Биполярные клетки

Описание

Основные физиологические типы биполярных клеток — деполяризационные (с on-центром рецептивного поля) и гиперполяризационные (с off-центром) [Kaneko, 1970, Matsumoto, Naka, 1972; Бызов, 1984]. Установлены также реакции биполярных клеток на цветовые раздражители, не одинаковые у различных позвоночных [Yazulla, 1976; Kato, Negishi, 1979, Witkovsky, Stone, 1983]. Морфология биполярных нейронов известна из классических трудов А. С. Догеля [Dogiel, 1883; Догель, 1888] и Ramon у Cajal [1972]. Polyak [1941] обнаружил в сетчатке приматов новый тип нейрона — карликовую биполярную клетку.
Существенные детали строения, дополняющие классические описания мы находим в интересных современных исследованиях сетчатки рыб [Scholes, 1975; Saito, Kujiraoka, Yonaka, 1983], суслика [West, 1976], кошки [Boycott, Kolb, 1973b], приматов [Boycott, Dowling, 1969; Kolb, 1970; Mariani, 1981] и многих других.
Биполярные клетки в сетчатке рыб весьма разнообразны. Еще Ramon у Cajal [1972] на рисунке сетчатки окуня прекрасно показал семь их разновидностей, отличающихся характером ветвлений дендритов и аксонов. В тексте, однако, Ramon у Cajal выделил лишь два основных типа: палочковые и колбочковые биполяры, принципиально отличающиеся по своим соединениям с рецепторами и ганглиозными клетками, с небольшими вариациями в структуре внутри каждого основного типа. Полностью подтвердились данные Ramon у Cajal о колбочковых биполярных клетках. Они действительно меньшей величины, восходящие дендриты тонки и длинны и образуют синапсы с колбочками [Stell, 1967; Scholes, 1975]. В отношении же палочковых биполярных клеток выяснилось, что в сетчатке рыб они связаны синаптически не только с большим числом палочек, но и с некоторым количеством колбочек.
Детально описаны биполярные клетки в сетчатке рыбы Scardinus erythrophtalmus [Scholes, 1975]. Автор считает, что имеется не менее десяти разновидностей нейронов, отличающихся как по общеморфологическим признакам, так и по синаптическим связям с колбочками и палочками. В основной схеме Scholes представил семь видов биполярных клеток. Палочковые биполяры с узким дендритным полем образуют синапсы с палочками и основными членами двойных колбочек (красночувствительными). Избирательные колбочковые биполярные клетки образуют синапсы с добавочным членом двойной колбочки (зеленочувствительным). Другие избирательные колбочковые биполярные клетки вступают в синаптическую связь с одиночными колбочками (синечувствительными). Смешанные биполярные клетки связаны с двумя разновидностями колбочек, красно- и зеленочувствительными, либо с зелено- и синечувствительными; они характеризуются очень малым дендритным полем и окончанием аксона па разных уровнях. Таких нейронов, по Scholes,— три разновидности.
Детальный анализ связей биполярных клеток с фоторецепторами в сетчатке рыб проведен при помощи внутриклеточной инъекции пероксидазы хрена с последующей электронной микроскопией. Так, у американского сомика (Ictalurus punctatus) дендриты биполяров обоих функциональных типов образовывали инвагинирующие синапсы в фоторецепторах, но окончания их были различны. У off-биполяров терминали отростков приближались максимально к синаптическому гребню непосредственно под синаптической лентой. Терминальные отростки дендритов on-биполяров никогда не располагались под синаптическим гребнем и лентой, между ними всегда находились терминали off-биполяров [Sakai, Naka, 1983]. В основном сходная организация синаптической связи биполярных клеток с фоторецепторами выявлена теми же методиками в сетчатке карпа [Saito, Kujiraoka, Yonaka, 1983].
У некоторых разновидностей биполярных клеток в сетчатке ряда рыб имеются колбы Ландольта [Witkovsky, Stell, 1973; Locket, 1970], что не было отмечено Ramon у Cajal.
