Вегетативная (автономная) иннервация глаза │ Часть 2


Содержание:
Описание
↑ Зрачковый рефлекс
Размер зрачка определяется многими факторами. Это возраст, эмоциональное состояние, степень освещения сетчатки, степень аккомодации и др. Изменение диаметра зрачка контролируется действием парасимпатических и симпатических эфферентных трактов.
Зрачковый рефлекс заключается в содружественном и равном сужении зрачков при освещении одного из глаз, обеспечивая при этом уменьшение светового потока, падающего на сетчатку. Сужение зрачка выявляется при исключительно низкой интенсивности освещения и пропорционально интенсивности и продолжительности стимула.
Свет, проходя через преломляющие среды глаза, попадает на сетчатку. Фоторецепторы сетчатой оболочки и являются началом рефлекса. Парасимпатическая иннервация сфинктера является эфферентным плечом зрачкового рефлекса рефлекторной дуги.
Афферентный путь (рис. 4.5.10).

Рис. 4.5.10. Схема иннервации сфинктера и дилятатора радужной оболочки глаза: 1 — наружное коленчатое тело; 2 - плечо верхних бугорков; 3— верхние бугорки четверохолмия; 4 — задняя спайка; 5 — претектальная область; 6 — ядро оливы; 7 — глазодвигательный нерв; 8— ресничный ганглий; 9 — короткий ресничный нерв; 10— сфинктер радужной оболочки; 11 — переднее срединное ядро; 12— медиальная колонка соматических нейронов; 13— латеральная колонка соматических нейронов; 14— передне-боковой пучок волокон спинного мозга; 15 — цилиоспинальный центр; 16 — дентальная связка; 17 — вентральные корешки; 18 — соединительная веточка; 19 — первый грудной симпатический ганглий; 20 — нижний шейный симпатический ганглий: 21 — средний шейный симпатический ганглий; 22 — верхний шейный симпатический ганглий; 23 — симпатическое сплетение сонной артерии; 24 — глазная ветвь тройничного нерва; 25 — носо-ресничный нерв; 26 — длинный ресничный нерв; 27 — дилятатор радужной оболочки
Афферентный путь начинается в палочках и колбочках сетчатой оболочки и проходит в составе зрительного нерва к структурам центральной нервной системы. До сих пор обсуждается вопрос — «зрительные» и «зрачковые» волокна зрительного нерва идентичны или нет? Если даже волокна «зрачкового рефлекса» являются самостоятельными и не обеспечивают передачу зрительной информации, они все же располагаются вблизи волокон, несущих зрительную информацию. Об этом свидетельствуют факты исчезновения зрачкового рефлекса на слепом глазу (поражение зрительного нерва).
Проходя в зрительном нерве, зрачковые волокна достигают зрительного перекреста, где они частично перекрещиваются и часть их переходит на противоположную сторону.
Затем волокна поступают в зрительный тракт. Повреждение этой области приводит к развитию гемианопсической реакции зрачка Вернике (Wernicke).
В задней трети зрительного тракта, не достигая наружного коленчатого тела, волокна покидают зрительный тракт и проходят поверхностно в составе ручки верхнего бугорка по направлению к латеральной части верхнего холмика четверохолмия (рис. 4.5.10). Разрушение обеих ручек верхнего холмика приводит к тому, что зрачок не реагирует при освещении обоих глаз.
Ни одно из волокон зрачкового рефлекторного пути, по-видимому, не заканчивается в наружном коленчатом теле. Однако некоторые исследователи полагают, что возможно переключение части волокон, идущих в претектальную область, в прегеникулярном ядре, хотя морфологическими методами существование подобных связей не установлено.
В последующем «зрачковые» волокна проходят к среднему мозгу по боковой поверхности верхнего четверохолмия и достигают парного претектального ядра (плохо очерченное скопление мелких клеток, расположенных впереди латерального края верхнего четверохолмия). Здесь волокна прерываются, образуя терминалы (рис. 4.5.10, б).
