Наружные мышцы глаза │ часть 2


Содержание:
Описание
↑ Аномалии наружных мышц глаза
В глазнице иногда обнаруживаются так называемые аномальные мышцы. К ним относятся:
- Gracilis orbitis исходит из проксимальнодорсальной поверхности верхней косой мышцы. Прикрепляется она к блоку или к окружающей блок соединительной ткани. Эта мышца иннервируется блоковым нервом.
- Добавочная мышца наружной прямой мышцы иногда обнаруживается у обезьян. Иннервируется она отводящим нервом.
Иногда аномальные мышцы сопутствуют леватору верхнего века.
- Мышца, напрягающая блок, отделяется от внутренней поверхности леватора и прикрепляется к блоку или вблизи него.
- Поперечная мышца глазницы (т. transversus orbitalis) располагается между внутренней и наружной стенками глазницы и соединяется с леватором по пути своего следования.
Обе мышцы иннервируются верхней ветвью глазодвигательного нерва.
↑ Функциональная анатомия наружных мышц глаза
Наружные мышцы глазного яблока в гистологическом отношении относятся к поперечнополосатым мышцам. Этим они принципиально отличаются от гладких внутриглазных мышц (дилятатор и сфинктер радужной оболочки и ресничного тела).
Подробно структурная организация скелетной поперечнополосатой мышечной ткани приведена в 1-й главе. Мы рекомендуем читателю обратиться к ней. Это значительно упростит восприятие излагаемых в настоящем разделе сведений.
Необходимо сразу подчеркнуть, что основной задачей раздела, посвященного функциональной анатомии наружных мышц глаза, является довольно подробное описание отличительных особенностей наружных мышц глаза.
Особенности микроскопического строения наружных мышц глаза изучаются давно и интенсивно до сих пор. Полученные к настоящему моменту данные довольно противоречивы. Это связано как с наличием существенных межвидовых различий строения мышц, так и с возникающими сложностями при попытке сопоставления особенностей их структурной организации с функцией.
При микроскопическом исследовании сразу обращает на себя внимание то, что они не столь интенсивно оплетены плотной соединительной тканью, как другие скелетные мышцы. При этом в соединительнотканной оболочке обнаруживается много эластических волокон и необычно большое количество нервных стволов. Об этом свидетельствует хотя бы тот факт, что в скелетных мышцах одно нервное волокно приходится на 100 и более мышечных волокон, а в наружных мышцах глаза это отношение равняется приблизительно 1:5 — 1:10.
Многие исследователи выявили и другие различия. Волокна наружных мышц тоньше, чем волокна скелетных мышц. Причем их толщина постепенно увеличивается, как в эмбриональном периоде, так и в постнатальной жизни (табл. 2.8.3).

Таблица 2.8.3. Средний диаметр мышечных волокон наружных мышц глаза человека
Время сокращения мышечных волокон наружных мышц глаза значительно короче, чем сокращения скелетной мышцы, а амплитуда сокращения незначительная, на что обратил внимание еще в 1904 г. Schiefferdecker.
Для наружных мышц глаза характерно наличие так называемых «тонических» волокон, которые в скелетных мышцах вообще не обнаруживаются. Близкие по строению волокна выявляются лишь в мышце, напрягающей барабанную перепонку человека, и скелетной мускулатуре амфибий и птиц. Иннервируются такие волокна многочисленными нервными окончаниями, беспорядочно распределенными по всей поверхности мышцы.
Отличаются мышцы глаза и тем, что в отличие от скелетных мышц длина их мышечных волокон различная. Некоторые волокна короче длины всей мышцы.
Расположение мышечных волокон мышц глаза менее плотное. Именно по этой причине на поперечном срезе мышечные волокна мышц глаза округлые, а скелетной мышцы в виде многогранников.