Во внутреннем плексиформном слое сетчатки рыб утолщения по ходу аксона биполярных клеток распределены четко регулярно, образуя правильные решетки [Vrabec, 1966; Подугольникова, Максимов, 1975]. По Т. А. Подугольниковой и В. В. Максимову [1975], в сетчатке сельдевых рыб периодические решетки расположены на разных уровнях внутреннего плексиформного слоя, причем они взаимосогласованы, различаясь лишь ориентацией, периодами и величиной входящих в них утолщений. В сетчатке ставриды черноморской сочетание обычных гистологических методик с импрегнацией по Гольджи позволило уточнить взаимоотношения решеток, образованных утолщениями аксонов на пяти уровнях внутреннего плексиформного слоя с типами биполярных клеток. Из восьми типов биполярных клеток у трех — с неразветвленным основным дендритом и с малым размахом дендритных ветвлений — утолщения аксонов формируют решетки различных уровней [Подуголышкова, 1981].
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Функциональные типы биполярных клеток отличаются как особенностями и топографией синаптических связей с фоторецепторами, так и уровнем окончания аксонов во внутреннем плексиформном слое. Например, в сетчатке карпа при внутриклеточной окраске проционовым желтым выявлены биполярные клетки с диаметром дендритных полей 35—50 мкм, реагирующие деполяризацией на освещение центра рецептивного поля. Аксон этих клеток оканчивается в подслое b внутреннего плексиформного слоя. У клеток, аксон которых оканчивается в подслое а внутреннего плексиформного слоя, реакция на пятно света в центре рецептивного поля — гиперполяризация. У малых колбочковых биполярных клеток тело и дендриты небольших размеров; аксон оканчивается либо в подслое b при деполяризационной реакции на освещение центра рецептивного поля, либо в подслое а при гиперполяризационной реакции [Famiglietti, Kaneko, Tachibana, 1977].
Внутриклеточные инъекции пероксидазы хрена или проционового желтого применялись также после микроэлектродных отведений от биполярных клеток сетчатки плотвы (Tribolodon hakonensis) [Hashimoto, Inokuchi, 1981]. Выделено два основных типа клеток — цветовые и нецветовые. У нецветовых клеток с оп-центром терминали аксона достигают подслоя b внутреннего плексиформного слоя, с off-центром — подслоя а. Цветовые биполярные клетки имеют рецептивные поля с антагонистической периферией, морфологически они сходны с нецветовыми клетками с on-центром.