Многочисленные подгруппы нейронов относят к претектальным ядрам, хотя их функциональное значение не совсем ясно. К ним относят
- ядро оливы,
- подчечевицеобразное ядро,
- ядро зрительного тракта,
- заднее ядро
- и предкрышечное ядро (рис. 4.5.11).
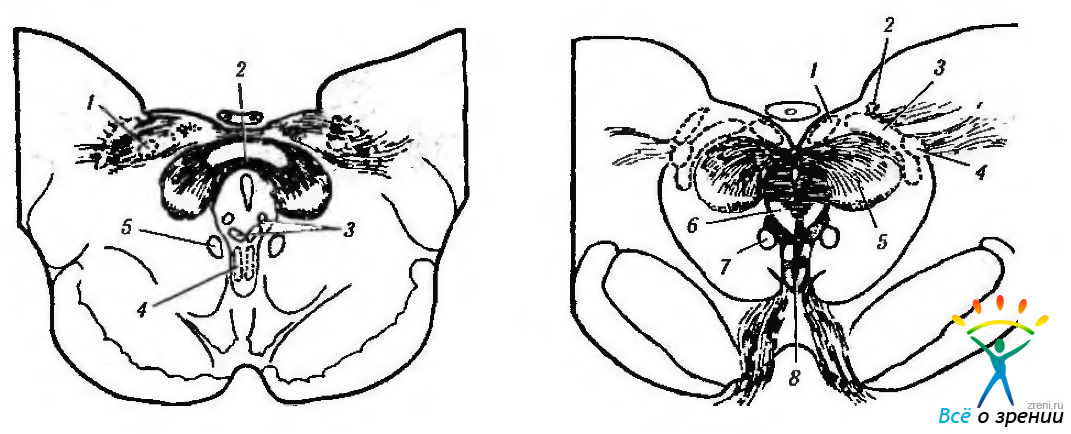
Рис. 4.5.11. Схематическое изображение локализации висцеральных ядер глазодвигательного нерва в дорзальной части среднего мозга (по Carpenter, Pierson, 1973): а — отношения переднего срединного ядра, ядра Якубовича— Эдингера- -Вестфаля с ядрами претектальной области (1 — ядро оливы: 2 — задняя спайка; 3 — латеральные и медиальные клеточные колонки: 4 — переднее срединное ядро: 5 — ядро Кахала). Ядро Якубовича—Эдингера—Вестфаля состоит из двух клеточных групп — латеральной и медиальной клеточных колонок. Переднее срединное ядро расположено непосредственно вентральней и рострально висцеральных клеточных колонок ядра Якубовича—Эдингера—Вестфаля; б — большое претектальное ядро и его отношение с передним срединным ядром (1 — область претектальных ядер; 2 — ядро зрительного тракта; 3 — подчечевицеподобное ядро; 4 —ядро оливы; 5 — ядро задней спайки; б — ядро Даршкевича; 7 — ядро Кахала; 8 — висцеральное глазодвигательное ядро)
Волокна, идущие от сетчатки, заканчиваются преимущественно в дорзомедиальной части ядра оливы (n. olivaris) с этой же стороны, а также в подчечевицеобразном ядре противоположной стороны (п. sublentiformis). Аналогичная проекция выявляется и на ядро предкрышечной области.
Аксоны нейронов ядра оливы и подчечевицеподобного ядра частично перекрещиваются в задней спайке, а также в вентральной части сильвиевого водопровода и направляются к «центру сфинктера» как с этой же стороны, так и на противоположную сторону, проходя через среднюю продольную связку. Число перекрещивающихся аксонов примерно такое же, как и неперекрещивающихся. Из-за симметрического перекреста волокон зрачки обоих глаз обычно одинакового размера. Физиологи предложили модель зрачковой функции, в которой каждый глаз производит сигнал, пропорциональный логарифму интенсивности света, а размер зрачка определяется в среднем мозге взвешенным арифметическим силы двух поступивших сигналов.
