Вегетативная (автономная) иннервация глаза │ Часть 1


Содержание:
Описание
↑ Вегетативная система и ее контроль центральной нервной системой
Вегетативная нервная система, иннервируя гладкую мускулатуру всех органов, кровеносных сосудов, сердце и железы, отвечает за регуляцию внутренней среды организма. Для офтальмолога наиболее важным является то, что она обеспечивает зрачковый рефлекс, аккомодацию, секреторную функцию слезной железы. Под ее контролем находится внутриглазное давление, функции различных структур глаза и глазницы.
Вегетативная (автономная) нервная система получила свое название в связи с тем, что ранее предполагали полное отсутствие контроля над ней со стороны коры головного мозга, поскольку функционирует она даже при нарушении связи между спинным и головным мозгом. Именно этим вегетативная нервная система отличается от произвольной, сознательно управляемой, соматической системы.
Наиболее высокими уровнями контроля деятельности вегетативной нервной системы являются стволовая часть мозга, гипоталамусе и лимбическая система. Эти структуры вовлечены в большинство жизненно важных «бессознательных» функций по обработке информации, поступающей от органов и тканей организма, и управлению их деятельностью. В свою очередь, стволовая часть мозга, гипоталамусе и лимбическая система находятся под волевым контролем коры головного мозга. Таким образом, понятие автономии вегетативной нервной системы довольно относительно.
О значении коры головного мозга и нижележащих структур в деятельности вегетативной нервной системы свидетельствует хотя бы такой факт. Стимуляция коры лобной, затылочной долей, а также стимуляция многих областей промежуточного мозга вызывает сужение или расширение зрачка.
Большую роль играет гипоталамус. Описано развитие синдрома Горнера после случайных повреждений гипоталамуса во время проведения стереотаксических операций. Стимуляция каудальной части гипоталамуса и серого вещества ствола приводит к расширению зрачка, в то время как их разрушение приводит к сонливости и сужению зрачка. О роли гипоталамуса в деятельности автономной системы свидетельствует и активация ее при сильном эмоциональном возбуждении. Кроме того, гипоталамусе обеспечивает надъядерное торможение зрачкового рефлекса, усиливающееся с возрастом.
Вегетативная нервная система существенно отличается от соматической и особенностями структурной организации. В первую очередь, она является двухнейронной системой. Один синапс образуется уже после выхода из центральной нервной системы в ганглиях, а второй синапс сформирован уже в эффекторном органе.
Следующее отличие сводится к тому, что соматическая нервная система формирует синапс (нейромышечный), обладающий достаточно стабильной структурой, в то время как синапсы автономной нервной системы представляют собой довольно разнообразные по строению структуры, диффузно распределенные на эффекторном органе.
В функциональном отношении обращает на себя внимание то, что если при стимуляции соматической нервной системы происходит возбуждение эффекторного органа (мышцы), то при стимуляции вегетативной могут наблюдаться как явления возбуждения, так и торможения.
В своей деятельности автономная нервная система использует большое количество различных типов нейромедиаторов и рецепторов.
Существуют различия и в функциональных проявлениях посттравматической регенерации вегетативных и соматических нервов. После денервации мышцы, иннервируемой автономной нервной системой, снижается мышечный тонус, но не наступает истинного паралича. В последующем нормальный тонус восстанавливается, а также возможно развитие гиперчуевствительности мышцы к медиаторам (ацетилхолинус для парасимпатической системы, норадреналинус для симпатической системы). Фармакологические механизмы гиперчувствительности при денервации симпатической и парасимпатической нервной системы различны. В первом случае определяется преюнкциональная, а во втором — постъюнкциональная гиперчувствительность. Преюнкциональная гиперчувствительность связана с утерей способности пресинаптическим аксоном поглощать избыток медиатора, в результате чего существенно увеличивается концентрация норадреналина в области синапса. Постъюнкциональная гиперчувствительность связана со структурно-функциональным изменением самой мышцы. При этом отмечается утеря специфичности рецептора к нейромедиатору.
В структурном отношении периферический отдел вегетативной нервной системы является исключительно эфферентным. Нейроны, расположенные в стволовой части мозга и спинном мозге, и их аксоны, направляющиеся к вегетативным ганглиям, называются преганглионарными нейронами. Нейроны, лежащие в вегетативных ганглиях, называются постганглионарными, так как их аксоны выходят из ганглиев и идут к исполнительным органам (рис. 4.5.1).
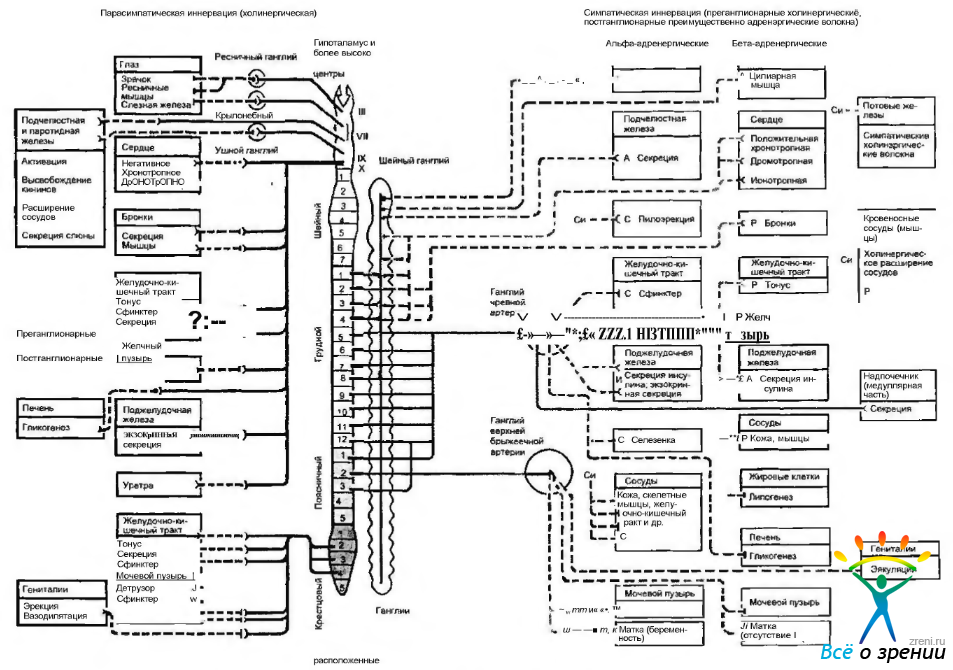
Рис. 4.5.1. Структурная и функциональная организация автономной нервной системы: а — активация; И — ингибирование; С — сокращение; Р - расслабление; Д — дилятация; Си — сегментная иннервация
Аксоны преганглионарных нейронов обладают миелиновой оболочкой. По этой причине их еще называют белыми нервными веточками. Аксоны постганглионарных нейронов немиелинизированы (серые веточки), за исключением постганглионарных аксонов, исходящих из ресничного ганглия. Направляясь к исполнительному органу , вегетативные нервы образу ют в их стенке густое сплетение.