В сетчатке лягушки впервые биполярные клетки целиком со всеми отростками наблюдал А. С. Догель [Dogiel, 1888b]. При окраске метиленовой синью он увидел один тип нейрона, тело которого расположено в центре внутреннего ядерного слоя, дендриты и аксоны образуют сети в наружном и внутреннем плексиформном слоях. От основного дендрита всегда отходит колба Ландольта. В наиболее полном исследовании Ramon у Cajal [1972] они подразделяются на два основных типа: большие наружные без колб Ландольта, малые внутренние с колбами Ландольта. На иллюстрациях Ramon у Cajal видно необычайное разнообразие этих нейронов как по структуре дендритов, так и по аксонам, хотя в тексте наружные биполяры подразделены по характеру дендритов лишь на две разновидности (палочковые и колбочковые), а внутренние подразделены лишь на основании многообразия и расслоенности аксонов.
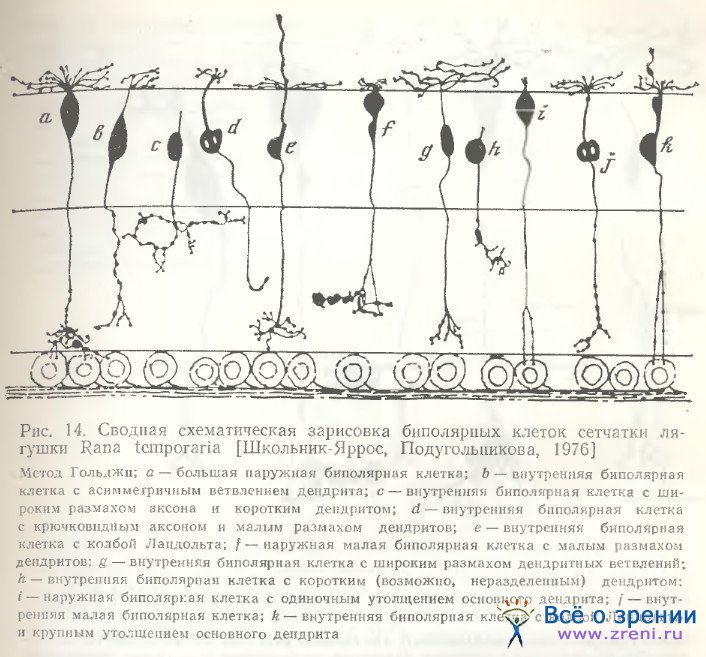
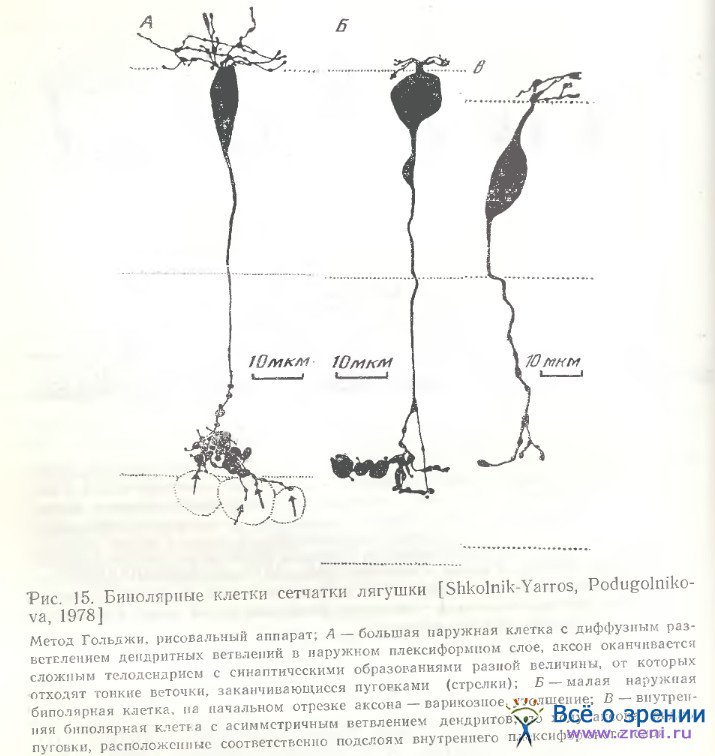
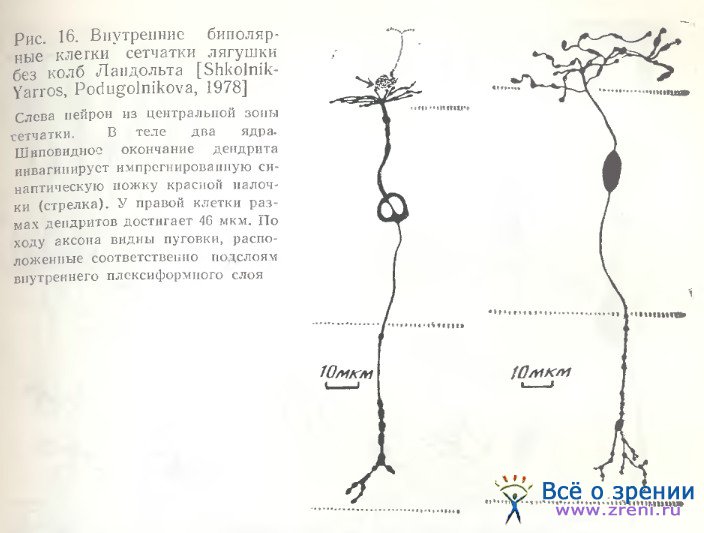
Как мы могли убедиться [Школьник-Яррос, Подугольникова, 1976; Shkolnik-Yarros, Podugolnikova, 1978], наружные биполярные клетки не только крупные (рис. 14, а; 15, А), среди них наблюдаются и малые (рис. 15, Б), и средних размеров (рис. 14, г), с небольшим размахом дендритов или одиночным его ветвлением (рис. 14, f, t). Внутренние биполярные клетки не обязательно малые, встречаются и крупные (рис. 14, b; 15, В). Можно выделить и несколько разновидностей внутренних биполярных нейронов, лишенных колб Ландольта (рис. 14, b, f, g; 15, В; 16; 17, A), включающих клетки с широким размахом дендритов (рис. 16, справа), с узким размахом дендритов (рис. 14, d), с асимметричным их ветвлением (рис. 14, b; 15, В) и неразветвленные. Представленные на рис. 14 (с, h) нейроны неполностью импрегнированы. На рисунках Ramon у Cajal есть сходные разновидности, a Matsumoto, Naka [1972] получили не только импрегнацию такой же клетки, но и окрасили ее при внутриклеточной метке. Эти данные позволяют предположительно отнести клетки па рис. 14 (с, h) к имеющим неразветвленный дендрит.