«Центр сфинктера» сформирован ядрами Якубовича—Эдингера—Вестфаля глазодвигательного нерва и нейронами претектального ядра. Многие клетки добавочных глазодвигательных ядер, если не их большая часть, обеспечивают механизмы аккомодации. Попытки отличать центры, сужающие и расширяющие зрачок, от «аккомодационных» до настоящего времени полностью успехом не увенчались.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Эфферентный путь. Аксоны добавочных глазодвигательных нейронов проходят в составе глазодвигательного нерва (III), располагаясь при этом на его дорзомедиальной поверхности. Отсюда волокна направляются медиально и вниз, поступают в нижнюю ветвь глазодвигательного нерва, с которым и проникают в глазницу (рис. 4.5.10). Большинство волокон в глазодвигательном нерве лежат поверхностно под эпиневрием.
От нижней ветви глазодвигательного нерва, посредством ветви, идущей к нижней косой мышце, волокна достигают ресничного ганглия. Эти мякотные преганглионарные парасимпатические волокна заканчиваются на теле и дендритах ганглиозных нейронов. Этот парасимпатический узел состоит из двух групп нейронов, меньшая из которых функционально связана с сужением зрачка, а большая — с процессом аккомодации.
Мякотные постганглионарные волокна покидают ресничный ганглий и поступают в глазное яблоко в составе коротких ресничных нервов. Большинство волокон этого пути (90%) направляется к ресничной мышце и только 3—5% к радужной оболочке. Остальные волокна иннервируют кровеносные сосуды и слезную железу. Приведенная рефлекторная дуга обеспечивает рефлекторное сужение зрачка при ярком освещении.
↑ Рефлекс при прекращении освещения глаза («темновой рефлекс»)
При прекращении освещения глаза развивается быстродействующая реакция — расширение зрачка. На начальной стадии эта реакция вызвана сокращением дилятатора радужки, а на поздней — подавлением функции сфинктера. Реализуется она через ядро Якубовича—Эдингера—Вестфаля и переднее срединное ядро.
Считают, что афферентный путь должен следовать вместе со зрительными волокнами к зрительному тракту. Дальнейший ход волокон к «центру, расширяющему зрачок», не изучен. Предполагают, что при отсутствии светового раздражения сетчатки возникает активное рефлекторное расширение зрачка. При этом сигналы могут передаваться в прегеникулярное ядро или в претектальную область, а далее в какую-либо часть ретикулярной формации среднего мозга. Последняя, в свою очередь, может передать сигналы преганглионарным симпатическим нейронам интермедиолатеральной группы нейронов спинного мозга (клеточная колонка на уровне 1—4-го грудных сегментов (Т, 4)). От симпатических нейронов импульсы передаются в ростральном направлении через симпатический ствол, волокна которого оканчиваются на постганглионарных клетках верхнего шейного узла (рис. 4.5.10). Эти постганглионарные клетки способны вызвать активное расширение зрачка благодаря своим связям с мышцей, расширяющей зрачок. Следует, однако, иметь в виду, что расширение зрачка может, по крайней мере частично, происходить пассивно просто из-за отсутствия раздражения, вызывающего его сужение.
Волокна, «ингибирующие» сужение зрачка, проходят через кортико-таламо-гипоталамический или кортико-лимбический пути и ингибируют парасимпатическую активность среднего мозга, в частности претектальных ядер. Подтверждением тому являются следующие данные. Стимуляция промежуточного мозга электрическим током после симпатэктомии у кошек и обезьян вызывает расширение зрачка и потерю светового рефлекса. Расширение зрачка было достигнуто также стимуляцией коры лобной области (область 8), затылочной доли и сенсомоторной коры. Эти данные указывают на вовлечение в процесс расширения зрачка гипоталамуса, который стимулирует дилятатор и тормозит сфинктер. При стимуляции гипоталамуса развивается мидриаз. поднимается веко и повышается кровяное давление. При этом расширение зрачка происходит даже у децеребрированных животных.
К быстрому расширению зрачка приводит также стимуляция довольно обширных областей лимбической системы, особенно поясной извилины (gyrus cinguli).
В стволовой части мозга выявлено два афферентных пути, которые приводят к расширению зрачка. Кегг проследил такие волокна от спинного мозга к глазодвигательным ядрам. Установлено, что восходящие волокна спинно-ретикулярного пути (tractus spinoreticularis) прямо тормозят мотонейроны, контролирующие сужение зрачка.