Как было указано выше, периферическую часть вегетативной нервной системы подразделяют на два отдела — симпатическую и парасимпатическую. Центры этих отделов лежат на разных уровнях центральной нервной системы.
Многие внутренние органы получают как симпатическую, так и парасимпатическую иннервацию. Влияние этих двух отделов часто носит антагонистический характер, а нередко действуют «синергично». В физиологических условиях деятельность органов зависит от преобладания влияния той или иной системы. Основные структурные и функциональные особенности вегетативной иннервации органов и тканей человека представлены на рис. 4.5.1.
↑ Парасимпатическая система
Знание строения и функции парасимпатической нервной системы необходимо офтальмологу по ряду причин. Она обеспечивает аккомодацию и реакцию зрачка на свет, замедляет сердечную деятельность при воспроизведении окулокардиального рефлекса и мн. др.
Тела преганглионарных парасимпатических нейронов лежат в стволе мозга (ядра черепномозговых нервов, ретикулярная формация ствола) и в сакральном отделе спинного мозга (сакральные сегменты 2, 3 и иногда 4). От этих нейронов отходят миелинизированные и немиелинизированные аксоны, обладающие значительной длиной, которые в составе черепно-мозговых нервов направляются к постганглионарным парасимпатическим нейронам (рис. 4.5.1; 4.5.2).
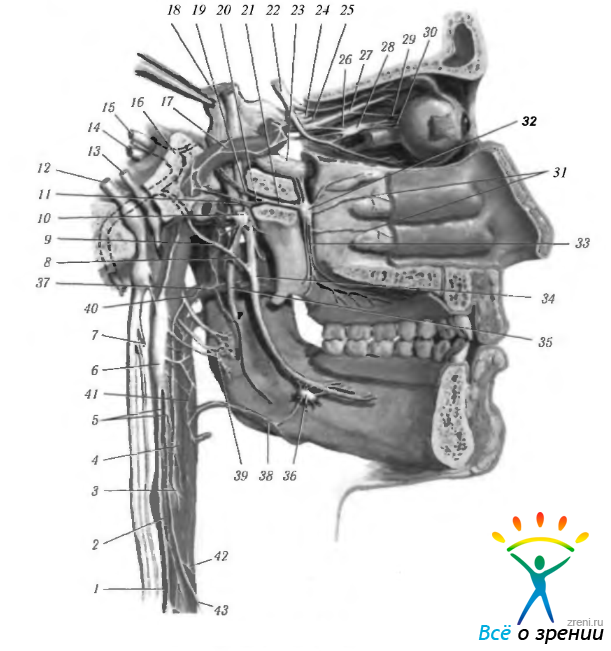
Рис. 4.5.2. Особенности организации автономной нервной системы головы (по Netter, 1997): 1 — верхняя шейная ветвь блуждающего нерва; 2 — шейный симпатический ствол; 3 — каротидный синус; 4 — ветвь языкоглоточного нерва; 5—внутренняя сонная артерия и сплетение; 6—верхний шейный симпатический ганглий; 7— верхний нерв гортани; 8 — барабанная струна; 9 — внутренний сонный нерв; 10 — ушной ганглий; 11 — нижнечелюстной нерв; 12 — блуждающий нерв; 13 — языкоглоточный нерв: 14 — статически-слуховой нерв: 15 -лицевой нерв; 16 — коленчатый ганглий: 17 — внутренняя сонная артерия и сплетение; 18 — тройничный нерв; 19 — большой каменистый нерв: 20 — глубокий каменистый нерв: 21 — нерв крыловидного канала (видиев); 22 — глазодвигательный нерв; 23 — верхнечелюстной нерв; 24 — глазной нерв; 25 — лобный и слезный нервы; 26 — носо-ресничный нерв; 27 — корешки ресничного ганглия; 28 — ресничный ганглий; 29 — длинный ресничный нерв; 30 — короткие ресничные нервы; 31 — задние латеральные носовые нервы; 32 — крылонебный ганглий; 33 — небные нервы; 34 — язычный нерв; 35 — нижний луночный нерв: 36 - поднижнечелюстной ганглий: 37 — средняя оболочечная артерия и сплетение; 38 — лицевая артерия и сплетение: 39 — сплетение гортани; 40 — верхнечелюстная артерия и сплетение; 41 — внутренняя сонная артерия и сплетение; 42 — общая сонная артерия и сплетение; 43 — верхний шейный симпатический нерв сердца
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Преганглионарные парасимпатические волокна, снабжающие внутриглазные мышцы и железы головы, покидают ствол мозга в составе трех пар черепно-мозговых нервов — глазодвигательного (III), лицевого (VII) и языкоглоточного (IX). К органам грудной и брюшной полости преганглионарные волокна идут в составе блуждающих нервов, а к органам полости таза в составе тазовых нервов подходят парасимпатические волокна крестцового отдела.
Парасимпатические ганглии расположены только в области головы и вблизи тазовых органов. Парасимпатические клетки других участков тела разбросаны на поверхности или в толще органов (желудочно-кишечный тракт, сердце, легкие), образуя интрамуральные ганглии.
В области головы к парасимпатическим ганглиям относятся ресничный, крылонебный, поднижнечелюстной и слуховой. Через перечисленные ганглии проходят также чувствительные и симпатические волокна (рис. 4.5.1, 4.5.2). Более подробно ганглии мы опишем несколько ниже.
Перед тем как изложить данные относительно анатомической организации парасимпатической системы в области головы и шеи, необходимо остановиться на нейромедиаторах этой системы.
