Электрофизиология фоторецепторов


Описание
Фотоизомеризация и мембранный потенциал.Проблема «посредника». Реакцией фоторецептора позвоночных на свет является гиперполяризация, возникающая в результате уменьшения проницаемости плазматической мембраны наружного сегмента для ионов Nа+.
До настоящего времени не выяснен механизм, посредством которого фотоиндуцированные превращения зрительного пигмента приводят к изменению натриевой проводимости (механизм «трансдукции»). Предполагается, что трансдукция осуществляется с помощью внутриклеточного химического посредника.
Предположение возникло из чисто формальных требований динамической модели фотопотенциала, было подкреплено в дальнейшем рядом экспериментальных данных. Электронно-микроскопическими методами было обнаружено, что диски палочек, содержащие родопсин, структурно и электрически разобщены с плазматической мембраной (между ними 100-200 А в местах наиболее близкого смыкания).
Таким образом, требуется переносчик сигнала с диска на поверхностную мембрану. Статистический анализ темнового «электрического шума» в наружных сегментах рецепторов показал, что фотолиз одной молекулы пигмента приводит к закрытию 300-400 натриевых каналов в палочке и около 25 каналов в колбочке. Откуда следует, что одна молекула пигмента не может служить регулятором отдельного канала и необходим посредник, осуществляющий «размножение» сигнала.
В начале 70-х годов на роль внутриклеточного посредника были предложены ионы кальция и циклические нуклеотиды (циклический аденозинмонофосфат — цАМФ, и циклический гуанозинмонофосфат — цГМФ). По «кальциевой гипотезе» свет вызывает выход Са2+ из дисков в наружных сегментах палочек (вход ионов Са2+ в наружные сегменты колбочек), который диффундирует к плазматической мембране и блокирует натриевые каналы.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Светоиндуцированный выход ионов Са2+ можно зарегистрировать кальцийчувствительными электродами. Повышение содержания Са2+ на свету происходит довольно быстро и в количествах, достаточных для вызова ответа фоторецептора. Однако искусственно вызванные изменения концентрации Са2+ не воспроизводят полностью влияния реального света на кинетику и чувствительность фоторецепторного потенциала. Поэтому кальциевую гипотезу безоговорочно принять невозможно.
Согласно другой, альтернативной, гипотезе в темноте цГМФ- (цАМФ) держит открытыми натриевые каналы и натрий входит в клетку. На свету распад пигмента активирует фосфодиэстеразу, которая разрушает цГМФ (цАМФ), что сопровождается закрытием натриевых каналов и, как следствие, гиперполяризацией рецептора.
Циклические нуклеотиды являются универсальными внутриклеточными посредниками в нервной системе — участвуют в синаптической передаче, рецепции гормонов, регуляции внутриклеточного кальция, фосфорилировании ферментов и сократительных белков.
Обесцвечивание одной молекулы пигмента приводит к гидролизу 1000-2000 молекул цГМФ за 100-300мс, что сравнимо со временем развития рецепторного потенциала при пороговых интенсивностях света.
Наружные сегменты фоторецепторов содержат фермент гуанилатциклазу, синтезирующую цГМФ из АТФ, и в небольших количествах — фермент аденилатциклазу, опосредующую синтез из АТФ цАМФ. Однако проводимость натриевых каналов наружного сегмента палочки не зависит от концентрации цГМФ. С другой стороны, свет вызывает большие изменения в метаболизме молекул цГМФ, а экспериментально индуцированные изменения в метаболизме цГМФ оказывают большое влияние на кинетику и чувствительность реакции фоторецептора.
Эти данные свидетельствуют о том, что синтез цГМФ может играть важную регуляторную роль в механизме генерации ответов фоторецепторов, даже если цГМФ сам по себе и не является посредником, контролирующим натриевые каналы. Наличие каскада ферментативных реакций, регулирующих уровень цГМФ, показано только в палочках. В колбочках светорегулируемым является, по-видимому, цАМФ.
Ни «кальциевая», ни «циклонуклеотидная» гипотезы по отдельности не могут объяснить механизма трансдукции. Все больше данных в пользу их совместного действия. Согласно одной из последних гипотез посредником в процессе трансдукции является кальций, высвобождение которого из дисков и обратный захват регулируются цМГФ как частью системы сервоконт- роля.
Ионные механизмы реакций фоторецепторов.
Структурное разделение рецепторов на наружный и внутренний сегменты соответствует функциональным различиям ионных каналов и механизмов их регуляции в наружном и внутреннем сегментах. Мембрана наружного сегмента рецептора позвоночных содержит только натриевые ионные каналы, управляемые светом через систему трансдукции.
Мембрана внутреннего сегмента обладает селективной проницаемостью к ионам Са2+, С1_ и К+. Калиевые каналы делятся на два типа: вольт-зависймые и Са2+-зависимые. Ионные каналы внутреннего сегмента отличаются от ионных каналов наружного сегмента тем, что они не чувствительны к свету и являются вольт-зависимыми. В мембране внутреннего сегмента имеется также натрийкалиевый насос.
В темноте мембрана фоторецептора максимально проницаема для ионов Nа+ и К+ и значительно меньше для ионов С1- и Са2+, что определяет темновой уровень мембранного потенциала в —35-;—45 мВ, близкий к потенциалу равновесия возбуждающего постсинаптического потенциала (ВПСП).
Темновой ток поддерживается непрерывной работой АТФ-зависимого натрий-калиевого насоса во внутреннем сегменте, закачивающего калий в клетку и выводящего натрий наружу. Ионы натрия входят в наружный сегмент, а ионы калия выходят из внутреннего в соответствии со своими электрохимическими градиентами (в палочках потенциал равновесия для Na+ равен +5-+10 мВ, для К+ — —60-—65 мВ). Суммарный темновой потенциал (—35-—45 мВ) равномерно распределен по всей оси рецептора и имеет одно и то же значение в наружном и внутреннем сегменте. Мембранный потенциал рецептора в темноте Vm равен:
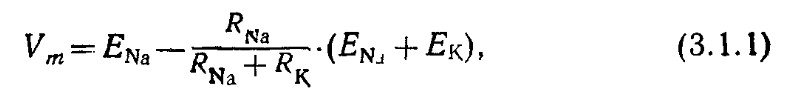
Где ENa и Ек — потенциалы равновесия (потенциалы Нернста) для ионов натрия и калия соответственно. Сопротивление мембраны определяется как эквивалентное сопротивление натриевых и калиевых каналов:

Уравнение (3.1.1) и (3.1.2) следует из эквивалентной электрической схемы фоторецепторной мембраны, составленной с учетом проводимости для двух основных потенциалобразующих ионов: калия и натрия. На свету внутренний трансдуктор («посредник»: Са2+, цГМФ или цАМФ) блокирует натриевые каналы наружного сегмента, что приводит к гиперполяризации рецептора, которая электротонически достигает внутреннего сегмента.
Закрытие натриевых каналов сопровождается увеличением сопротивления мембраны наружного сегмента с 103-104 ом-см2 в темноте до 106 ом-см2 на свету. При уменьшении натриевой проводимости потенциал фоторецептора определяется в основном проводимостью по калию: действительно, при больших яркостях света, когда все натриевые каналы закрыты, мембранный потенциал рецептора близок к потенциалу Нернста для ионов калия.
Изменение мембранного потенциала на свету — это результат не только блокады натриевых каналов наружного сегмента под воздействием света, но и последующего подключения вольт-зависимых кальциевых и калиевых ионных каналов мембраны внутреннего сегмента. Потенциал-зависимые кальциевые каналы имеют важное значение, поскольку вход кальция в пресинаптическую часть фоторецептора определяет выход медиатора. Фоторецептор реализует градуальное управление выходом медиатора. Гиперполяризация, вызванная светом, ведет к закрытию потенциал-зависимых кальциевых каналов и уменьшению концентрации свободного кальция в клетке, что в свою очередь уменьшает выход квантов медиатора из пресинаптической части рецептора.
Таким образом, гиперполяризационная реакция рецептора на свет является прекращением длящейся в темноте деполяризации мембраны. Необычность ситуации в том, что стимулом, возбуждающим фоторецептор (деполяризующим его), служит здесь не свет, а темнота. Это отличие его от рецепторов других модальностей у позвоночных, генерирующих рецепторньгй потенциал в виде деполяризации, а также от фоторецепторов беспозвоночных, реакция которых на свет носит деполяризационный характер.
Различия в знаке реакции фоторецепторов позвоночных и беспозвоночных связывают с различиями в эволюционном развитии фоторецепторов разных типов: 1) рабдомерный тип фоторецептора у беспозвоночных (моллюски, черви, членистоногие) связан всегда с деполяризацией на свету и 2) ресничный (жгутиковый) тип фоторецепторов у позвоночных — с гиперполяризацией.

Комментариев 0