Нисходящие симпатические пути возникают в задних и боковых областях гипоталамуса и занимают боковое положение в стволе мозга. Имеются синапсы в мосту и покрышке.
Кегг и Brown электрофизиологически идентифицировали нисходящие пупилломоторные волокна у обезьян. Эти волокна располагаются поверхностно в передне-боковых колонках спинного мозга, занимая вентральное положение и образуя синапсы с преганглионарными нейронами, лежащими на уровне C8—Т2. Их стимуляция вызывает умеренный мидриаз с противоположной стороны. По этой причине считают, что существует перекрещивание волокон на уровне цилиоспинального центра. У человека подобные волокна не обнаружены.
↑ Конвергентно-аккомодационно-зрачковый рефлекс
При переводе взгляда от далеко расположенного предмета к предмету, лежащему вблизи, происходит одновременно сужение зрачка, аккомодация и конвергенция. Все три компонента рефлекса экспериментально воспроизводятся стимуляцией затылочной коры головного мозга. Необходимо отметить, что конвергентно-аккомодационно-зрачковый рефлекс (рис. 4.5.12, 4.5.13)
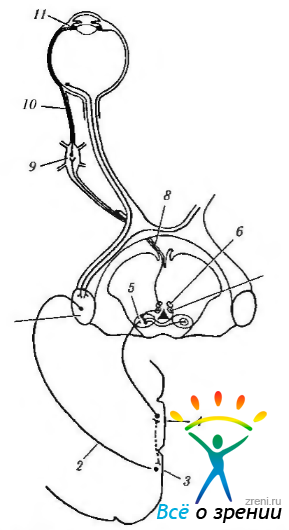
Рис. 4.5.12. Схема иннервации мышц ресничного тела: 1 — наружное коленчатое тело; 2 — зрительная лучистость; 3 — зрительная кора; 4 — перистриарная кора; 5 — претекталь-ная область; 6 — медиальная колонка висцеральных нейронов; 7 — латеральная колонка висцеральных нейронов; 8 — глазодвигательный нерв; 9 — ресничный ганглий; 10 — короткие ресничные нервы; 11 — мышцы ресничного тела
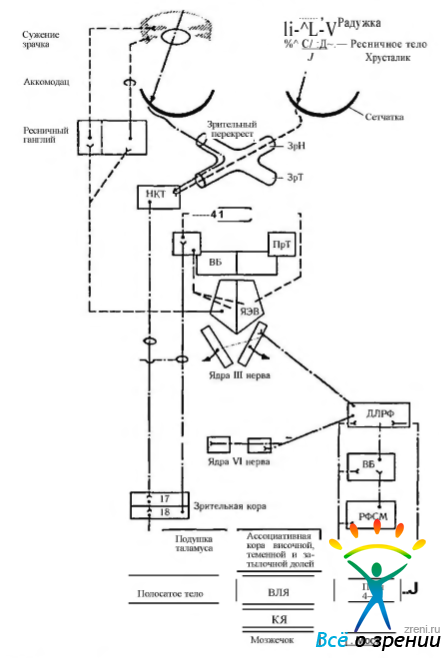
Рис. 4.5.13. Рефлекторные дуги зрачково-конвергентно-аккомодационного рефлекса (по Шаде, Форд, 1976): ЯЭВ — ядро Якубовича—Эдингера- -Вестфаля; НКТ — наружное коленчатое тело; ПрТ — претектальная область; ВБ — верхние бугорки четверохолмия; ЗК — задняя комиссура; ДЛРФ — дорзолатеральная ретикулярная формация; РФСМ — ретикулярная формация среднего мозга; ВЛЯ — вентролатеральное ядро зрительного бугра; КЯ — красное ядро
не является истинным рефлексом. Изменение размера зрачка, процессы аккомодации и сведение глазных яблок являются ассоциированным движением, обеспечиваемым надьядерными связями между нейронами, обслуживающими сфинктер, ресничную мышцу и наружные мышцы глаза. Об этом свидетельствует то, что зрачок сокращается при отсутствии изменения освещенности глаза.