Медиатором парасимпатической нервной системы является ацетилхолин, который высвобождается в окончаниях всех преганглионарных вегетативных волокон и большинстве постганглионарных парасимпатических нейронов. Действие ацетилхолина на постсинаптическую мембрану постганглионарных нейронов может быть воспроизведено никотином, а действие ацетилхолина на эффекторные органы — мускарином. В связи с этим возникла концепция о наличии двух типов рецепторов ацетилхолина, и влияние на них этого медиатора было названо никотиноподобным и мускариноподобным. Существуют препараты, избирательно блокирующие то или иное влияние. Никотиноподобное действие ацетилхолина на постганглионарные нейроны выключается четвертичными аммониевыми основаниями. Такие вещества называются ганглиоблокаторами. Мускариноподбный эффект ацетилхолина избирательно блокируется атропином.
Вещества, действующие на клетки эффекторных органов так же, как холинэргические постганглионарные парасимпатические нейроны, называются парасимпатомиметическими, а вещества, выключающие или ослабляющие влияние ацетилхолина на эти органы, называются парасимпатолитическими.
После деполяризации постсинаптической мембраны ацетилхолин удаляется из синаптической щели двумя путями. Первый путь сводится к тому, что ацетилхолин диффундирует в окружающие ткани. Второй путь характеризуется тем, что ацетилхолин подвергается гидролизу под действием ацетилхолинэстеразы. Образующийся при этом холин активно транспортируется назад в пресинаптический аксон, где участвует в синтезе ацетилхолина. Ацетилхолин гидролизуется не только специфическим энзимом — холинэстеразой, но и рядом других неспецифических эстераз, но этот процесс происходит вне синапсов (ткани, кровь).
Теперь мы подробно опишем анатомию основных образований парасимпатической системы в области головы.
Центральный путь парасимпатической системы. Центральный путь парасимпатической системы изучен недостаточно хорошо. Известно, что двигательные (центрифугальные) волокна идут от коры затылочной области по направлению предкрышечных ядер (nuclei pretectales) (ядро оливы, подчечевицеобразное ядро, ядро зрительного тракта, заднее и основное претектальное ядро; см. ниже). Об этом свидетельствует хотя бы тот факт, что стимуляцией коры затылочной области (поля 18, 19 и некоторые другие) можно индуцировать миоз. Этим же можно объяснить нарушение зрачкового рефлекса у больных с повреждением структур, расположенных выше наружного коленчатого тела.
Проецируются центральные пути первоначально на претектальную область, а затем на комплекс нейронов, включающий в свой состав ядро Якубовича—Эдингера—Вестфаля, переднее медиальное ядро и ядро Перлиа (рис. 4.5.5, 4.5.6. 4.5.11).
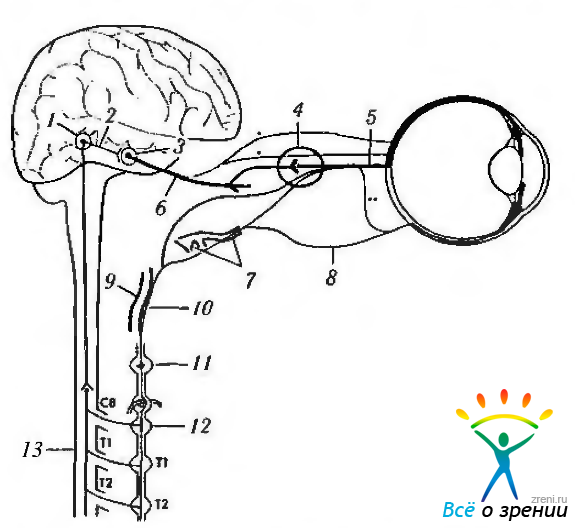
Рис. 4.5.5. Контроль автономной нервной системы центральной нервной системой: 1 — гипоталамический центр; 2 - симпатический ингибирующий путь; 3— ядро Якубовича—Эдингера—Вестфаля; 4 — ресничный ганглий; 5 короткие ресничные нервы; 6 - III нерв; 7 - носоресничный нерв; 8 — длинный ресничный нерв; 9 — тройничный ганглий; 10— сонное сплетение; 11 — верхний шейный ганглий; 12—нижний шейный ганглий; 13 — цилиоспинальный центр
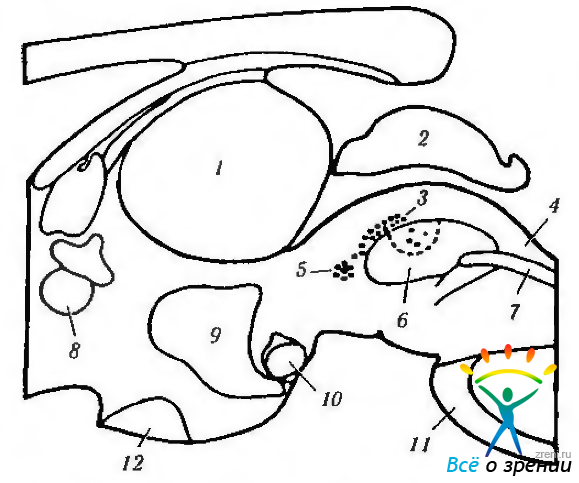
Рис. 4.5.6. Схематическое изображение локализации висцеральных ядер глазодвигательного нерва в дорзальной части среднего мозга (по Burde, Loewv, 1980): парасагиттальный срез, иллюстрирующий взаимоотношения срединного ядра (5), ядра Якубовича—Эдингера—Вестфаля (3) и ядра Перлиа (4) (1 — зрительный бугор; 2 — верхние бугорки; 3 — ядра Якубовича—Эдингера—Вестфаля; 4 — ядро Перлиа; 5 — срединные ядра; 6 — глазодвигательные ядра; 7 — медиальный продольный пучок; 8—передняя спайка; 9 - III желудочек; 10 — сосцевидное тело; 11 — мост; 12— зрительный перекрест)
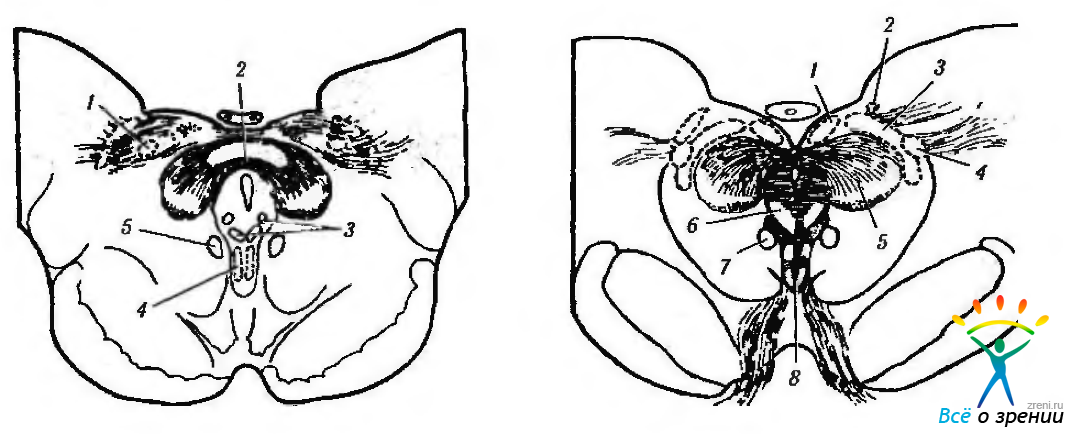
Рис. 4.5.11. Схематическое изображение локализации висцеральных ядер глазодвигательного нерва в дорзальной части среднего мозга (по Carpenter, Pierson, 1973): а — отношения переднего срединного ядра, ядра Якубовича— Эдингера- -Вестфаля с ядрами претектальной области (1 — ядро оливы: 2 — задняя спайка; 3 — латеральные и медиальные клеточные колонки: 4 — переднее срединное ядро: 5 — ядро Кахала). Ядро Якубовича—Эдингера—Вестфаля состоит из двух клеточных групп — латеральной и медиальной клеточных колонок. Переднее срединное ядро расположено непосредственно вентральней и рострально висцеральных клеточных колонок ядра Якубовича—Эдингера—Вестфаля; б — большое претектальное ядро и его отношение с передним срединным ядром (1 — область претектальных ядер; 2 — ядро зрительного тракта; 3 — подчечевицеподобное ядро; 4 —ядро оливы; 5 — ядро задней спайки; б — ядро Даршкевича; 7 — ядро Кахала; 8 — висцеральное глазодвигательное ядро)
