Нуклеотидрегулируемые каналы сенсорных клеток (ЧАСТЬ ІІ)


Содержание:
Описание
Область поры ЦН-канала. В настоящее время полагай IT, что петля между S5 и S6 трансмембранными сегментами (П-область) потенциал-зависимых каналов и гомологичных им ЦН-каналов формирует внутреннюю часть канальной поры. Этот участок канала формирует петлю, вытянутую в направлении центральной оси канала. Помимо того, что П-область является ионселективным фильтром, она функционирует как канальный запирающий механизм, структура которого изменяется, когда канал открывается. Область поры контролирует как проводимость одиночного канала, так и собственно диаметр поры канала. Важной задачей является выяснение того, как ион проникает через канал и каков механизм ионной селективности.
Balasubraminian et al. [19971 считают, что единственное место связывания катионов находится внутри канальной поры. Известно, что движение ионов Ca2+ является одной из составляющих ионного тока ЦН-каналов. В то же время Са2+блокирует ток моновалентных катионов через канал. Предположительно, такое поведение обусловлено высоким сродством Са2+ к кислому аминокислотному остатку - глутамату, расположенному в области канальной поры (Е-363 для ЦН-канала палочки и Е-333 для обонятельных ЦН-каналов сомика). Показано также, что остаток глутаминовой кислоты Е-363 (Е-333) играет ключевую роль при блокаде ЦН-канала внеклеточными протонами, так как два глутамата образуют сайт связывания прогона. Аналогично, данные, полученные Gavazzo et al., свидетельствуют о том. то глутамат F-140 из П-области обонятельного канала цыпленка, эквивалентный Е-363 из канала палочки, контролирует проницаемость и блокаду внеклеточными ионами Са2+ и Mg2+.
Как уже было сказано выше, ЦН-каналы палочек и колбочек по-разному блокируются ионами Са2+ и отличаются по своей относительной Са2+-проводимостп. Исходя из оценки кинетики проводимости ЦН-каналов, Picones, Korenbrot [1995] полагают, что этот эффект обусловлен наличием двух сайтов связывания Са2+. Первый сайт идентичен в палочках и колбочках второй же отличается — либо его энергия взаимодействия в ЦН-канале палочки выше, либо он расположен так что менее подвержен влиянию мембранного потенциала. Такую же модель ионной проницаемости с двумя сайтами связывания предлагают Wells, Tanaka.
Модуляция внутриклеточными протонами а субъединицы обонятельных и палочковых ЦН-каналов цыпленка вызывает увеличение внутриклеточной концентрации протонов и частичную блокаду тока одиночных каналов, как обонятельных, так и палочковых, что является следствием протонирования единственного кислотного сайта с рК1 в пределах 4,5-4,7. Степень активации ЦН-каналов также зависит от внутриклеточного рН. При низкой концентрации циклических нуклеотицов вероятность открытого состояния ЦН-канала значительно увеличивается при понижении рН, причем увеличение это при активации канала цАМФ больше, чем при активации цГМФ Таким образом, внутриклеточная концентрация протонов влияет и на проницаемость ЦН-каналов, и на степень активации, вызывая уменьшение тока одиночного канала и увеличение вероятности открытого состояния.
Таким образом, аминокислотная последовательность П-области ЦН-канала формирует катионселективный фильтр, причем аминокислотный остаток глутамат играет огромную роль в механизме блокирования тока двухвалентными ионами и рН-модуляции проводимости канала.
Сайт связывания Са2+/кальмодулина. Са2+играет огромную роль в работе ЦН-каналов: помимо того, что ток ионов Са2+является существенным компонентом тока ЦН-канала, Са2+ участвует в механизме обратной связи в процессе фототрансдукции: Са2+связывается с модуляторным белком кальмодулином, который, в свою очередь, связываясь с ЦН-каналом, ингибирует канальный ток. Grunwald et al. [1993], изучая меха низм влияния Са2+ /кальмодулина на работу ЦН-канала, идентифицировали два независимых сайта (СаМ1 и СаМ2), с которыми связывается кальмодулин. Они расположены в N и С-концевой областях ?-субъединицы палочкового цГМФ- зависимого канала. Однако только делеция СаМ1 в N-терминальной области лишала ЦН-канал чувствительности к действию Са2+/кальмодулина. Таким образом, можно сделать вывод, что ингибиторное действие Са2+/кальмодулина опосредовано в основном доменом из N-концевой области.