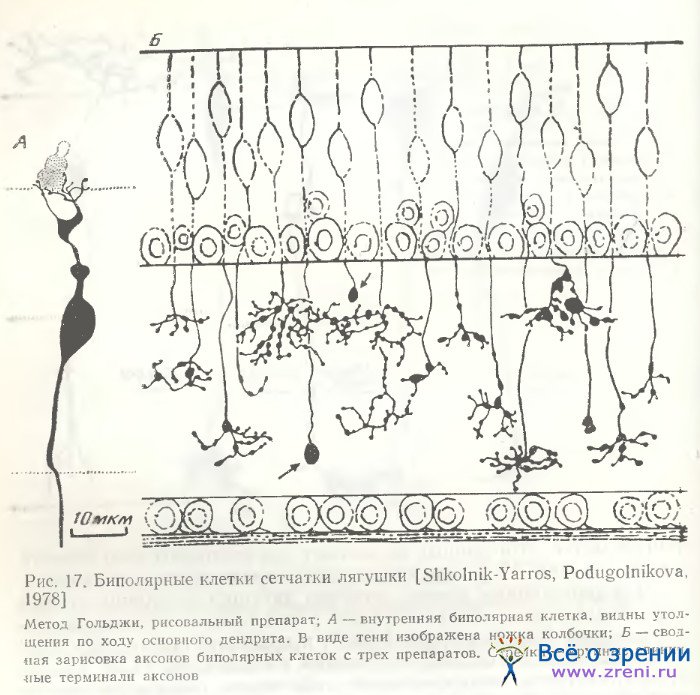
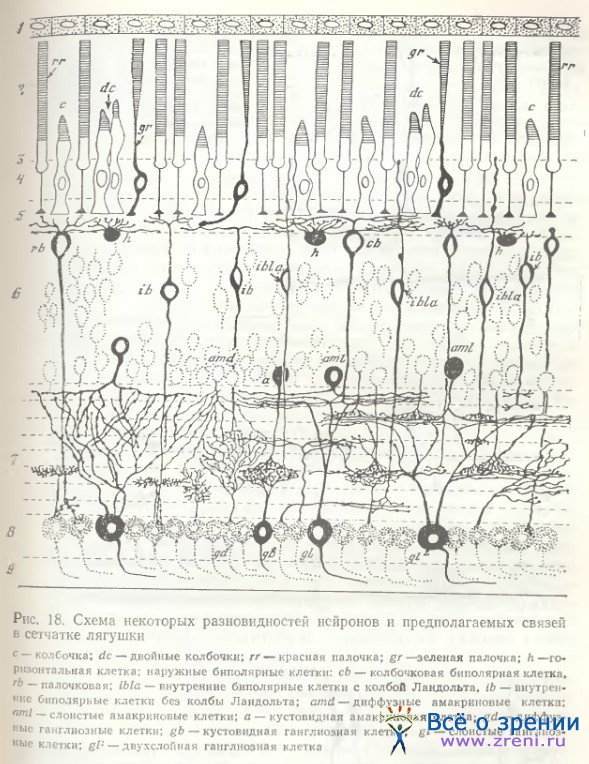
Для биполярных клеток сетчатки лягушки в отличие от рыб характерна также многоэтажность разветвлений их аксонов, обнаруженная Ramon у Cajai [1972], и предполагаемые аксосоматические связи с телами ганглиозных клеток (рис. 14, а; 15, А). Подобная связь осуществляется либо очень тонким разветвлением раздвоенного аксона, терминали которого охватывают небольшое тело ганглиозной клетки (рис. 14, i, k), либо входящими от крупного, сложноразветвленного окончания аксона нежными пуговчатыми терминалями (рис. 14, а; 15, А). Установить, однако, имеется ли в данном случае химический синапс или щелевой контакт, либо связь лишь кажущаяся, можно только при электронно-микроскопическом исследовании. Среди других окончаний аксонов следует отметить также единичное крупное образование, многократно описанное в сетчатке рыб [Vrabec, 1966; Подуголышкова, Максимов, 1975; Scholes, 1975], по впервые наблюдаемое нами в сетчатке лягушки (рис. 17, Б, стрелки). Часто по ходу аксонов можно наблюдать мелкие синаптические пуговки, иногда расположенные на одинаковых расстояниях друг от друга (рис. 14, b; 15, В; 16; 18) [Shkolnik-Yarros, Podugolnikova, 1978].