Эти нейроны контролируют наиболее важные рефлексы глаза (зрачковый рефлекс, аккомодация и др.) До настоящего времени точно не установлена локализация нейронов, отвечающих за ту или иную функцию. Так, Jampel, Mindel обнаружили, что нейроны, обеспечивающие сужение зрачка, лежат вентральней и каудальней, чем клетки, отвечающие за аккомодацию. Однако Sillito, Sillito, Zbrozyna, Pierson, Carpenter утверждают, что суживающие зрачок нейроны располагаются ростральней ядра Якубовича—Эдингера—Вестфаля.
Использование иммуноморфологических методов выявило, что афференты зрачкового рефлекса идут от ядра задней спайки, которое, в свою очередь, получает афференты от претектальной области противоположной стороны (рис. 4.5.11). Предполагают, что ядро задней спайки является образованием, объединяющим как симпатические, так и парасимпатические входы зрачкового рефлекса. При этом оно получает афференты от претектальной области и отдает эфференты в направлении спинного мозга и ядра Якубовича—Эдингера—Вестфаля.
Тормозные (расширяющие зрачок) входы к ядру Якубовича—Эдингера—Вестфаля направляются от гипоталамуса, спинно-таламических путей, парамедианной ретикулярной формации и вестибулярной системы.
Выявлено два нисходящих пучка волокон, идущих от ядра Якубовича—Эдингера—Вестфаля. Первый пучок называется латеральным путем. Использует он покрышечно-спинномозговой тракт. Этот тракт проецируется на спинной мозг (рис. 4.3.3). Второй путь (средний путь) проецируется на заднее добавочное ядро оливы (nucleus olivaris accessorius posterior).
Аксоны нейронов ядра Якубовича—Эдингера—Вестфаля формируют парасимпатические волокна, направляющиеся к ресничному ганглию (рис. 4.5.2; 4.5.5).
Помимо ядра Якубовича—Эдингера—Вестфаля парасимпатическую иннервацию обеспечивают также нейроны верхнего слюноотделительного ядра (nucleus salivarius superior), аксоны которых в составе лицевого нерва направляются к крылонебному и подчелюстному ганглиям. Аксоны нижнего слюноотделительного ядра (nucleus salivarius inferior) формируют волокна, направляющиеся в составе языкоглоточного нерва к ушному ганглию (ganglion oticum) (рис. 4.5.2).
Ресничный ганглий (g. ciliare). После выхода из центральной нервной системы парасимпатические волокна направляются к ресничному ганглию по глазодвигательному нерву (рис. 4.5.5).
Ресничный ганглий расположен в глазнице в мышечной воронке вблизи глазного яблока (рис. 4.5.2). Размеры и форма его разнообразны, но локализация постоянна.
Большинство пупилломоторных и аккомодационных волокон в момент выхода глазодвигательного нерва из среднего мозга располагаются на дорзальной поверхности нерва. При гистологическом исследовании парасимпатические волокна отличаются от соматических волокон своим небольшим диаметром. Расположение их с дорзомедиальной стороны нерва объясняет рано развивающееся расширение зрачка при развитии патологии этой области, приводящей к сдавлению нерва.
В области турецкого седла пупилломоторные волокна лежат в центре нерва, а в глазнице они обнаруживаются только в нижней ветви глазодвигательного нерва. Именно по ней они направляются к нижней косой мышце и поступают в ресничный ганглий.
Помимо парасимпатических волокон ресничный ганглий содержит и симпатические волокна, поступающие из симпатического сплетения внутренней сонной артерии (рис. 4.5.5). Имеются и чувствительные волокна. Чувствительный (сенсорный) корешок ресничного ганглия присоединяется к носо-ресничной ветви тройничного нерва. Возможны и прямые связи между короткими ресничными и носо-ресничным нервом, обходящие ганглий.
Из ресничного ганглия постганглионарные мякотные волокна в составе коротких ресничных нервов проникают в глазное яблоко и проходят к сфинктеру радужной оболочки и ресничной мышце (рис. 4.5.2).
Некоторые из парасимпатических волокон остаются преганглионарными, т. е. проходят через ресничный ганглий без образования в нем синапсов. Эти волокна образуют синапсы с ганглиозными клетками, в большом количестве диффузно распределенными на внутренней поверхности ресничной мышцы. Электронномикроскопически и гистохимически выявлено, что некоторые из парасимпатических волокон заканчиваются на волокнах дилятатора радужки и, возможно, обладают тормозной функцией. И, наоборот, тормозные симпатические волокна выявлены на сфинктере.
Необходимо напомнить и то, что короткие ресничные нервы также обеспечивают парасимпатическую иннервацию сосудистой оболочки глаза, но уже благодаря волокнам, идущим от крылонебного ганглия (см. ниже).