N-С-взаимодействие. Работы последних лет свидетельствуют о том, что взаимодействие N- и С-концевой областей весьма существенно для механизма открывания канала.
Varnum, Zagotta [1996] показали, что внутримолекулярное взаимодействие между N-концевой областью и С-концевым лиганд связывающим доменом ЦН-канала обонятельного нейрона крысы влияет на активацию канала; в свою очередь, Са2+/кальмодулин модулируют активность канала, оказывая влияние именно на это внутримолекулярное взаимодействие. Gordon et al [1997], работая с ?-субъединицей палочкового ЦН-канала, также показали наличие взаимодействия между N- и С-концевыми участками молекулы канала. В этом случае взаимодействие происходило между двумя цистеиновыми остатками: образовывалась дисульфидная связь между С-35 (N-конец) и С-481 (С-связывающая область). Необходимо отметить, что С-481 фоторецепторного канала соответствует С-460 молекулы обонятельного ЦН-канала.
Лиганд-зависимая активация ЦН-каналов. Активация ЦН- канала достигается в результате связывания с несколькими лигандами. Оценить функциональный эффект каждого события связывания молекулы циклического нуклеотида для ЦН-канала, как и для других аллостерических белков, довольно сложно, так как лигандь. постоянно находятся то в связанном, то в несвязанном состоянии (спонтанный, мерцательный характер работы канала). Оценки показывают, что ЦН-каналы находятся в состоянии частичного связывания со своими лигандами, и потому важно было определить поведение таких каналов. Ruiz, Karpen, используя метод высокого разрешения регистрации одиночного канала и фотосвязывающийся аналог цГМФ, который обладает свойством ковалентно связывать цГМФ с его сайтом связывания, показали, что единичный ЦН-канал палочки может связывать до четырех лигандов. Величина канального тока зависит от количества связанных молекул циклических нуклеотидов минимальна - когда связаны два лиганда (~1% от максимального тока), существенно больше - когда связаны три лиганда (-33% от максимального тока), и максимальна - когда связаны четыре лиганда. Более того, в каждом лиганд-связанном состоянии существуют два или три различных состояния проводимости: амплитудные гистограммы показывают, что канал свободно переключается и существует в трех состояниях проводимости (10, 18, 29 pS) независимо от количества связанных лигандов. Особенно четко это свойство ЦН-канала палочки проявляется в состоянии неполной активации. Так, результаты исследования ЦН-кана, :ов с одним несвязанным лигандом показывают, что эти каналы разделяются на два равных класса с константами диссоциации 1,2 и 19 мкМ, что демонстрирует функциональную гетерогенность связывающих сайтов природного ЦН-канала из фоторецепторной палочки, обладающих разным сродством к пГМФ. Дальнейшие исследования позволяют авторам предположить, что подобное поведение канала объясняется, по крайней мере частично, различными свойствами ?- и ?-субъединиц, каждая из которых, как было прямо показано с помощью биохимических экспериментов, связывает цГМФ.
Модуляция ЦН-каналов. В настоящее время известно несколько факторов, регулирующих работу ЦН-каналов: Са2+/кальмодулин, двухвалентные катионы, фосфатаза, эндогенные Са2+-связывающие протеины.