Благодаря исследованиям Kato впервые было установлено, что, в отличие от скелетных мышц, в наружных мышцах глаза четко различаются две зоны. Если на момент рождения диаметр мышечных волокон одинаков, то с возрастом волокна, обращенные в сторону орбиты, становятся тоньше, а волокна, обращенные в сторону глазного яблока — толще. Именно по этой причине эти участки мышц были названы «орбитальной» и «глазной» зонами. Диаметр волокон «орбитальной» зоны колеблется между 5 и 15 мкм, в то время как диаметр «глазных» волокон 10—40 мкм. В пределах этих зон были идентифицированы различные подтипы мышечных волокон, отличающихся структурой, обменом веществ и сократимостью.
Каждое мышечное волокно наружных мышц глаза состоит из сарколеммы, окружающей зернистую саркоплазму, содержащую многочисленные миофибриллы диаметром 1—2 мкм. В целом строение мышечного волокна наружных мышц глаза соответствует строению волокна скелетной мышцы, подробно описанному в 1-й главе, но выявляются и различия, приведенные на рис. 2.8.14.

Рис. 2.8.14. Схема ультраструктурных особенностей наружных мышц глаза человека (по Martinez et al., 1976): 1— мультивезикулярные структуры; 2— включение, расположенное под сарколеммой; 3 — тельце Хирано; 4 — пластинчатое тело; 5 — капли липидов; 6 — гранулы гликогена; 7 — митохондрия; 8— Т-система; 9 — волокнистые палочки; 10— сарколемма; 11 — базальная мембрана; 12— лептофибриллы; 13— внутримышечный нерв; 14 — немиелинизированное нервное волокно; 15 — миелинизированное нервное волокно; 16 — шванновская клетка; 17 — тельце Люзе; 18 — коллагеновое волокно; 19 — капилляр; 20 — клетка сателлит
Самые ранние исследования выявили в наружных мышцы глаза как «темные» волокна, содержащие большое количество ядер и обильную саркоплазму, так и «светлые» волокна, отличающиеся небольшим количеством ядер и слабо выраженной саркоплазмой.
Thulin обнаружил, что мышечные волокна с обильной саркоплазмой («темные» волокна) отличались регулярным расположением миофибрилл. В связи с этим такие волокна он назвал fibrillenstruktur. «Светлые» волокна, наоборот, отличались неправильным расположением миофибрилл и получили название feiderstruktur. Эти немецкие термины трудно перевести на другие языки. По этой причине они используются до сих пор в англо-американской и французской литературе в немецкой транскрипции. Эта светооптическая классификация и в настоящее время является классификационной основой разделения волокон наружных мышц глаза на два типа.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Не только характером расположения микрофибрилл отличаются эти два типа волокон. Существенные различия выявляются и при анализе структуры миофибрилл, внутриклеточной организации, типе иннервации и др. (табл. 2.8.4).

Таблица 2.8.4. Структурные различия fibrillenstruktur и feiderstruktur волокон наружных мышц глаза
С целью классификации волокон наружных мышц глаза, помимо светооптических различий мышечных волокон, были использованы и другие методы —
- электронная микроскопия,
- гистохимия,
- иммуногистохимия.
Довольно четкие критерии выявлены при использовании электронной микроскопии. Наиболее надежные ультраструктурные критерии дифференциации волокон — учет размера и расположения миофибрилл, размер, количество и локализация митохондрий, степень развития саркоплазматической сети и Т-трубочек.
Мышечные клетки наружных мышц глаза возможно классифицировать и на основании их гистохимических особенностей. Гистохимический профиль мышечных волокон отражает особенности метаболизма и коррелирует с типом их сокращения.
Первоначально мышечные волокна классифицировали по их цвету («красные» и «белые»). Dubowitz и Pearse обнаружили при исследовании скелетных мышц, что цвет мышечных волокон связан с различной активностью в них фосфорилазы и окислительных ферментов, а также концентрации миоглобина.
На основании гистохимического профиля также можно четко выделить два типа волокон. Волокна малого диаметра отличаются высокой активностью митохондриальных окислительных ферментов и фосфорилазы, а в волокнах большого диаметра активность ферментов низкая. Первый тип волокон, таким образом, в энергетическом обмене использует цикл Кребса, а второй — гликолиз.
Последующие исследования выявили существование различий в активности миозин-аденозин-трансферазы, дифосфоридин-нуклеотид дегидразы и других ферментов, (см. табл. 2.8.5).