Необходимо остановиться на покрышечно-спинномозговом (бульварном) тракте парасимпатической системы. Преганглионарные волокна этого тракта исходят из небольших нейронов слюноотделительного ядра, которое лежит вблизи дорзального ядра блуждающего нерва в колонке висцеральных эфферентных ядер III, VII, IX и X внутричерепных нервов. Общепринято делить это ядро на верхнюю и нижнюю части.
Верхнее слюноотделительное (и слезное) ядро располагается в ретикулярной формации ствола мозга каудальней ядра лицевого нерва и довольно близко к ядру блуждающего нерва (рисю 4.5.7).
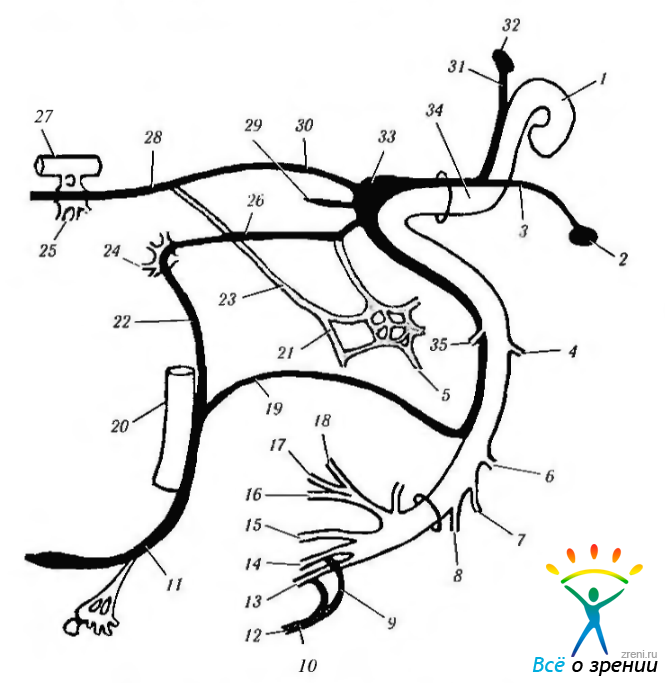
Рис. 4.5.7. Распределение вегетативных нервов: 1 — ядро лицевого нерва; 2 - ядро отдельного тракта; 3— афферентная ветвь промежуточного нерва; 4 — ушная ветвь блуждающего нерва; 5 — барабанная ветвь IX нерва; 6 — задняя ушная ветвь; 7 — к двубрюшной мышце; 8— к шило-подъязычной мышце; 9 — большой ушной; 10— шейное сплетение; Р — эфферентные волокна к подчелюстному и подъязычному ганглию и железам; 12— поперечный шейный; 13 — шейный; 14 — нижнечелюстной; 15 — щечный; 16 — подглазничный; 17 — челюстной; 18 — височный; 19 — барабанная струна; 20 — язычный нерв; 21 - барабанное сплетение; 22 — соединительная ветвь; 23 — большой глубокий каменистый нерв; 24 — ушной ганглий; 25 — крылонебный ганглий; 26 — малый поверхностный; 27 — верхняя ветвь верхнечелюстного нерва; 28 — видиев нерв; 29 - - наружный поверхностный каменистый; 30 — большой поверхностный каменистый; 31 эфферентная ветвь промежуточного нерва; 32 — верхнее слюноотделительное ядро; 33 — коленчатый ганглий; 34 — промежуточный нерв: 35 — к стременной мышце
Нейроны формируют секреторные волокна, которые покидают мозг в виде одного из компонентов лицевого нерва — промежуточного нерва (neruus intermedws). Этот нерв является смешанным нервом и несет вкусовые и чувствительные волокна от передних двух третей языка. В его состав также входят афферентные волокна от мимических мышц, твердой мозговой оболочки и сосудов средней черепной ямки.
Один из двух существующих путей характеризуется тем, что секреторные волокна покидают промежуточный нерв и присоединяются к барабанной струне (horda tympani), направляясь к поднижнечелюстному ганглию (ganglion submandibulare), а затем к подъязычной, передним язычным и подчелюстным слюнным железам (рис. 4.5.7).
Сосудорасширяющие волокна проходят первоначально по сосудам мозга, направляясь в большой каменистый нерв (n. petrous major) и каротидное сплетение (plexus caroticus internus) (рис. 4.5.7).
Секретомоторные волокна, распространяющиеся через большой каменистый нерв, в крылонебном ганглии (g. pterygopalatinum) образуют синапсы. Затем волокна проходят через коленчатый ганглий (gangl. geniculate) и через лицевой канал (canalis facialis) височной кости проникают в среднюю черепную ямку. Пройдя под тройничным ганглием они достигают слепого отверстия (foramen laсеrum). В фиброхрящевой части этого отверстия волокна соединяются с симпатическими волокнами глубокого каменистого нерва, отходящими от каротидного сплетения. При этом они образуют пере крыловидного канала (Виднее нерв), который заканчивается в крылонебном ганглии. Это место и является релейной станцией преганглионарных парасимпатических волокон (рис. 4.5.7).
Постганглионарные нервные ветви посредством скуловой ветви верхнечелюстного нерва направляются к слезной железе. В последние годы выявлены особенности иннервации слезной железы. Первоначально полагали, что постганглионарные волокна поступают в верхнечелюстной нерв (n. maxillaris) и распространяются со скуловой ветвью, вплоть до проникновения в слезную железу через скуловисочные веточки (ramus zygomaticotemporalis), идущие со слезным нервом. Однако Ruskell обнаружил слезные веточки, идущие к железе от сплетения, расположенного позади глаза (заглазничное сплетение) (рис. 4.5.6). В свою очередь это сплетение состоит из парасимпатических волокон, исходящих непосредственно из крылонебного ганглия. Более подробно с особенностями слезной рефлекторной дуги можно ознакомиться, изучив рис. 4.5.8.