В ЦН-каналах фсьорецепторных клегок и обонятельных нейронов Са2+/кальмодулин модулирует чувствительность канала к лиганду. В ЦН-каналах обонятельных нейронов Са2+ в комбинации с кальмодулином связывается с последовательностью из примерно 30 аминокислот в N-концевой области ?-субъединиц. По всей видимости, сайт связывания Са2+/кальмодулина в палочковых ЦН-каналах расположен в N-концевой области ?-субъединицы, хотя в С-концевой области ?- субьединицы также был идентифицирован потенциальный сайт, участвующий в модуляции Са2+/кальмодулином. Для обоих типов ЦН-каналов взаимодействие с Са2+/кальмодулином зависит от состояния канала: связывание осуществляется с высокой степенью сродства, когда канал закрыт.
?-субьединица ЦН-канала колбочки сетчатки человека также имеет сайт связывания Са2+/кальмодулина в N-концевой области, который расположен приблизительно так же, как в обонятельном ЦН-канале, и обладает способностью связывать Са2+/кальмодулин. Однако гомомерный канал, формируемый этой субъединицей, не обладает чувствительностью к функциональной модуляции Са2+/кальмодулином. Этот эффект, объясняется тем, что на взаимодействие с Са2+/кальмодулином влияет последовательность, расположенная рядом о сайтом связывания Са2+/кальмодулина. В случае обонятельных ЦН-каналов. механизм ингиоированчя которых Са2+/кальмодулином более понятен, обнаружены два конкурирующих типа связывания, зависящих от состояния канала. Первый тип - взаимодействие с ?-кальмодулина с N-концевой областью - вызывает ингибирование канала. Во втором случае в процесс вовлечена С-концевая область в отсутствие N-концевой. Существует мнение, что последовательность из 30 аминокислот, называемая сайтом связывания Са2+/кальмодулина, функционируе г как самовозбуждающий домен, связывающийся с рецепторной областью в С-концевой части предпочтительнее, когда канал открыт. Таким образом, становится ясен механизм ингибирования при связывании ЦН-канала с Са2+/кальмоду пином. В результате захвата N-концевой области Са2+/кальмодулин предотвращав г ее взаимодействие с С-концевой областью, т.е. Са2+/кальмодулин и С-концевая область осуществляют конкурентное взаимодействие с N-концевой областью. Суммируя сказанное выше, можно заключить, что прямое взаимодействие этих N- и С-концевых областей способствует открыванию ЦН-канала и может считаться компонентом этого механизма.
Са2+/кальмодулин модулирует чувствительность ЦН-каналов к лигандам. В палочковых каналах, когда Са2+/кальмодулин взаимодействует с соответствующим связывающим сайтом ?-субъединицы, К1/2 (константа полуактивации канала циклическим нуклеотидом) увеличивается в 1 раза. В обонятельных каналах эффект воздействия Са2+/кальмодулина значительно больше и выражается в 20-кратном уменьшении сродства к цАМФ.
Следует отметить, что сродство ЦН-канала к Са2+ также зависит от мембранного потенциала и внеклеточного рН. Более того, модуляторные субъединицы, коинтегрироьанные с основной функциональной субъединицей, изменяют структуру сайта связывания Са2+, находящегося внутри канальной поры.
Механизм работы канала. Подытоживая сказанное, можно полагать, что ЦН-канал является гетероолигомерным белком, «стоящим из четырех субъединиц. Несмотря на отличие моле-кулярных масс ?- и ?-субъединиц, принципиально их структуры подобны, в том числе схожи ключевые аминокислоты. Все субъединици ЦН-каналов обладают общими структурны ми особенностями - наличием шести трансмембранных сегментов, шпилькообразной области, формирующей канальную пору, домена, связывающего циклические нуклеотиды, Ц-связывающей области и сайта связывания Са2+/кальмодулина.
В результате связывания нескольких лигандов, максимум четырех, происходит аллостерическая трансформация молекулы ЦН-канала, следствием которой является увеличение проводимости. Благодаря изучению молекулярной структуры ЦН-канала стало ясно, что важную роль в этом процессе помимо собственно области связывания ЦН играют N-концевой участок и Ц-связывающая область.