Таблица 2.8.5. Гистохимический профиль различного типа волокон наружных мышц глаза (пo F. Spencer, J. D. Porter, 1988)
Значение в классификации волокон имеет также учет активности специфического фермента поперечнополосатой мышечной ткани, а именно АТФ-азы. Ферментативной активностью обладает сам белок миозин. Наибольшую АТФазную активность миозины различной изоформы могут проявлять в кислой или щелочной среде. Именно это свойство применяют в дифференциации различных типов мышечных волокон наружных мышц глаза (табл. 2.8.5, 2.8.6).
Большинство исследователей на основе гистохимических различий между мышечными волокнами выделяют до 6 типов волокон (табл. 2.8.5).
Современные классификации мышечных волокон являются комплексными и основаны на одновременном учете данных различных методов. На одной из них мы остановимся.
↑ Классификация волокон наружных мышц глаза
Asmussen et al.,, Spencer, Porter, Porter et al. создали довольно четкую классификацию мышечных волокон наружных мышц глаза. В ней использованы морфологические (включая ультраструктурные) и гистохимические различия между мышечными волокнами. При этом наиболее существенны различия между волокнами, расположенными в «орбитальной» или «глазной» зонах. Именно эту классификацию мы и приводим подробно в табл. 2.8.5.
«Орбитальная» зона. В «орбитальной» зоне различают 2 типа волокон.
Волокна 1-го типа. В «орбитальной» зоне преобладают небольшого диаметра мышечные волокна (80%). Эти волокна обладают обильной саркоплазматической сетью, развитой Т-системой. Миофибриллы располагаются строго организованным образом (fibrillenstmktur), и в них выявляются многочисленные собранные в группы митохондрии. Отдельные волокна окружены густой сетью капилляров.
При гистохимическом исследовании выявляется, что волокна 1-го типа богаты окислительными ферментами, но при этом способны и к анаэробному обмену веществ. Наиболее близки эти волокна к скелетным волокнам типа II.
Необходимо отметить, что состав изоформ миозина изменяется по длине волокна. Так, в средней части мышцы преобладает «быстрый» миозин, а в других участках мышцы обнаруживаются сразу несколько различных типов миозина. К таковым относятся миозин, характерный для волокон типа ПА скелетной мышцы, зародышевый миозин и миозин, определяемый только в наружной мышце глаза.
Тем не менее обнаруживаются и отличия от скелетных волокон типа НА. Волокна 1-го типа наружных мышц глаза характеризуются высокой степенью выносливости. Обладают они и уникальным, свойственным только им, миозином. Кроме того, каждое мышечное волокно иннервируется отдельным нервным окончанием, образующим сложные двигательные бляшки, буквально окружающие мышечные волокна. Насыщены эти волокна «спиральными» чувствительными окончаниями.
Волокна 2-го типа. В «орбитальной» зоне выявляются волокна 2-го типа, относящиеся к медленным волокнам. Характер сокращения волокна 2-го типа напоминает сокращение тонических волокон земноводных.
Волокна 2-го типа составляют около 20% всех волокон «орбитальной» зоны. Они отличаются высокой активностью миозин-АТФ-азы в кислой среде и менее выраженной активностью в щелочной среде. В этих волокнах миофибриллы распределены неравномерно (feiderstruktur) и напоминают быстрые волокна скелетных мышц (тип ПС).
Хотя миофибриллы 2-го типа иннервируются многочисленными нервными волокнами, они способны к быстрым сокращениям в центральных участках мышцы и характеризуется низкой скоростью сокращения в дистальных отделах мышцы. Это обеспечивается структурными различиями, выявляемыми по длине мышцы.
«Глазная» зона. В «глазной» зоне существует три типа волокон. Все они относятся к быстрым волокнам и подобны скелетным волокнам типа II.
Волокна 3-го типа. Мышечные волокна этого типа относятся к «красным» волокнам. Иннервируются они отдельным нервным волокном. Количество подобных волокон достигает 30% всех волокон «глазной» зоны.