Таким образом, биполярные клетки сетчатки лягушки характеризуются значительным многообразием. По характеру и способу окончания дендритов и аксонов и наличию либо отсутствию колб Ландольта можно выделить не менее 12 их разновидностей (см. рис. 14—18). При дальнейших электронно-микроскопических исследованиях эти цифры могут быть уточнены и сведены к меньшему числу основных типов в зависимости от синаптических связей.
В сетчатке черепахи (Pseudemys scripta elegans) девять типов биполярных клеток; колба Ландольта имеется у всех видов нейронов. По характеру окончания аксонов однослойными являются лишь две разновидности биполяров, остальные — двух- и трехслойные [Kolb, 1982]. Как и у других позвоночных, аксоны деполяризационных (с on-центром) биполяров оканчиваются во внутренней половине внутреннего плексиформного слоя (подслой b). Морфология гиперполяризационных биполярных клеток сетчатки черепахи не полностью соответствовала предложенной Famiglietti, Kolb [1976] схеме разделения внутреннего плексиформного слоя па два функциональных подслоя. Так, аксоны гиперполяризационных биполяров оканчивались не только в наружной половине внутреннего плексиформного слоя, по и частично во внутренней его половине (подслой b) [Weiler, Marchiafavа, 1981].
В сетчатке птиц (куриных) биполярные клетки морфологически весьма четко подразделяются на два типа: крупные с телом, расположенным в наружной части внутреннего плексиформного слоя, и малые, тело которых находится в средних его отделах. У малых клеток имеется колба Ландольта, тонкие аксоны отдают коллатерали на двух или трех уровнях внутреннего плексиформного слоя [Ramon у Cajal, 1972].
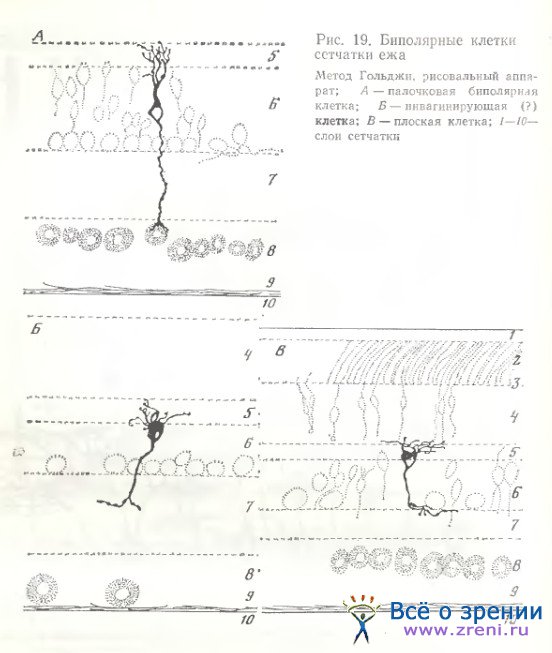
Биполярные клетки сетчатки млекопитающих. В сетчатке ежа биполярные клетки лишены колб Ландольта. Часть нейронов сходна с палочковым типом биполяра, аксонное окончание этих клеток прилегает к телам ганглиозных клеток (рис. 19, А). у других нейронов ветвления дендритов и аксонов подобны кол-бочковым: аксоны более плоски и расположены выше во внутреннем плексиформном слое (рис. 19, В). Некоторые биполярные клетки сетчатки ежа (рис. 19, Б) по морфологии отростков сходны с инвагинирующими в сетчатке кошки (нами приведены лишь предварительные данные по сетчатке ежа). В сетчатке суслика [West, 1976] импрегнацией по Гольджи выявлено не менее семи различных подтипов биполярных клеток. При Гольджи - электронной микроскопии West обнаружил следующую закономерность. Биполярные клетки с наружно расположенным (склерально) телом и внутренне расположенными (витреально) аксонными терминалями преимущественно образуют инвагинирующие синапсы в фоторецепторах. Все биполярные клетки (три подтипа) с внутренне расположенными клеточными телами и наружно расположенными аксонами образуют лишь плоские синапсы (не инвагинирующие) с фоторецепторами. West также отметил, что плоские биполярные клетки характеризуются более сложными и разветвленными аксонными терминалями, в частности, они могут быть двухслойными.