В активированном состоянии через ЦН каналы течет ток, состоящий в основном из ИОНОВ Na+, Ca2+ и Mg2+, направленный внутрь клетки и вызывающий ее деполяризацию. Например, для палочковых ЦН-каналов соотношение ионов Na+, Са2+ и Mg2+ при физиологических условиях составляет примерно 1 : 0,15 : 0,05 соответственно. В области поры ЦН-канала содержится два ион-связывающил с 1Йта: один расположен ближе к внутриклеточной поверхности, другой - остаток глутаминовоч кислоты - ближе к внеклеточной поверхности и находится в позиции 363 в ЦН-канале палочки, в позиции 333 в обонятельном ЦН-канале. Видимо, этот сайт ответствен за блокаду двухвалентными катионами, рН-модуляцию и селективность к одновалентным катионам. Е отсутствие двухвалентных катионов проводимость канала мало зависит от мембранного потенциала.
↑ Регулируемая цАМФ проводимость в мембране слуховых волосковых клеток
Сегодня известно два тина биологических структур, в которых нуклеотьдрегулируемые каналы играют ключевую роль в восприятии сигнала - фоторецепторы и обонятельные нейроны. Сходство ионных механизмов генерации сенсорного сигнала в обонятельных и зрительных рецепторах позволяло предположить, что регулируемые циклическими нуклсотидами каналы могут являться универсальным компонентом мембран рецепторных клеток, участвующим в восприятии сенсорного сигнала. В связи с этим возникло предположение, что каналы такого типа могут присутствовать также и в мембранах рецепторных клеток еще одного класса - механорецепторных клетках. Нами были обнаружены и охарактеризованы нуклеотидрегулируемые каналы в волосковых клетках внутреннего уха.
Проводимость мембран изолированных внутренних и наружных волосковых клеток улитки морской свинки исследовалась методом пэтч-кламп в конфигурации inside-out. После стабилизации тока с цитоплазматической стороны inside-out фрагмента мембраны добавлялся цАМФ. Аппликация 100 мкМ цАМФ в среду инкубации приводила к обратимому увеличению проводимости мембраны волосковых клеток. Результаты одного из таких экспериментов приведены на рис. 4.2. Добавление 1 мМ Mg2+ или Са2+в присутствии цАМФ приводило к обратимому блокированию проводимости.

Аппликация цГМФ в концентрациях до 1 мМ не оказывала влияния на проводимость мембраны. Как и регулируемые циклическими нуклеотидами каналы фоторецепторной и обонятельной клеток, цАМФ-зависимая проводимость волосковых клеток блокировалась 1-цис-одиалтиаземом в концентрации 0,5 мМ. цАМФ-регулируемая проводимость была зарегистрирована на 18 из 37 фрагментов мембраны наружных волосковых клеток и 9 из 22 исследованных фрагментов мембраны внутренних волосковых клеток.
На рис, 4.3, а представлено семейство вольтамперных характеристик deout фрагмента плазматической мембраны волосковой клетки при различных концентрациях цАМФ, а на рис. 4.3, б - соответствующая зависимость проводимости фрагмента мембраны от концентрации цАМФ.
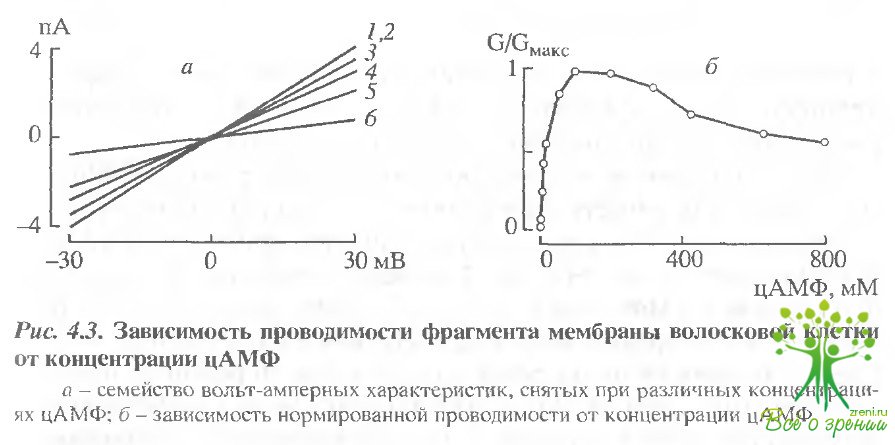
Полумаксимальная активация проводимости достигается при концентрации агониста 20 мкМ. Концентрационная зависимость имеет колоколообразную форму, т.е. большие концентрации агониста частично блокируют проводимость (как и для проводимости фоторецепторной и обонятельной клеток) Коэффициент Хилла, определенный по линейному участку кривой (0-100 мкМ), составляет 1,2.