Для волокон 3-го типа характерно проявление высокой активности окислительных ферментов. Выражен также гликолитеческий обмен. По этим характеристикам волокна 3-го типа можно отнести к выносливым волокнам.
По всей длине волокна определяется только одна изоформа миозина, напоминающая миозин скелетного волокна типа ПА.
Волокна 4-го типа иннервируются отдельным нервным волокном и составляют до 25% волокон «глазного» слоя.
В соответствии с ультраструктурными характеристиками и активностью АТФ-азы эти волокна относятся к быстрым волокнам. Миозин напоминает миозин скелетных волокон типа ИВ.
Волокна 4-го типа проявляют умеренную активность окислительно-восстановительных и аэробных ферментов. Саркоплазма волокон насыщена мелкими митохондриями.
Волокна 5-го типа иннервируются отдельным нервным волокном и составляют 30% волокон «глазной» зоны наружных мышц. Напоминают они скелетные волокна типа ИВ. Волокна проявляют низкую активность окислительных ферментов. При этом хорошо выражен анаэробный метаболизм. Активность миозин-АТФ-азы характерна для быстрых волокон. Между миофибриллами располагаются мелкие митохондрии.
Приведенные структурно-функциональные особенности этих волокон свидетельствуют о возможности их отнесения к быстрым волокнам с невысокой выносливостью.
Волокна 6-го типа относятся к медленным и напоминают тонические волокна амфибий. Составляют они 10% волокон «глазной» зоны. Ультраструктурно выявлено, что они относятся к «Ге1с1ег5{гик1иг»-волокнам с очень большим диаметром миофибрилл. Волокна 6-го типа иннервируются многочисленными «гроздеподобными» нервными окончаниями, равномерно распределенными по всей длине мышцы.
Отличаются эти миофиламенты и наличием различных изоформ миозина. Для них характерен миозин, свойственный медленным волокнам (тип I), а также альфа-миозин сердечной мышцы.
Существенное уточнение классификации мышечных волокон наружных мышц глаза, как уже можно было заметить, произошло в результате применения иммунопктохимических методов, позволяющих определять различные изоформы миозина.
На основании проведенных исследований выявлено десять изоформ тяжелого миозина. Каждая форма отличается сократимостью, а их синтез контролируется различными генами. Классификация мышечных волокон, основанная на наличии в них различных подтипов миозина Rowlerson, приведена в табл. 2.8.4. Мы кратко остановимся на ней.
«Орбитальная» зона. Много волокон орбитальной зоны содержат миозины зародышевого типа. Соотношение зародышевого миозина снижается с возрастом, но он не исчезают полностью. Маленький диаметр и менее организованную ультраструктуру этих волокон некоторые авторы связывают с незрелостью миофибрилл. Предполагают, что сохраняются незрелые миозины при пониженной нагрузке на наружные мышцы глаза.
Использование антител против миозинов медленных волокон позволяет выявить еще два других типа волокон — медленные и «тонические» волокна.
«Глазная» зона. Большая часть волокон наружной мышцы глаза содержит миозин, обнаруживаемый только в наружных мышцах глаза. Волокна отличаются активностью АТФ-азы. характерной для семейства быстрых миозинов. При этом перекрестной реакции с миозинами скелетной мышцы нет. Этот тип миозина преобладает в «глазной» переходной зоне, а также обнаруживается в волокнах «орбитальной» зоны.
Другой тип волокон «глазной» зоны, выявляемый особенностями изоформы миозина, относится к медленным волокнам («тонические» волокна земноводных). Тем не менее эти волокна иннервируются многочисленными нервными волокнами, а гистохимически и ультрастурктурно напоминают скелетные мышечные волокна 1-го типа.