Из представителей отряда хищных наилучшим образом изучены биполярные клетки сетчатки кошки [Boycott, Kolb, 1973b, Famiglietti, 1980; Kolb, Nelson, Mariani, 1981; Nelson, Kolb, 1983]. По данным Гольджи - электронной микроскопии различаются три основных типа этих нейронов: палочковые, колбочковые плоские и колбочковые инвагинирующие биполярные клетки. У палочковых биполярных клеток тонкие терминали дендритов инвагинируют сферулу палочки, занимая центральное положение в триаде, напротив синаптической ленты, между терминалями отростков горизонтальной клетки. С одной палочковой биполярной клеткой контактируют примерно 15 палочек [Kolb, Nelson, 1983]. В наружном плексиформном слое типичные синапсы с палочковыми биполярными клетками образуют восходящие отростки интерплексиформных клеток. Крупные аксонные терминали палочковых биполярных клеток оканчиваются во внутреннем отделе внутреннего плексиформного слоя, вблизи от тел ганглиозных клеток, не вступая с ними в синаптическую связь.
Множество реципрокных синапсов наблюдается между этими терминалями и несколькими разновидностями амакриновых клеток. Плоские колбочковые биполярные клетки образуют поверхностные синапсы с основанием ножки колбочки. От 8 до 14 колбочек контактирует с одной плоской клеткой. Тела плоских колбочковых биполяров расположены в среднем или нижнем отделе внутреннего ядерного слоя. Boycott, Kolb [1973b] наблюдали две разновидности плоских биполяров, различающихся томографией тела и характером аксонного окончания. Тела инваnik-Yarros [1971] соответствует инвагинирующему нейрону, а не палочковому, как ранее обозначено. Однако без электронном микроскопии окончательно установить природу связей клетки нельзя. Недавно Famiglietti [1980] в работе, специально посвященной колбочковым биполярным клеткам в сетчатке кошки, выделил значительно большее количество их разновидностей. Так, автор считает, что по характеру ветвления дендритов, аксонов и их стратификации во внутреннем плексиформном слое можно выделить 10 отчетливо различающихся типов клеток.
Новыми типами биполяров являются широкоразмашистые, контактирующие с колбочками в большой зоне сетчатки, и биполярные клетки с узким дендритным полем, аксонное ветвление которых оканчивается как выше, так и ниже границы между двумя основными подслоями внутреннего плексиформного слоя. По Famiglietti [1980], по-видимому, существует несколько параллельных путей от колбочек через биполяры к одному и тому же подслою внутреннего плексиформного слоя. По данным Kolb, Nelson, Mariani [1981], в сетчатке кошки имеется 8 различных типов колбочковых биполяров (метод Гольджи).
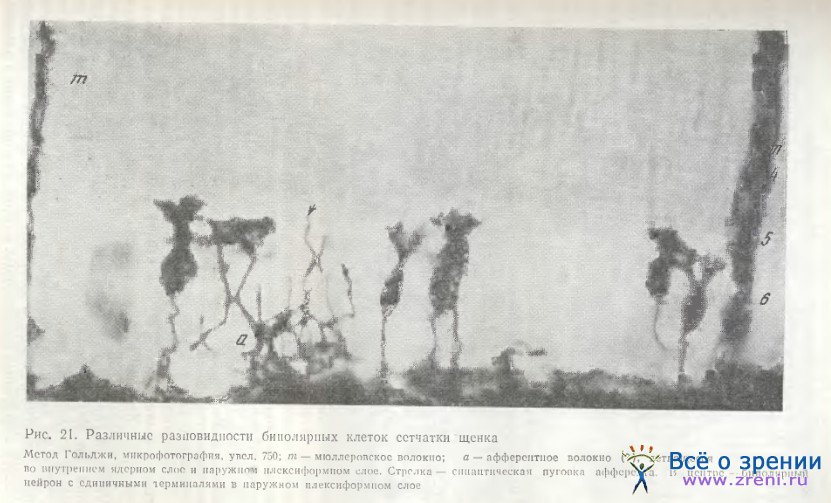
Чрезвычайное разнообразие дендритных ветвлений у биполярных клеток сетчатки собаки было продемонстрировано ранее [Школьник-Яррос, 1968а]. Различные варианты их можно видеть также на рис. 21. Наблюдаются как густые кустики в наружном плексиформном слое, так и единичные терминали, либо широковетвящиеся дендритные веточки. Гольджи-электронно-микроскопические исследования биполярных клеток сетчатки собаки пока отсутствуют, поэтому суждения о типах нейронов преждевременны.