Селективность цАМФ-регулируемой проводимости определяли по сдвигу потенциала реверсии цАМФ-зависимого тока, вызванному заменой NаСl с цитоплазматической стороны мембраны на эквимолярные количества NаСl, СsСl, RbСl. Сдвиги потенциала реверсии составили -0,3 ± 0,5; 5.4 ± 0.8 и 7,3 ± 1 мВ для Сs+,Li+и Rb+соответственно. Таким образом, проводимость характеризуется слабой селективностью для одновалентных катионов в соответствии с рядом Сs > Na >Li > Rb.
Точно определить сдвиг потенциала реверсии при замене Nа+ на К+не удалось, поскольку мембрана волосковых клеток содержит большое количество К+каналов. Однако можно утверждать, что эта величина не превышает 10 мВ, и таким образом проницаемость цАМФ зависимых каналов для К+ - того же порядка, что и для других одновалентных катионов.
К сожалению, не удалось получить контакт, достаточно высокоомный для регистрации одиночных каналов. Поэтому параметры каналов были определены из анализа флуктуаций цАМФ- активируемого тока. На рис. 4.4 представлен спектр мощности флуктуаций цАМФ-зависимого тока.
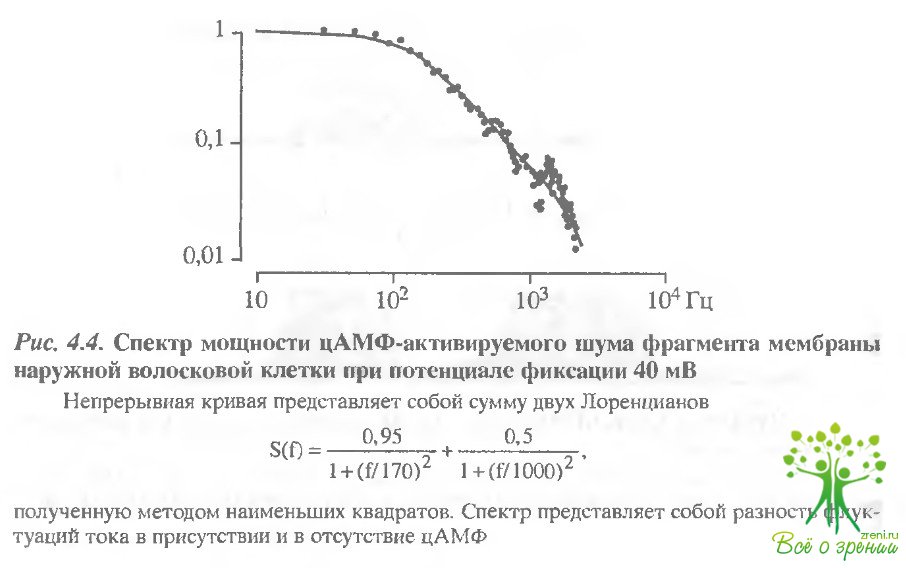
Он представляет собой разность спектров флуктуаций в присутствии и в отсутствие цАМФ. Спектр мощности шума описывается двумя Лоренцианами с характерными частотами 170 и 1000 Гц, что соответствует времени флуктуаций канала около 1 мс. Проводимость одиночного цАМФ-активируемого канала в отсутствие двухвалентных катионов поставляла 6 пСм. Поскольку в присутствии Са2+ или цАМФ-актиьируемая проводимость уменьшалась более чем на порядок, проводимость одиночного канала при физиоло1Нческой концентрации двухвалентных катионов должна быть не более 1 пСм.