↑ Особенности двигательной иннервации различных типов мышечных волокон
Иннервация быстрых волокон. Быстрые волокна иннервируются ветвями одиночных аксонов двигательных нервов. При этом иннервируется небольшое число мышечных волокон. Заканчиваются аксоны двигательной бляшкой (нервно-мышечный синапс) (рис. 2.8.16),
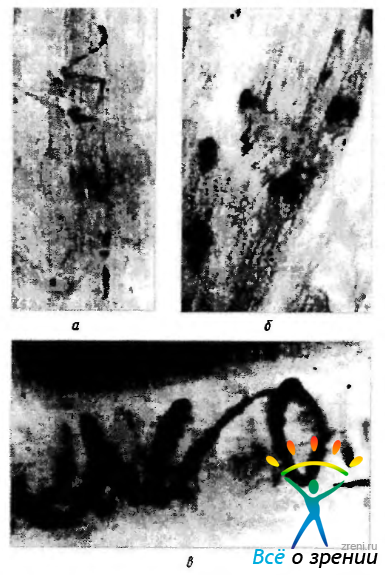
Рис. 2.8.16. Светооптические особенности двигательных нейромышечных соединений наружных мышц глаза (no Cheng, 1963): а - двигательная бляшка; б - «гроздевидные» нейромышечные соединения; в — спиралевидное окончание
по строению напоминающей двигательные бляшки скелетной мышцы. Синаптическая мембрана обладает многочисленными складками, а терминальная часть аксонов насыщена синаптическими пузырьками, содержащими холинэстеразу.
Двигательные бляшки распределеляются в области дистальной средней трети брюшка мышцы шириной 1—2 мм.
При морфометрическом исследовании диаметра аксонов двигательных нервов, подходящих к наружным мышам глаза, выявлено бимодальное распределение волокон с пиками в области 5 мкм и 11 мкм. Аксоны большего диаметра иннервируют быстрые волокна, а маленького диаметра иннервируют «тонические» волокна.
Наружные мышцы глаза являются самыми быстрыми в организме человека (например, 7,5—10 мс для внутренней прямой мышцы по сравнению с 40 мс для gastrocnemius). Частота раздражения, необходимая для стимуляции слияния ответов мышцы, равняется 350— 450 Гц, а для gastrocnemius составляет всего 100 Гц. Скорость восстановления наружных мышц глаза приблизительно в десять раз выше.
Иннервация «тонических» волокон. «Тонические» волокна иннервируются многочисленными «гроздевидными» окончаниями (рис. 2.8.17).

Рис. 2.8.17. Особенности двигательных нервных окончаний наружных мышц глаза (по Locket, 1978): а — двигательное окончание в прямой мышце глаза (серебрение по Бильшовскому); б схема двигательных окончаний мышечного волокна малого диаметра; в—двигательное волокно мышечного волокна большого диаметра (1 — нервное волокно (миелиновая оболочка исчезает при вхождении волокна в мышцу); 2 — нервные окончания, оплетающие ядра мышечных волокон; 3 - поперечная исчерченность мышечного волокна)
Их двигательные волокна меньшего диаметра, чем диаметр нервных волокон, иннервирующих быстрые мышечные волокона. Выявляются и структурные отличия двигательных бляшек. Это, в первую очередь, относится к строению синаптических пузырьков. Синаптические пузырьки содержат зернистое содержимое, напоминающее таковое некоторых нервных окончаний вегетативной нервной системы. Правда, необходимо отметить, что обнаруживаются и синаптические пузырьки обычного строения.
Еще одним отличием является то, что между оболочкой нерва и сарколеммой нет базальной мембраны, обнаруживаемой в мышечном веретене и окончаниях вегетативной системы.
«Гроздевидные» окончания «тонических» волокон концентрируются преимущественно в дистальной трети мышцы, образуя при этом группы, расстояние между которыми колеблется от 10 мкм до 2—3 мм Mukuno описал шесть типов двигательных бляшек, обнаруживаемых в наружных мышцах глаза человека
Нервная стимуляция «тонических» мышечных волокон приводит к медленному градуированному их сокращению. Частота регенерации равняется 200 Гц. При стимуляции с частотой 400 Гц скорость сокращения не увеличивается.
В отличие от быстрых волокон в «тонических» волокнах после возбуждения происходит пассивное электротоническое распространение деполяризации поперек поверхности волокон. Потенциалы «гроздевидных» синапсов меньше, чем потенциалы двигательных бляшек, что связывается с меньшим размером постсинаптической области и меньшим количеством высвобождаемого медиатора.
