Исследование роли цитоскелета в функционировании фоторецепторных клеток сетчатки


Описание
Наружные сегменты палочек сетчатки содержат разветвленную сеть цитоскелетных структур, в то время как в наружных сегментах колбочек цитоскелет гораздо менее развит.
Прежде всего это связано с чисто морфологическими особенностями. Палочки, работающие в условиях низких освещенностей, должны содержать намного больше зрительного пигмента и поэтому они намного длиннее колбочек. Основная функция цитоскелета - поддерживать правильную форму клетки и поэтому в палочках его морфологическая роль существенно выше. Однако как палочки, так и колбочки состоят из двух обособленных частей - наружного и внутреннего сегментов. Все метаболические процессы, необходимые для поддержание жизнедеятельности клетки, сосредоточены во внутреннем сегменте и поэтому их транспорт в наружный сегмент может быть значительно затруднен из-за больших размеров клеток. Существенную роль в этом процессе мог бы сыграть аксональный транспорт. В этом разделе представлены эксперименты, в которых с помощью ингибиторного анализа мы попытались подойти к исследованию вопроса об участии цитоскелета в регуляции процесса фототрансдукции.
Влияние колхицина на регистрируемый внутриклеточно фотоответ палочек сетчатки лягушки.В этом исследовании фотоответ палочек регистрировался внутриклеточно, что позволяет контролировать как величину фотоответа, так и величину разности потенциалов на плазматической мембране фоторецептора. Эксперименты проводились на препаратах изолированной сетчатки. Колхицин, ингибитор роста микротрубочек, хорошо проникает в клетки через плазматическую мембрану. Поэтому его действие проявляется уже при экстраклеточной аппликации. Результаты такого эксперимента представлены на рис. 7.1. Как оказалось, колхицин оказывает существенное влияние на состояние поляризации мембраны.
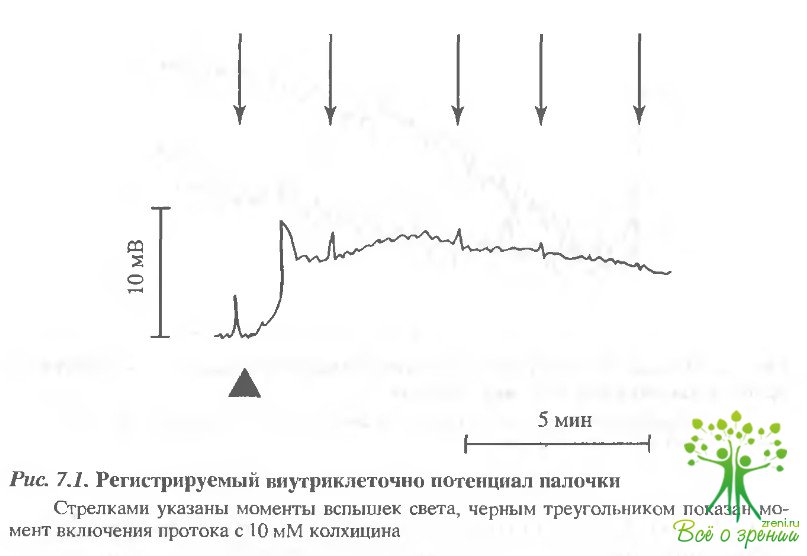
После удачного проникновения в клетку и регистрации контрольного фотоответа на тестовую вспышку света включали проток с колхицином. Это приводило к кратковременной гиперполяризации и последующей деполяризации мембраны клетки. При этом амплитуда ответа на свет значительно снижалась. На рис. 7.2 представлены разрешенные во времени фотоответы палочки до воздействия колхицином, контрольные и через 5 мин после введения в проток колхицина, т.е в момент относительной стабилизации фотоответа Амплитуда существенно снизилась, кроме того, изменилась кинетика фотоответа. Из рис. 7,2, б, где фотоответы нормированы, видно, что спад фотоответа значительно ускорился, в то время как фронт нарастания практически не изменился.
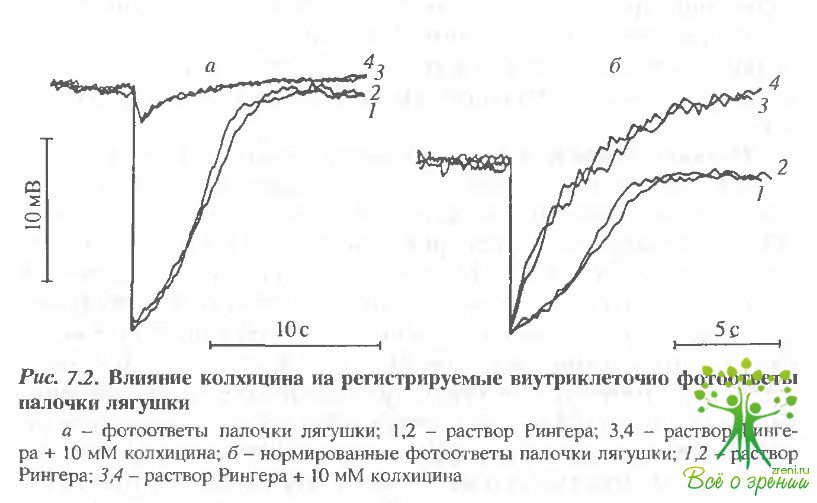
Эффект, как и предполагается, практически необратим. При отмывании сетчатки от колхицина раствором Рингера в течение 30 мин амплитуда фотоответа частично восстанавливалась, но кинетика ответа не возвращалась к исходному виду.