В сетчатке приматов обнаружены палочковые биполярные клетки, плоские колбочковые и карликовые двух разновидностей: инвагинирующие и плоские Boycott, Dowling, 1969; Kolb, Boycott, Dowling, 1969; Kolb, 1970]. Аксоны палочковых биполярных клеток характеризуются наибольшей величиной терминальных утолщений, расположенных вблизи от тел ганглиозных клеток. Иногда наблюдаются щелевые контакты между аксоном биполяра и телом ганглиозной клетки па периферии сетчатки [Dowling, Boycott, 1966]. У плоских биполярных клеток аксоны оканчиваются выше, чем у палочковых, никогда не доходя до уровня тел ганглиозных клеток.
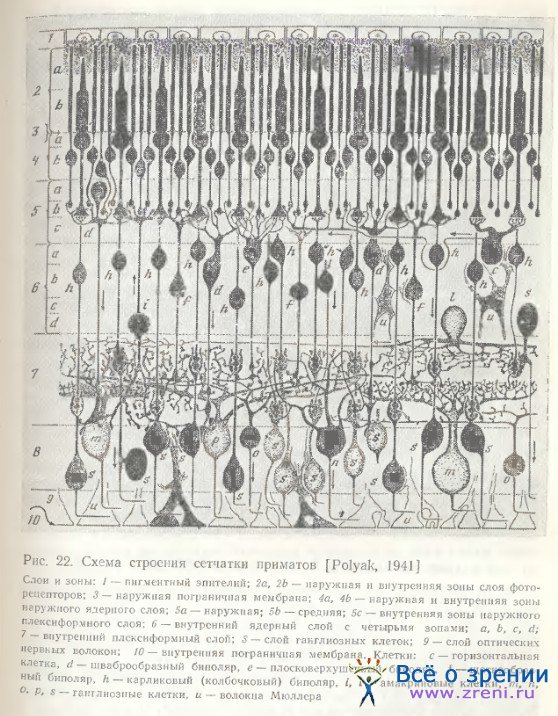
Карликовые биполярные клетки сетчатки приматов (рис. 22) впервые были описаны Polyak [1941]. Kolb [Kolb, Boycott, Dowling, 1969; Kolb, 1970] обнаружила, что они не единообразны, а образуют с колбочками синапсы различного рода: либо плоские, либо инвагинирующие. По данным Mariani [1981], в сетчатке приматов колбочковыми биполярными клетками являются не только карликовые нейроны, по и диффузные. Диффузные клетки, так же как и карликовые, наблюдаются двух разновидностей — инвагинирующие и плоские. Тонкие дендриты инвагинирующих диффузных колбочковых клеток, как показали Гольджи-электронно-микроскопические исследования, контактируют с 7 колбочками, образуя центральный элемент ленточного синаптического комплекса.
Аксонные терминали достигают внутренних отделов внутреннего плексиформного слоя (па уровне подслоя 4, по Ramon у Cajal [1972]), и этим отличаются от палочковых биполяров, аксон которых оканчивается ближе к слою ганглиозных клеток (в подслое 5, по Ramon у Cajal). Boycott, Dowling [1969] не подтвердили предположение Polyak [1941] о гинирующих биполярных клеток чаще всего располагаются в наружной зоне внутреннего ядерного слоя. Дендритные терминали их образуют центральные элементы в колбочковых ножках. Один такой нейрон контактирует с 4—10 колбочками При внутриклеточном отведении и окраске клетки пероксидазой хрена с последующим электронно-микроскопическим исследованием синаптических связей выявлены новые детали морфологии колбочковых биполярных клеток в сетчатке кошки [Nelson, Kolb, 1983]. Колбочковый биполяр, отвечающий гиперполяризацией на освещение, образует инвагинирующие ленточные синапсы с колбочками, а его аксон оканчивается во внутреннем отделе внутреннего плексиформного слоя, в его подслое b.
Аксонные терминали другого кол бочкового биполяра оканчиваются в наружном подслое а внутреннего плексиформного слоя, дендриты формируют базальные (поверхностные) синапсы с колбочками, а реакция па освещение также гиперполяризационная. У деполяризационных колбочковых биполярных клеток аксон оканчивается в подслое b, но контакты с колбочками происходят посредством полуинвагинирующего синапса. Авторы отказались от прежнего взгляда на деполяризационные колбочковые биполярные клетки как обязательно инвагинирующие в фоторецепторы и выделили новый тип связи — полуинвагинируюший синапс. Карликовые биполярные клетки, подобные найденным Polyak [1941], Boycott, Dowling [1969 , Kolb [1970] в сетчатке приматов, в сетчатке кошки не обнаружены.