↑ Сенсорный аппарат наружных мышц глаза
Ранее предполагали, что чувствительные волокна, берущие свое начало в наружных мышцах глаза, проходят к центральной нервной системе в составе глазодвигательного, блокового и отводящего нервов. При этом они завершаются в нейронах ядра среднемозгового пути тройничного нерва (n. mesencephalicus n. trigemini). В настоящее время считают, что в глазодвигательном нерве проходит незначительное количество центростремительных волокон. Рассечение глазного нерва обезьяны приводит к дегенерации лишь 0,9—2,7% аксонов нерва, идущих к нижней косой мышце.
Ряд исследователей зарегистрировали сенсорные ответы у некоторых животных (овцы, кролика) в дорзолатеральнои части тройничного ганглия. При этом выявлена соматотопическая зеркальная пространственная организация нервных ответов, соответствующая объемному расположению структур глазницы. Эти данные были подтверждены Porter, Guthrie, Sparks при исследовании кошек и обезьян.
Нейроны первого порядка заканчиваются в чувствительных ядрах тройничного нерва, а также и спинномозговых ядрах. Нейроны второго порядка проходят к покрышке и крыше, а также к вентролатеральному ядру зрительного бугра (n. uentrolateralis thalami).
Информацию эти волокна передают от следующих типов рецепторов наружных мышц глаза (рис. 2.8.18; табл. 2.8.5):
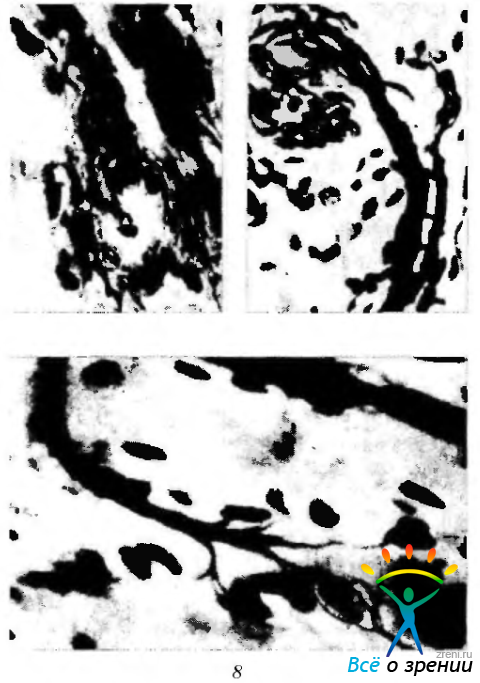
Рис. 2.8.18. Сенсорный аппарат наружных мышц глаза (па Wolter, 1955): а— щеткоподобные окончания; б—почкоподобные окончания; в — древоподобные окончания
- Мышечное веретено.
- Сухожильные органы Гольджи.
- Палисадоподобные окончания.
- Спиральные нервные окончания.
Мышечное веретено. Строение мышечного веретена скелетной мышцы описано в 1-й главе. Здесь мы лишь напомним о нем, а затем остановимся на особенностях этих образований в наружных мышцах глаза.
Мышечные веретена в наружных мышцах глаза человека впервые описали Siernmerling и Buzzard, но длительное время эти наблюдения не подтверждались. Лишь благодаря работам Daniel, Cooper, Daniel, эти рецепторы вновь обнаружены и описаны многими исследователями.
Мышечные веретена сконцентрированы вдали от средней трети брюшка мышцы, содержащей двигательные окончания. Наибольшее их количество в проксимальной (нижняя прямая мышца) и дистальной частях мышцы (верхняя косая мышца). Их можно обнаружить и в обеих зонах. Количество образований колеблется в одной мышце от 22 до 71. Blumer, Lukas, Aigner выявили следующее количество веретен в различных мышцах: во внутренней прямой 18,8 ±3,0; наружной прямой 19,3+ 1,9; в нижней прямой 34,0 ±4,4; в верхней косой 27,3 ±8,2; в нижней косой 4,3 ±1,8.