Плотность цАМФ-активируемых каналов на теле волосковой клетки, по нашим оценкам, не превосходит 1 мкм. Известно, что плотность регулируемых циклическими нуклеотидами каналов на геле обонятельной клетки и во внутреннем сегменте фоторецептора также очень низка. Каналы сосредоточены в цилии обонятельной клетки и в наружном сегменте фотопецептора, т.е. там, где локализованы процессы сенсорной трансдукции. Если исходить из аналогии с другими типами рецепторов, то можно
было бы ожидать, что плотность цАМФ регулируемых каналов на мембране стереоцилий будет значительно выше, чем на теле волосковой клетки. Однако пока нам не удалось получить гигаомный контакт на мембране стереоцилий, и вопрос о плотности каналов на стероцилиях остается открытым.
Свойства обнаруженных нами цАМФ-активируемых каналов волосковых клеток (селективность, концентрационная зависимость, проводимость одиночного канала, блокаторьи очень сходны со свойствами регулируемых циклическими нуклеотидами каналов (ЦН-каналов) фоторецепторных и обонятельных клеток. цАМФ-активируемая проводимость волосковых клеток частично блокируется большой концентрацией агониста, как и ЦН-проводимость фоторецепторов и обонятельных клеток. В фоторецепторах активируемые цГМФ каналы непосредственно включены в механизм генерации сенсорного сигнала. Поглощение света в фоторецепторе приводит к гидролизу цГМФ и закрытию цГМФ-активируемых каналов
Предполагается, что обнаруженные в мембране обонятельных клеток ЦН-каналы также играют важную роль в механизме трансдукции в этих клетках. Б связи с этим представляется интересным обсудить возможную роль цлМФ-активируемых каналов в механизме трансдукции в волосковых клетках.
Механизм механотрансдукцки в волосковых клетках, по-видимому, принципиально отличается от трансдукции в фоторецепторе и обонятельном нейроне. Трансдукционный ток в волосковых клетках прямо регулируется механочувствительными каналами, которые прямо управляются смещением стереоцилий, причем проводимость этих каналов во много раз выше, чем проводимость ЦН-каналов в фоторецепторах и обонятельных рецепторах. По разным оценкам, эта проводимость составляет до 100 пС, так что трансдукционный ток может обеспечиваться открыванием нескольких подобных каналов. Остается неясным, как при такой высокой проводимостч одиночных каналов в волосковой клетке обеспечивается высокое соотношение сигнал—шум.
По некоторым параметрам свойства ЦН-каналов в волосковых клетках блики к свойствам трансдукционных каналов. Показано, что потенциал реверсии трансдукционного тока в волосковой клетке близок к 0 мВ. Это означает, что трансдукционные каналы должны быть неселективными. Обнаруженные нами цАМФ-активируемые каналы неселективны по отношению к одновалентным катионам, в отличие от всех других известных в настоящее время ионных каналов в волосковой клетке. Однако другая вычисленная характеристика транедукционных каналов - единичная проводимость - не совпадает с проводимостью одиночного цАМФ- регулируемого канала при физиологических концентрациях двухвалентных катионов. По нашим оценкам, проводимость одиночного цАМФ-регулируемого канала в присутствии Mg2+ и Са2+составляет менее 1 пСм, в то время как самая низкая оценка проводимости трансдуционного канала - 12 пСм.