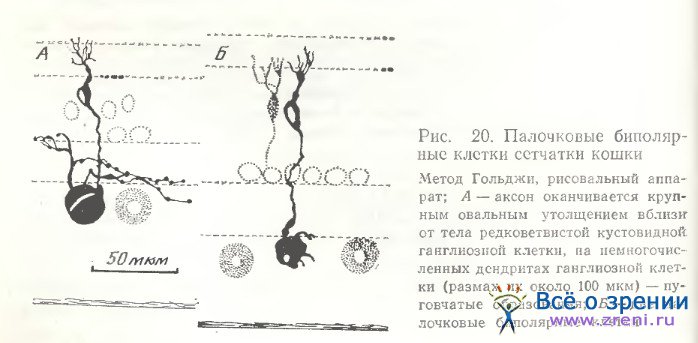
Биполярные клетки сетчатки кошки изучались также нами [Shkolnik-Yarros, 1971]. Описания палочковых и плоских колбочковых биполярных клеток, данные в 1971 г., не требуют изменений, так как вполне соответствуют более позднему детальному исследованию [Boycott, Kolb, 1973Ь]. Тела палочковых биполяров часто находятся в наружном отделе внутреннего ядерного слоя (рис. 20, А) либо в средней его части (рис. 20, Б). Дендриты отходят от тела либо несколькими тоненькими стволиками, сразу же переходящими в терминали, либо одним крупным основным стволом, рассыпающимся на небольшой веер вторичных дендритов в наружном плексиформном слое (рис. 20). На конце аксона часто наблюдается крупное утолщение, расположенное вблизи от тела ганглиозной клетки. По ходу основного дендрита у палочковых клеток иногда наблюдаются утолщения (см. рис. 20, Б). Плоские клетки отличаются внутренним низким расположением дендритных ветвлений в наружном плексиформном слое (см. рис. 10, b, f) и наружным расположением горизонтально направленных аксонов во внутреннем плексиформном слое. Размах как дендритных, так и аксонных ветвлений значительно шире, чем у палочковых клеток.
Как сказано выше, Boycott, Kolb [1973b] установили наличие третьего типа биполярной клетки в сетчатке кошки — инвагинирующей. Судя по характеру дендритных терминалей и расположению тела, вероятно, клетка на рис. 6, D.
В сетчатке у некоторых видов позвоночных обнаруживаются смещенные биполярные клетки, впервые детально описанные А. С. Догелем [Dogiel, 1885]. Многие авторы их видели, но не всегда правильно идентифицировали (см. обзор Т. В. Давыдовой [1981]). Особенно много смещенных биполярных клеток в сетчатке черепахи, где их тела образуют регулярно расположенный сплошной ряд в самом внутреннем отделе наружного ядерного слоя. Апикальный отросток у таких клеток выходит за пределы наружной пограничной мембраны, где оканчивается, образуя выступ, наполненный плотно упакованными мелкими вытянутыми митохондриями с ресничкой. По данным автора, наружный отросток смещенных биполярных клеток является колбой Ландольта (как это предполагал Ramon у Cajal), что подтверждено исследованием ультраструктуры.
Для понимания морфологических особенностей биполярных клеток у различных видов позвоночных следует учитывать разнообразие рецепторов, контактирующих с дендритными терминалями, а также сложность и дифференцированность внутреннего плексиформного слоя, с которыми связана длина, коллатеральные ветвления, пуговки по ходу и другие контактные структуры аксонов. У рыб, амфибий, рептилий, птиц фоторецепторы, а следовательно, и биполярные клетки более разнообразны, чем у млекопитающих, а многослойные ветвления аксонов биполярных клеток в сложно-дифференцированном внутреннем плексиформном слое наиболее выражены в сетчатке рептилий и птиц. Кроме этого, в сетчатке многих низших позвоночных от дендритов биполярных клеток отходит колба Ландольта, отсутствующая в сетчатке млекопитающих.
---
Статья из книги: Нейроны сетчатки | Е.Г. Школьник-Яррос, А.В. Калинина

Комментариев 0