Мышечное веретено сообщает о скорости изменения длины мышцы. Оно представляет собой продолговатое тельце, окруженное соединительнотканной капсулой. Содержит мышечное веретено до 11 интрафузальных мышечных волокон небольшого диаметра (5—30 мкм) и длиной приблизительно 500 мкм. Размещены интрафузальные мышечные волокна параллельно экстрафузальным волокнам. Ядра интрафузальных волокон группируются двумя способами. В первом случае ядра лежат в центре, образуя ядерную сумку. Во втором случае они лежат линейно в виде цепочки.
На интрафузальных волокнах имеются два типа чувствительных центростремительных окончаний: первичные окончания иннервируют как волокна с ядерной цепочкой, так и волокна с ядерной сумкой (морфологически относятся к аннулоспиральным окончаниям).
Второй тип окончаний (афференты группы II типа) иннервируют исключительно волокна с ядерной цепочкой (рис. 2.8.18). Сокращение экстрафузальных мышечных волокон деформирует веретено, в результате чего и формируется нервный сигнал.
Афференты второго типа информируют центральную систему о длине волокна, а афференты первого типа — о длине и скорости изменения длины волокна. Гамма-волокна, направляющиеся к интрафузальным волокнам, управляют чувствительностью детектирования длины.
Fuchs и Komhuber предполагают, что мышечное веретено обеспечивает мозжечок информацией относительно размера и конечных точек саккадических движений глаза. Они придают им также роль в фиксационном нистагме. Sears et al. обнаружили доказательства того, что рецепторы растяжения мышцы подавляют двигательные нейроны наружных мышц глаза, расположенные в стволе мозга. Предполагают, что информация, идущая от мышечного веретена, исправляет положение глаза при «пролете» глаза мимо зрительной цели.
Сухожильные органы Гольджи. Сухожильные органы были описаны в скелетной мышце Golgi в 1880 г. Правда, он не обнаружил их в наружных мышцах глаза. В наружной мышце глаза человека эти чувствительные рецепторы описаны Marchi в 1882 г..
Сухожильные органы Гольджи информируют нервную систему о степени ригидности скелетных мышц. В мышцах эти рецепторы локализуются в сухожилии.
Сухожильные органы Гольджи в мышце глаза имеют веретеновидную форму и исключительно маленький размер. Средний размер этих образований в мышце обезьяны составляет 270x36 мкм, т.е. всего лишь третью часть аналогичных образований в скелетной мышце.
В скелетной мышце к сухожильному органу подсоединяются многочисленные мышечные волокна, а в мышцах глаза только одно или два. Кроме того, в скелетной мышце один конец сухожильного органа расположен в мышце, а другой в сухожилии. В противоположность этому в наружной мышце глаза человека все образование располагается в сухожилии.
Капсула сухожильного органа Гольджи образована периневрием, окруженным цитоплазматическими отростками фибробластов. Пучки коллагеновых волокон лежат обособленно. Между коллагеновыми волокнами распространяются нервные окончания.
Электронная микроскопия сухожильного органа Гольджи выявляет обычное распределение нервных окончаний, которые состоят из округлых нервных отростков, окруженных цитоплазмой шванновских клеток и выстланных пучками коллагеновых волокон. Плазматическая мембрана нервных окончаний иногда непосредственно прилежит к коллагеновым волокнам, а иногда между ними выявляется материал, напоминающий материал базальной мембраны. Между ними может находиться и цитоплазма шванновской клетки.
Палисадоподобные окончания. Палисадоподобные окончания (миосухожильные цилиндры) являются основным чувствительным аппаратом наружных мышц глаза млекопитающих. Они состоят из скрытых нервных окончаний, находящихся в месте перехода мышцы в сухожилие (мышечно-сухожильные), переплетенных с мышечными волокнами. Richmond, Johnston, Baker, используя импрегнацию серебром мышц человека, описали их в виде сети мелких нервных волокон, оплетающих кончик экстрафугального нервного волокна. Несколько соседних мышечных волокон иннервируются одним миелинизированным ветвящимся аксоном.
Чаще всего палисадообразные окончания обнаруживаются в горизонтальных прямых мышцах глаза. Несколько меньше их в вертикальных прямых мышцах и косых мышцах.