При рефлекторной зрачковой реакции и аккомодации глаза импульсы первоначально возникают в небольшой по размерам области сетчатки, предположительно в районе центральной ямки. Раздражителем, вызывающим аккомодационный рефлекторный ответ, служит нарушение четкой фокусировки изображения на поверхности сетчатки, которое регистрируется зрительной корой.
Из сетчатки импульс передается по зрительному нерву и зрительному тракту к наружному коленчатому телу. При этом в области зрительного перекреста волокна от носовых полей сетчатки переходят на противоположную сторону, а волокна от височных отделов сетчатки остаются неперекрещенными.
Из наружного коленчатого тела сигнал передается в первичную зрительную кору (поле 17) и далее во вторичную зрительную кору (поле 18). Из поля 18 сигналы поступают в претектальную область через корково-покрышечный путь, где и образуют релейные синапсы (рис. 4.5.13).
Нейроны претектальной области устанавливают связь с ядром Якубовича—Эдингера— Вестфаля, эфференты которого являются преганглионарными волокнами, идущими в составе глазодвигательного (III) нерва к ресничному ганглию.
Ядро Якубовича—Эдингера—Вестфаля находится под ингибирующим действием ретикулярной формации моста. Усиление ингибирования приводит к расширению зрачка, а снижение — к сужению. Разрушение ингибирующих путей (например, при кровоизлияниях в области моста или сифилитическом поражении) вызывает сужение зрачка, которое сочетается с типичными клиническими признаками заболевания.
В последние годы получены данные, свидетельствующие о том, что волокна, обеспечивающие сужение зрачка при конвергентно-аккомодационно-зрачковом рефлексе проходят несколько иным путем, чем волокна, обеспечивающие зрачковый («световой») рефлекс. Полагают, что волокна конвергентно-аккомодационно-зрачкового рефлекса приближаются к претектальному ядру с вентральной стороны. Этим можно объяснить потерю реакции зрачка при освещении глаза прежде, чем разобщается конвергентно-аккомодационно-зрачковый рефлекс. Подобное состояние встречается при дорзальных инфильтративных (сдавливающих) повреждениях покрышки среднего мозга на уровне верхнего двухолмия (синдром Парино).
Аксоны нейронов ядра Якубовича—Эдингера—Вестфаля поступают затем в ресничный ганглий, оканчивась синапсами на клетках обоих его компонентов. Некоторые волокна заканчиваются на нейронах, иннервирующих ресничное тело, и вызывают сокращение его мышечных волокон. В результате этого уменьшается напряжение связки, поддерживающей хрусталик, что и приводит к изменению фокусного расстояния. Именно благодаря этому на сетчатке формируется четко сфокусированное изображение.
Другие преганглионарные волокна образуют связи с частью ресничного узла, обеспечивающей сужение зрачка.
Третьим компонентом реакции аккомодации является конвергенция глаз при рассматривании близко расположенных предметов. Первоначально полагали, что центры конвергенции находятся в среднем мозге, однако существование таких центров не было доказано. В настоящее время считается, что эта реакция осуществляется при участии корковых связей. Так, предполагают, что при этом реализуется связь между полем 18 зрительной коры с подушкой зрительного бугра (pulvinar thalami), которая, в свою очередь, посылает волокна в ассоциативные центры височной, теменной и затылочной долей.
Эти центры могут быть связаны с ретикулярной формацией среднего мозга, верхними бугорками и дорзолатеральными отделами ретикулярной формации. Благодаря таким связям с ретикулярной формацией сигналы могут идти к нейронам ядра глазодвигательного нерва, иннервирующим внутреннюю прямую мышцу глаза, а также к ядру отводящего нерва с целью торможения клеток, иннервирующих наружную прямую мышцу глаза.