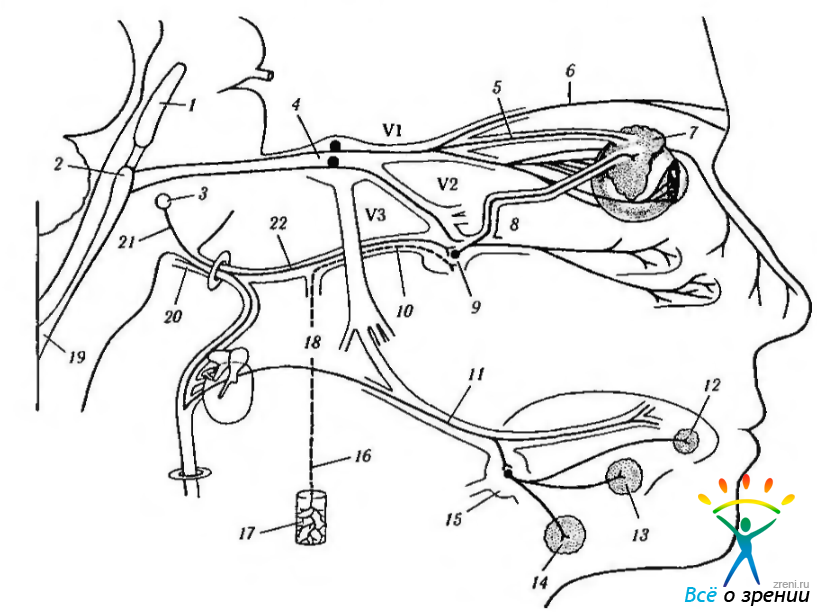
Рис. 4.5.8. Рефлекторная дуга слезной железы: 1 - среднемозговое ядро V нерва; 2 — главное чувствительное ядро V нерва; 3 — верхнее слюноотделительное ядро; 4 — тройничный ганглий; 5 — слезный нерв; 6 — лобный нерв; 7 — слезная железа; 8— заглазничное сплетение; 9 — крыловидный ганглий; 10— нерв крыловидного канала; 11 — язычный нерв; 12 — язычная железа; 13 — подъязычная железа; 14 — подчелюстная железа; 15 — подчелюстной ганглий; 16 — глубокий каменистый нерв; 17 — внутреннее сонное сплетение; 18 — chorda tympani; 19 — ядро спинномозгового тракта V нерва; 20 — VIII нерв; 21 — VII нерв; 22 — большой каменистый нерв. Афферентный путь формируется первой и второй ветвями тройничного нерва. Эфферентный тракт начинается в слезном ядре расположенного вблизи слюнного ядра, проходит по лицевому нерву, через коленчатый ганглий, большой поверхностный каменистый нерв и нерв крыловидного канала (где он соединяется с симпатическими волокнами глубокого каменистого нерва). Нерв проходит крыловидный ганглий, где он образует синапсы с третьим нейроном. Затем волокна вступают в верхнечелюстной нерв. Слезная железа иннервируется посредством волокон ретро-орбитального сплетения, образованного ветвями верхнечелюстного нерва. Они несут парасимпатические и VIP-эргические волокна
Крылонебный ганглий (g. pterygopalatinum). Крылонебный ганглий представляет собой небольшое образование (3 мм), расположенное в крылонебной ямке. Нейроны ганглия дают начало исключительно постганглионарным парасиматическим волокнам. В ганглии различают три корешка (рис. 4.5.2, 4.5.4, 4.5.8):
- Парасимпатический корешок от нерва крыловидного канала, дающий волокна структурам носоглотки.
- Симпатический корешок от нерва крыло видного канала, несущий преганглионарные симпатические волокна. При этом прерывания волокон в ганглии нет.
- Чувствительный, наиболее мощный корешок. Несет он веточку от верхнечелюстного нерва, а также афференты от слизистой полости носа, языка, неба, носоглотки, включая вкусовые волокна, предназначенные главному чувствительному ядру и спинальному ядру тройничного нерва.
Наиболее важными для офтальмолога ветвями, исходящими из ганглия, являются следующие:
- к слезной железе (парасимпатическая) (рис. 4.5.8);
- к мышце Мюллера глазницы (симпатическая);
- к надкостнице;
- веточка к ресничному ганглию, оболочкам зрительного нерва, отводящему и блоковому нервам, задней решетчатой и клиновидной пазухам:
- к глазной артерии и ее ветвям;
- к сосудистой оболочке.
При этом парасимпатические волокна достигают глазной артерии и сосудистой оболочки посредством ветвей, исходящих из заглазничного (ретро-орбитального) сплетения. В состав заглазничного сплетения входят также симпатические волокна, исходящие из сплетения внутренней сонной артерии (рис. 4.5.8).
От заглазничного сплетения отделяется 4—6 волокон (глазные веточки), которые проходят вперед по ходу глазодвигательного нерва и поступают в глазницу через верхнеглазничную щель. Эти волокна вплотную прилегают к глазной артерии и разветвляются. Затем они распределяются между ресничными артериями и проникают в глаз.
Хотя сплетение смешанное, глазные веточки состоят практически полностью из пучков безмякотных постганглионарных парасимпатических волокон, исходящих из крылонебного ганглия. Несколько глазничных веточек (rami orbitale) от крылонебного ганглия обходят заглазничное сплетение и иннервируют глазное яблоко непосредственно. Другие волокна от глазного сплетения (их rami vasculares) распределяются между ветвями глазной артерии.
Особенности иннервации артерий глазницы. Все артерии глазницы иннервируются веточками, исходящими от глазного сплетения (rami vasculares). Подходят они первоначально к адвентиции сосудов, а затем проникают до средней оболочки. Некоторые нервы берут свое начало из глазных веточек (rami oculare).
Нервы артерий содержат от 10 до 60 аксонов. Приблизительно 9,8% терминалов аксонов, найденных в стенках ресничных артерий, относятся к симпатическим (сосудосуживающим), поскольку после ганглиэктомии шейного узла наблюдается их дегенерация. Другие терминалы аксонов подвергаются дегенерации после ганглиоэктомии крылонебного узла, что свидетельствует об их парасимпатическом происхождении.
Крылонебный ганглий и регуляция внутриглазного давления. Многочисленными исследованиями показано, что после травмы крылонебного ганглия, его удаления или нейрэктомии каменистого нерва снижается внутриглазное давление. Это явление связывают с повреждением парасимпатических нервов, иннервирующих сосудистую оболочку. Указанные нервы исходят из глазных веточек (rami oculare). Основной их функцией является расширение просвета кровеносных сосудов сосудистой оболочки глаза.
Нижнее слюноотделительное ядро (n. salivatorius inferior) также относится к покрышечно-спинномозговому пути. Обеспечивает оно иннервацию околоушной железы и расположено в нижнем отделе ромбовидной ямки. В составе барабанной ветви языкоглоточного нерва секреторные волокна направляются к малому каменистому нерву, образуют синапсы в ушном ганглии (g. oticum) и лишь затем поступают в околоушную железу.