Считается, что транедукционные каналы должны быть сосредоточены в стеоцилиях волосковых клеток. К сожалению, нам не удалось получить гигаомный контакт на мембране стереоцилий, однако, если исходить из аналогии с фоторецепторами и обонятельными клетками, концентрация цАМФ-регулируемых каналов в стереоцилиях могла бы быть велика, поскольку в фоторецепторах и обонятельных клетках концентрация ЦН-регулируемых каналов в аналогах стереоцилий, наружном сегменте фоторецептора и цилии обонятельной клетки значительно выше, чем соответственно во внутреннем сегменте фоторецептора и теле обонятельной клетки. Возможно, ЦН- каналы играют в клетке регуляторную роль. Например, при увеличении концентрации цАМФ в клетке они могут регулировать входящий кальциевый ток. Известно, что внутриклеточная концентрация кальция играет существенную роль в механизме адаптации механорецепторных клеток. Подтверждением этого может служить тот факт, что транедукционный ток в волосковых клетках может регулироваться не только кальцием, но и цАМФ.
Таким образом, в настоящее время нет достаточных аргументов, чтобы утверждать, что цАМФ-регулируемые каналы включены в механизм транедукции в волосковых клетках улитки, однако некоторые их свойства соответствуют свойствам транедукционных каналов. Возможно, механотранедукция в волосковых клетках осуществляется по нескольким параллельным механизмам, и функционирование цАМФ-регулируемых каналов не исключает присутствия других каналов, проводимость которых непосредственно регулируется механическим смещением стереоцилий.
↑ Действие реагентов, модифицирующих SH-группы, и оксида азота на нуклеотид-регулируемые ионные каналы
Известно, что оксид азота (N0) играет важную роль в работе нервной системы. Гистохимические и иммуноцитохимические эксперименты с использованием NADPH-диафоразы показали, что NO-синтаза - фермент, образующий оксид азота из L-аргинина - широко распространен в резных отделах мозга позвоночных. В сетчатке NO-синтаза была обнаружена в разных слоях- в эллипсоидах фоторецепторных клеток, амакриновых клетках, в мюллеровских клетках, в горизонтальных клетках, в биполярных клетках.
Существуют свидетельства того, что NO может модулировать работу ионных каналов фоторецепторных клеток. Экзогенный NO влияет на ток кальциевых каналов из внутреннего сегмента палочки и на неселективный, дилтиаземчувствительный ток (возможно, идентичный цГМФ-зависимому току) в наружном сегменте. Обнаружено также, что оксид азота действует на цГМФ-зависимый синаптический ток в оn-биполярных клетках, может активировать цГМФ-зависимый ток катионов в ганглиозных клетках сетчатки и в фоторецепторных колбочках. В этих случаях эффект NO, по-видимому, обусловлен активацией растворимой гуанилатциклазы, так как увеличение концентрации цГМФ вызывает увеличение тока цГМФ-зависимых каналов.
Однако возможен и другой механизм регуляции оксидом азота. Выяснилось, что действие NO многообразнее и, в частности, он может и прямо активировать ионные каналы нервных клеток. Это было продемонстрировано для НМДА рецептора и для калиевых каналов.
В рецепторных тканях также обнаружено и прямое действие NO на каналы. Таким образом, NO может влиять на величину активации цГМФ-зависимых каналов различными способами. В этом разделе мы остановимся на описании экспериментов по прямому действию оксида азота на нуклеотидрегулируемые каналы. Основанием для такого подхода послужили описанные выше данные по прямой активации цГМФ-регулируемых каналов из зрительных клеток путем модификации их сульфгидрильных групп. Известно, что кроме действия N0 как лиганда в гемсодержащих белках, он также хорошо связывается с сульфгидрильными группами, образуя нитрозотиолы.
В описанной ниже серии экспериментов исследовалось прямое действие реагентов, модифицирующих сульфгидрильные группы, и продуцентов оксида азота на одиночные цАМФ-регулируемые каналы волосковых клеток и цГМФ-регулируемую проводимость наружного сегмента фоторецепторной клетки.
Статья из книги: Молекулярные механизмы зрительной рецепции | Каламкаров Г.Р., Островский М.А..

Комментариев 0