижения правого и левого глаза становятся асимметричными (Skavenski, Steinman, 1970; Haddad, Winterson, 1975), а «зона нечувствительности» увеличивается до 2-3 угл. град. (Becker, 1976).
Полученные экспериментальные данные указывают не только на зависимость микродвижений глаз от зрительной афферентации, но и на определяющую роль задачи: требование сохранить положение глаз в заданном направлении явно отлично от требования фиксации видимого элемента среды. В противоположность решению зрительной задачи, сохранив позиции глаза в темноте предполагает активное участие проприоцептивной и вестибулярной афферентации, с одной стороны, и схемы окружающего индивида пространства, соотнесенной со схемой его тела,—с другой. Сохранение направленности глаз в темноте—всегда усилие (Matin, Matin, Pearce, 1970), или волевое действие субъекта, в какой-то степени компенсирующее отсутствие тонкого зрительного механизма стабилизации взора (Шахнович, 1974).
При попытке сохранить эксцентрическое направление взора в темноте глаза дрейфуют в сторону позиции покоя. Общая закономерность состоит в том, что чем больше отклонен взор, тем выше скорость тонических движений. Максимальная скорость Дрейфа, соответствующая эксцентриситету в 35°, достигает 10 Угл. град/с. Частично плавные смещения глаз компенсируются возвратными, или коррекционными, саккадами, оформляясь в пилообразные периодические колебания—«нистагм конечной позиции» (Blomberg, 1958). Появление коррекционных саккад в темноте указывает на надмодальный (незрительный) характер репрезентации фиксируемого объекта и возможность использования экстраретинальной обратной связи в регуляции движений глаз. Большие (свыше 5°) отклонения глаз от заданного направления осознаются и корректируются произвольно (Becker, Fuch 1969; Becker, Klein, 1973; Barns, Gresty, 1973).
Движения закрытых глаз, сопровождающих целенаправленную деятельность человека, варьируют в диапазоне ±15° и состоят из ускоренных однонаправленных макродрейфов и возвратных саккад. Продолжительность дрейфа коррелирует с продолжительностью усилия (умственной работой) испытуемого. Во время измененных состояний сознания, которые снимают направленную активность испытуемого, саккады подавляются, а доминирующей формой окуломоторной активности становятся плавные периодические и апериодические колебания (вращения) глаз, в несколько раз превышающие амплитуду и скорость обычного дрейфа (Вуякас, Михеев, Пономаренко, 1985; Вуякас, Михеев, 1987).
Освещение демонстрируемых объектов стробоскопическим светом оказывает влияние преимущественно на дрейфовый компонент окуломоторной активности. Во время фиксации центра кольца при инструкции подавления саккад зрительные оси смещаются в сторону, совершая, одновременно синусоидальные колебания, синхронизированные с частотой вспышек света (латентный период 130 мс). Амплитуда колебаний уменьшается с увеличением частоты вспышек от 20 угл. мин. на частоте 0,5 Гц до 6 угл. мин.—на частоте 5 Гц. На частотах свыше 10 Гц колебания исчезают, но некомпенсированный дрейф, амплитудой в несколько угл. град., сохраняется (Haddad, Winterson, 1975). Если задача подавления микросаккад не ставится, стробоскопическое освещение (0,5-2 Гц) может приводить к увеличению фазических движений (West, Boyce, 1967). Стробоскопическая демонстрация объекта создает, следовательно, промежуточные условия организации движений на свету и в темноте, обнажая рассогласование разнонаправленных (по способу влияния) детерминант—зрительной (ретинальной) и незрительной (экстраретинальной) природы.
Таким образом, ретинотопическая модель следящей системы в лучшем случае описывает наиболее общие принципы сохранения (стабилизации) направленности взора: 1) предварительная заданность определенной направленности глаз; 2) существование отрицательной обратной связи окуломоторной системы и 3) наличие порога смещения направленности глаз. Главный постулат модели: предположение о неуправляемости дрейфа, случайности его направления экспериментально не подтверждается. Не подтверждается и представление о рассогласовании между локализацией ретинального образа и fovea centralis как стимуле, запускающем микросаккады в направлении объекта фиксации. По-видимому, ни дрейф, ни микросаккады не несут жестко специализированных функций, а их биомеханические возможности реализуются в зависимости от конкретного сочетания внешних и внутренних условий (детерминант) окуломоторного акта.
Как мы убедились, параметры дрейфа (скорость, направление, амплитуда) тесно связаны с параметрами оптической стимуляции (яркостью и контрастностью объектов, их ориентацией, структурой, величиной, скоростью перемещения, способом освещения, размером поля зрения), требованиями решаемой задачи (типом—интеллектуальная/зрительная/окуломоторная, содержанием, сложностью), характеристиками внимания (площадью настройки, устойчивостью, напряженностью), коэффициентом зрительной обратной связи ГДС, влиянием интермодальных взаимодействий, индивидуальными особенностями наблюдателей и др. Поэтому дрейф не только отклоняет оптическую ось глаз от заданной ориентации, но и содействует ее сохранению. Сходные обстоятельства (детерминанты) определяют характеристики микросаккад (их амплитуду, направление, частоту), которые наряду с коррекцией направления оптической оси глаз осуществляют сканирование миниатюрного объекта. Организуясь в самостоятельные структуры, микросаккады и дрейф обеспечивают относительную стабилизацию взора наблюдателя и подчинены гибкой системе детерминант. В число последних входят как зрительные, так и незрительные (экстраретинальные, интермодальные н др.) составляющие, действие которых нередко носит альтернативный характер.
Зрительно-окуломоторные отношения, складывающиеся во время фиксации выступают как многоуровневые, многократно oпосредствованные и носят скорее функциональный, а не морфологический характер. Совершенно необязательно, чтобы фиксируемый предмет проецировался именно в область fovea centralis, как необязательно совпадение «зоны нечувствительности» с величиной рецептивного поля в центре fovea или наличия освещенного объекта вообще. Приемлемая освещенность зрительного поля, анизотропность сетчатки и минимальный размер ее рецептивного поля создают необходимые, но недостаточные условия сохранения направленности взора. Последние определяются содержанием взаимосвязи субъекта восприятия с объектом, той задачей, которая выполняется наблюдателем. В зависимости от ее требований складывается готовность воспринимать определенные пространственные и временные отношения среды ( как бы устанавливается масштаб восприятия предметов некоторого класса), которая формирует соответствующий ей окуломоторный «центр тяжести» и «зону нечувствительности»—пространство допустимых отклонений направления глаз во время фиксаций. При этом особенно важен тип задачи: является ли она собственно зрительной, окуломоторной или интеллектуальной, а также участие волевого компонента в процессе ее решения. Именно функциональность зрительно-окуломоторных отношений объясняет тот факт, что при усложнении или затруднении условий фиксации дрейф развивает скорость и амплитуду диапазона прослеживающих движений (даже в отсутствие движущихся стимулов), а микроскачки «растягиваются» до параметров обычных саккад.
----
Статья из книги: Окуломоторные структуры восприятия | Барабанщиков В.А.

Комментариев 0