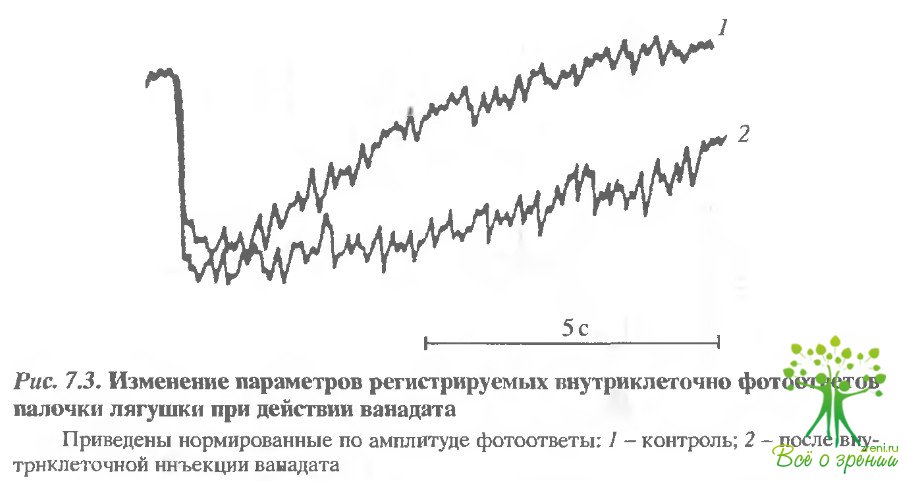
Влияние ванадата на фотоответ палочки. Известно, что ванадат является ингибитором АТФаз, связывающих фосфат Было прямо показано, что он блокирует Mg2+-АТФ-зависимый рост филаментов в наружных сегментах палочек быка. В отличие от колхицина, ванадат не проникает в клетки через плазматическую мембрану. Поэтому его вводили в клетку непосредственно через микроэлектрод с помощью ионофореза, импульсами тока I нА. На рис. 7.3 представлены совмещенные фотоответы, зарегистрированные до и после введения ванадата в клетку. Спад фотоответа под действием ванадата замедляется, в то время как фронт нарастания, как и в опытах с колхицином, практически не меняется. На амплитуду ответа ванадат существенного влияния на оказывал.
Влияние колхицина на суммарный ПРП сетчатки. Поскольку в колбочках не обнаружена столь развитая сеть цитоскелета, как в палочках, интересно было сравнить влияние ингибиторов цитоскелета на фотоответы палочек и колбочек. К сожалению, регистрация внутриклеточных фотоответов колбочек лягушки невозможна технически. Влияние колхицина на колбочки оценивалось по его действию на колбочковый компонент суммарного аспартатизолированного трансретинального ответа фоторецепторов сетчатки лягушки. Известно, что при низких интенсивностях свеча вклад в этот ответ в основном дают палочки, в то время как при высоких интенсивностях света фотоответ обусловлен в основном колбочками сетчатки. Выяснилось, что влияние колхицина на палочковую оставляющую суммарного фотоответа значительно больше, чем на колбочковую.
Колхицин сильно подавляет ответ палочек и практически не влияет на фотоответ колбочек. Факт различного действия колхицина на палочки и колбочки является дополнительным контролем за тем, что колхицин в этих экспериментах действует специфично на цитоскелет, организация которого в палочках и колбочках различна.
Таким образом, вещества, влияющие на состояние цитоскелета, оказывают заметное влияние на амплитуду и кинетику фотоответа фоторецептсров. При этом и колхицин, который ингибирует рост микротрубочек, и ванадат, ингибирующий рост микрофиламентов, оказывают воздействие только на кинетику спада фотоответа, кинетика же нарастания в обоих случаях практически не меняется. Тот факт, что воздействие на различные элементы цитоскелета приводит к изменению только заднего фронта фотопотенциала, свидетельствует, по-видимому, о том, что цитоскелет задействован в регуляции процессов "выключения" сигнала и в механизме релаксации.
Трудно объяснить факт противоположного действия колхицина и ванадата на кинетику спада фотоответа. Следует заметить, что, хотя и показано прямое действие этих веществ на цитоскелетные элементы, не исключена возможность, что они действуют и на какие-то другие белки, участвующие в механизме "выключения" сигнала.
Приведенных данных явно недостаточно для строгого обоснования каких-либо гипотез о механизмах участия цитоскелета в фототранедусции. Регуляторная функция цитоскелета могла бы состоять в избирательном транспорте белков или метаболитов, или в ограничении их свободной диффузии. Такой транспорт мог бы осуществляться как в радиальном, так и в продольном направлениях (вдоль оси наружного сегмента). Последнее может быть особенно существенно, поскольку, как обсуждалось в гл. 1, структура зрительной клетки такова, что синтез всех необходимых для фетотранедукции метаболитов осуществляется во внутреннем сегменте пространственно отделенном от наружного сегмента, где протекают процессы преобразования света.
Статья из книги: Молекулярные механизмы зрительной рецепции | Каламкаров Г.Р., Островский М.А..

Комментариев 0