Сенсорная функция этих структур обеспечивается тем, что отсутствует базальная мембрана между мышцей и нервом, а также наличием близкого контакта нервного и мышечного волокон. Возбуждение окончаний, вероятно, происходит в результате сжатия окончаний при мышечном сокращении.
Спиральные нервные окончания. Эти окончания выявляются в средней трети наружных мышц глаза. Спиральные окончания обматывают экстрафузальное мышечное волокно.
В настоящее время считают, что спиральные окончания в мышцах глаза выполняют двигательную функцию.
Одиночные спиральные окончания редки. Они делают 3—8 витков вокруг мышечного волокна и заканчиваются в структуре подобно концевой пластинке.
Более часто можно обнаружить спиралевидные окончания в виде двух волокон, распространяющихся во встречном направлении. Описаны и другие варианты строения этих образований.
↑ Другие морфологические особенности наружных мышц глаза
В наружных мышцах глаза человека в норме обычно выявляются изменения, которые, если бы они были обнаружены в скелетной мышце, расценивались как патологические (миопатия или дегенерация). Сводятся они к обнаружению вихреподобного расположения миофибрилл, вакуолей между миофибриллярными пучками, различного типа включений, аномалий саркомеров, к смазыванию Z-линий и телец Хирано. Кроме того, выявляются разнообразие в форме и размере мышечных волокон, нарушение соотношения между волокнами различного диаметра, наличие большого количества ядер и присутствие мононуклеарного инфильтрата. Причина подобных изменений неизвестна.
Необходимо отметить, что при старении в наружных мышцах глаза отмечается потеря мышечных волокон 2-го типа, первоначально в «орбитальной», а затем и в «глазной» зоне. Позже уменьшается и количество волокон 1-го типа в «глазной» зоне.
↑ Особенности реакции наружных мышц глаза на действие фармакологических средств
Мышечные волокна наружных мышц глаза отличаются от скелетных мышц фармакологическими свойствами. Это, в первую очередь, касается действия миорелаксантов типа сукцинилхолина. В скелетной мышце миорелаксанты обладают деполяризующим блокирующим свойством, нарушая нервно-мышечную передачу. В наружных мышцах глаза они выборочно инициируют мышечные волокна как «орбитальной», так и «глазной» зоне. Так, небольшие дозы препарата увеличивают регидность мышц. В то же время эти дозы не угнетают реакцию быстрых волокон. Большие дозы сукцинилхолина увеличивают регидность «тонических» волокон и угнетают быстрые волокна. При этом миорелаксанты устанавливают глазное яблоко в первичное положение во время общей анестезии.
Знание этих особенностей действия блокаторов на наружные мышцы глаза имеет практическое значение, поскольку эти препараты нередко используются при общей анестезии, а также в диагностике и лечении косоглазия. Дифференциальное влияние сукцинилхолина на «тонические» волокна, в частности, используют в клинике с целью прогнозирования возможности правильной установки наружных мышц после хирургического вмешательства.
Нередко вводят в глазницу местные болеутоляющие средства группы аминоацилов (лидокаин). Эти препараты обладают миотоксическим действием и могут привести к параличу мышцы. При инъекции указанных препаратов в паренхиму мышцы последние стимулируют высвобождение ионов кальция, приводя к развитию птоза или диплопии во время оперативного вмешательства под местной анестезией.
В клинике используется также ботулиновый токсин, маленькие дозы которого при введении в наружные мышцы глаза изменяют их сократимость. Ботулиновый токсин блокирует высвобождение ацетилхолина в нейромышечное соединение. Это свойство токсина используют в клинике для стимуляции наружных мышц глаза при косоглазии. При этом отмечается расслабление леватора верхнего века и мышц лица, участвующих в дистонических нарушениях.
Инъекция ботулина в скелетную мышцу, включая круговую мышцу глаза, вызывает паралич, иногда приводящий к атрофии мышцы в результате ее химической денервации. Восстановление функции мышц лица может произойти путем повышения функциональной активности мотонейронов (функциональная реиннервация). К сожалению, подобного восстановления наружных мышцах глаза не происходит.
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0