Заднее ядро блуждающего нерва (n. dorsalis nervi vagi). Заднее ядро блуждающего нерва лежит в продолговатом мозге в проекции дна ромбовидной ямки (треугольник блуждающего нерва). Двигательные волокна, возникающие в дорзальном ядре блуждающего нерва, оканчиваются в стенках сердца, легких и кишечника. Основные функции парасимпатической иннервации приведены на рис. 4.5.1.
↑ Симпатическая система
Тела преганглионарных нейронов симпатической системы располагаются в боковых рогах грудных и поясничных областей спинного мозга и покидают его в виде белых (миелинизированных) соединительных веточек (рис. 4.5.5, 4.5.9). Нейроны моторных постганглионарных волокон лежат в ганглиях по бокам позвоночника в виде цепи, а также в периферических ганглиях. Постганглионарные волокна безмякотные.
Медиатором преганглионарных волокон является ацетилхолин, а постганглионарных норадреналин. Исключением из этого правила являются симпатические волокна, иннервирующие потовые железы (ацетилхолин; холинергическая иннервация).
Поскольку в окончаниях симпатических постганглионарных нейронов выделяется норадреналин, эти нейроны называются адренергическими. Клетки мозгового слоя надпочечников, гомологичные постганглионарным симпатическим нейронам, высвобождают в кровоток главным образом адреналин. Как норадреналин, так и адреналин принадлежит к катехоламинам.
Существуют вещества, воспроизводящие действие симпатических адренергических нейронов (симпатомиметики) либо блокирующие это действие (симпатолитики).
Реакции различных органов на норадреналин и адреналин, так же как на ацетилхолин и другие медиаторы, опосредованы взаимодействием катехоламинов с особыми образованиями клеточных мембран, называемыми адренорецепторами. Благодаря фармакологическим исследованиям выделены альфа- и бета-адренорецепторы. Сущность фармакологических различий между двумя типами рецепторов можно найти в учебниках по физиологии и фармакологии. Практическому врачу необходимо знать, что в большинстве органов содержатся как альфа-, так и бета-рецепторы. Эффект возбуждения этих двух типов рецепторов, как правило, бывает противоположным, что необходимо помнить при применении различных фармакологических препаратов в лечении ряда глазных болезней.
В отличие от ацетилхолина, катехоламины после выполнения своей деполяризующей функции инактивируется иным образом. Существует два фермента, инактивирующих катехоламины. Первым является моноаминооксидаза (МАО), обнаруживаемая в большем количестве в нервных терминалах. Второй фермент называется катехол-О-метил трансфераза. Этот фермент обнаруживается только в постсинаптической мембране.
Симпатическая система иннервирует дилятатор радужной оболочки, гладкую мышцу Мюллера глазницы. Кроме того, она снабжает сосудосуживающими волокнами сосуды глаза и глазницы, а также иннервирует потовые железы и мышцу, поднимающую волосы кожи лица, и другие структуры.
Центральный путь. Центральный путь симпатической нервной системы начинается в заднем отделе гипоталамуса и проходит через ствол мозга, завершаясь в спинном мозге (рис. 4.5.5, 4.5.9).
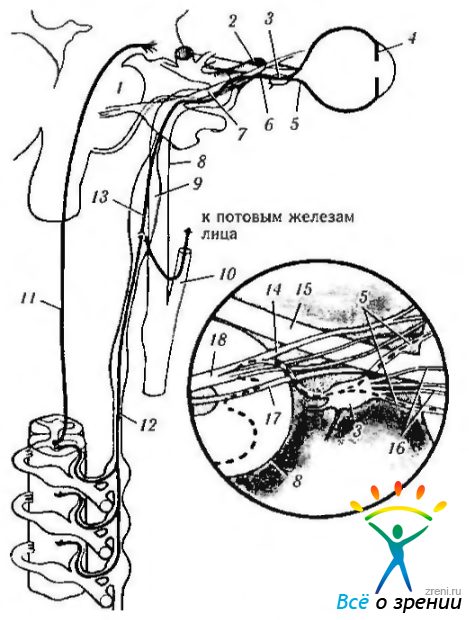
Рис. 4.5.9. Симпатическая иннервация глаза: 1 — мост; 2 - верхнеглазничная щель; 3 - ресничный ганглий; 4 — радужка; 5 — длинный ресничный нерв; 6 — носо- ресничная ветвь и VI; 7—первая ветвь тройничного нерва; 8—внутренняя сонная артерия; 9—верхний шейный симпатический ганглий; 10— наружная сонная артерия; 17 - первый нейрон; 12 — второй нейрон (преганглионарный); 13— третий нейрон (ностганглионарный); 14 — носо-ресничный нерв; 15 — зрительный нерв; 16 — короткие ресничные нервы; 17 — VI нерв; 18 — глазной нерв
В среднем мозге его волокна находятся с вентральной стороны и недалеко от срединной линии. В мосту волокна проходят вентральней серого вещества. На уровне нижней ножки мозга симпатические волокна лежат вентральней латерального спинно-таламического пути (tractus spinothalamicus lateralis). В продолговатом мозге волокна проходят через вентральную часть ретикулярной формации и спускаются в спинной мозг.
В спинном мозге симпатические волокна выявляются в одном миллиметре от переднебокового столба. Возможно частичное перекрещивание волокон в перекресте Фореля, расположенного по нижней границе среднего мозга. Часть симпатических волокон направляется к парасимпатическому ядру Якубовича—Эдингера—Вестфаля.
Нисходящие симпатические волокна располагаются в боковом канатике дорзомедиально и заканчиваются в боковом промежуточном столбе (coliimna intermediolateralis) (цилиоспинальный центр). При этом небольшое количество волокон перекрещивается (рис. 4.5.5, 4.5.9). Повреждение спинного мозга в месте прохождения симпатических волокон (ишемический инфаркт при синдроме Валленберга (Wallenberg), тромбоз задненижней мозжечковой артерии) приводит к развитию синдрома Горнера.
Преганглионарные волокна. Преганглионарные симпатические волокна возникают в нейронах бокового промежуточного столба, расположенных в боковых рогах спинного мозга в месте соединения грудных и шейных областей (так называемый «центр дилятатора») (а иногда С8 и С14). Эти волокна оставляют спинной мозг вместе с двигательными корешками и спинными нервами (рис. 4.5.2, 4.5.5).
К глазному яблоку волокна направляются преимущественно от первого грудного сегмента (Т.). Описаны больные, у которых после пересечения корешка Т, не развивался синдром Горнера. По этой причине предполагают, что некоторые пупилломоторные волокна берут свое начало в сегменте С8 или Т2.
Покидая спинной мозг, волокна спускаются по шейному стволу к верхнему шейному ганглию (ganglion superius), где и образуют синапсы с постганглионарными нейронами. При этом они проходят через нижний и средний шейные ганглии без образования в них синапсов (рис. 4.5.9). Palumbo на основании изучения больных после перенесенной симпатэктомии выявил, что симпатические пупилломоторные волокна покидают вентральные корешки С8, Т1 Т2 сегментов, проходят отдельным паравертебральным путем до нижнего или звездчатого ганглия.
Симпатические ганглии (рис. 4.5.2). Звездчатый ганглий (g. stellatum) образован слиянием первого грудного ганглия с двумя шейными ганглиями (слияние встречается в 30—80% случаев). Ганглий лежит рядом или латеральней боковой границы длинной мышцы шеи между поперечным отростком седьмого шейного позвонка и шейки первого ребра. При этом он находится позади позвоночной артерии, снизу отделен от плевры супраплевральной мембраной. По этой причине симпатический ствол нередко повреждается при развитии опухоли верхушки легкого. Следствием этого может возникнуть преганглионарный синдром Горнера, синдром Панкоаста (Pancoast; сочетание синдрома Горнера с каузалгическими болями в верхней конечности и груди с той же стороны, параличом мышц и гипо- или анестезией предплечья). Ганглий отдает веточки сплетению позвоночной артерии.
Средний шейный ганглий (g. cervicale medium) сформирован слиянием пятого и шестого шейных ганглиев и находится на уровне шестого шейного позвонка. Связан он со звездчатым ганглием.
Верхний шейный ганглий (g. cervicale superius) является самым большим (2,5 см) и находится на уровне второго и третьего шейных позвонков, вблизи их поперечных отростков. Этот ганглий образован слиянием ганглиев первых трех, а иногда и четырех шейных сегментов. Он отдает серые (постганглионарные) соединительные веточки нервным корешкам С3 и С4.
Близкое расположение верхнего шейного ганглия с внутричерепными нервами объясняет их одновременное поражение при травме или воспалительных заболеваниях основания черепа, а также ретропаротидного пространства.
Ганглий содержит холинергические преганглионарные и адренергические постганглионарные терминалы, а также катехоламинсодержащие хромафинные клетки, аминергические постганглионарные волокна.
↑ Постганглионарные волокна
Симпатические волокна области глазницы и глаза. Внутренний сонный нерв (п. caroticus internus) сопровождает внутреннюю сонную артерию в полости черепа проходя через каротидный канал. Нерв образует внутреннее сонное сплетение, вплотную прилежащие к артерии на всем ее протяжении (рис. 4.5.2).
Внутреннее сонное сплетение сформировано на боковой стороне артерии вблизи верхушки каменистой кости. Волокна от этого сплетения распределяются различным способом. Самый большой компонент симпатического сплетения на небольшом протяжении присоединяется к отводящему нерву. В последующем волокна сопровождают глазной нерв, а затем и носо-ресничный нерв (рис. 4.5.2, 4.5.5, 4.5.9).
Наиболее важными его ветвями являются:
- Ветвь к нерву крыловидного канала, которая достигает крылонебного ганглия посредством глубокого каменистого нерва. Волокна пересекают ганглий без образования синапсов и достигают глазницы через нижнеглазничную щель. Снабжают нервные волокна мышцу Мюллера орбиты, а также, возможно, слезную же лезу, сопровождая скуловой нерв (рис. 4.5.8).
- Ветви, направляющиеся к ветвям глазной артерии, включая слезную артерию, а также к отводящему (VI) нерву.
- Сонно-барабанные нервы в задней стенке каротидного канала, которые присоединяются к барабанной ветви языкоглоточного нерва. Они формируют барабанное сплетение. После про хождения через барабанное сплетение симпатические волокна снова включаются в сонное сплетение (рис. 4.5.8).
Пещеристое сплетение (plexus cavemosus). Пещеристое сплетение лежит на нижнемедиальной поверхности сонной артерии в области пещеристой пазухи. Ветви, исходящие из пещеристого сплетения, иннервируют глазное яблоко и почти всю глазницу. В пределах пещеристой пазухи ветви симпатического сплетения распределяются между глазными, передними мозговыми, средними мозговыми и передними хориоидальными артериями. Задняя соединительная артерия, вероятно, получает волокна от внутреннего сонного и позвоночного симпатических сплетений.
Пещеристое сплетение отдает следующие ветви:
- Ветви к ганглию тройничного нерва (Гассера) и глазной ветви тройничного нерва. Нервные волокна распределяются в носо-ресничном нерве и через верхнюю глазничную щель поступают в глазницу, достигая глазного яблока в составе длинных ресничных нервов. Они образуют волокна, расширяющие зрачок. Иногда часть волокон достигает глаза вместе с короткими ресничными нервами.
- Небольшую ветвь ресничному ганглию, проникающую в глазницу через верхнеглазничную щель. Она может непосредственно присоединяться к ганглию в виде симпатического ко решка, а также объединяться с соединительной ветвью, идущей от носо-ресничного нерва. Эти волокна проходят через ресничный ганглий без прерывания и по ходу коротких ресничных не рвов достигают глазного яблока, обеспечивая его кровеносные сосуды сосудосуживающими волокнами (рис. 4.5.5, 4.5.9). Иннервируют они также стромальные меланоциты увеального тракта.
- Ветви к глазной артерии и ее ветвям, а также к глазодвигательному и блоковому нервам. Ветви, направляющиеся к глазодвигательному нерву, иннервируют мышцу Мюллера века.
Наружные сонные нервы (n. carotid externi). Постганглионарные симпатические волокна, предназначенные для иннервации структур лица, оставляют верхний полюс верхнего шейного ганглия и присоединяются к наружной сонной артерии, образуя вокруг нее сплетение. Эти наружные сонные волокна иннервируют потовые железы лица и мышцу, поднимающую волосы. Покидая кровеносные сосуды, они затем распределяются в конечных ветвях тройничного нерва.
Теперь мы кратко остановимся на основных «глазных» рефлексах симпатической нервной системы. Начнем с описания зрачкового рефлекса.
Продолжение в следующей статье: Вегетативная (автономная) иннервация глаза ? Часть 2
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0