Вторая возможная система связей может идти через полосатое тело к вентролатеральным ядрам зрительного бугра (nuclei ventrolaterales (thalami)). От этих ядер связи направляются к 4-му и 6-му полям коры головного мозга (моторная и премоторная области). Из этих полей коры импульсы через дорзолатеральные отделы ретикулярной формации могут достигать ядер черепно-мозговых нервов. В этом варианте возможны также связи через ядра моста с мозжечком и связи с красным ядром, что делает реальной координирующую роль мозжечка в реакции конвергенции.
Как указано выше, при конвергентно-аккомодационно-зрачковом рефлексе происходит фиксация, которая, по существу, складывается из аккомодации и сведения глазных яблок. Некоторые исследователи предполагают, что рефлекс начинается с конвергенции. При этом проприоцептивные импульсы от внутренней прямой мышцы направляются через глазодвигательный нерв или первую ветвь тройничного нерва к ядру среднемозгового пути тройничного нерва (nucleus mesencephalicus nervi trigeminalis (n. mesencephalicus trigemini)). Отсюда импульс направляется к ядру глазодвигательного нерва. После оставления глазодвигательного нерва импульс проходит через ресничный ганглий и проходит к сфинктеру радужки.
↑ Мигательный рефлекс
Этот рефлекс был описан в 1896 г. Overend. Сводится рефлекс к сокращению круговой мышцы глаза при механическом или электическом раздражении верхнего глазничного нерва первоначально со стороны раздражения, а несколько позже с двух сторон. Рефлекторная дуга замыкается на уровне ядер ствола мозга благодаря наличию интернейронов. Именно по этой причине возникает патологическое сокращение круговой мышцы глаза при заболеваниях ствола мозга, таких как синдром Валленберга.
↑ Цилиоспинальный рефлекс
Мидриаз. возникающий при болях в области шеи, развивается в результате подавления функции ядра Якубовича—Эдингера—Вестфаля. Известен этот рефлекс как цилиоспинальный рефлекс. Рефлекторный путь, объясняющий это явление, изучен Кегг.
Афферентные импульсы передаются в спинной мозг. Постсинаптические связи располагаются в дорзальном роге. Проходят они, главным образом, контрлатерально в наиболее поверхностном слое бокового столба. От этого уровня они поднимаются к стволу мозга и достигают ядра Якубовича—Эдингера—Вестфаля с двух сторон. По своему ходу эти волокна взаимодействуют и с другими структурами на уровне зрительного бугра или гипоталамуса.
В последнее время появились свидетельства того, что этот рефлекс может быть вызван не только возбуждением симпатических волокон, но и торможением парасимпатической системы.
↑ Нарушение зрачкового рефлекса в зависимости от уровня поражения
Сложный путь нервных волокон автономной нервной системы (парасимпатических и симпатических) в полости черепа, глазнице и их связь с различными черепно-мозговыми нервами, предопределяет развитие сложных сочетаний патологических изменений не только вегетативной иннервации, но также чувствительной и двигательной. Сочетание таких признаков складывется в синдромные поражения Наиболее рационально рассматривать их особенности в зависимости от уровня поражения (рис. 4.5.14).

Рис. 4.5.14. Особенности клинического проявления различных видов офтальмоплегии в зависимости от поражения вегетативной нервной системы (по Reeh et al., 1981): I - поражение афферентов зрительного нерва — нарушение зрачкового рефлекса при освещении глаза; II — поражение перекрещенных волокон зрительного перекреста — нарушение зрачкового рефлекса с двух сторон; III — зрительный факт — гомонимная гемианопсия, сопровождающаяся нарушением зрачкового рефлекса противоположного глаза (симптом Бэра (Веrr)) (а) или гемиакинезия зрачка (симптом Вернике (Wernicke)) (б); IV — верхние ручки — гемиакинезия зрачка; V — ядра претектальной области — зрачок Арджилля-Робертсона, реагирующего на приближение цели, но не реагирующего на свет; VI ядра задней спайки — симптомы, анологичные поражению ядер претектальной области или альтернирующая анизокория; VII — кортикотектальный тракт — надъядерный паралич конвергенции и аккомодации; VIII — поражение моста — миоз; IX — ядра Якубовича—Эдингера—Вестфаля— внутренняя офтальмоплегия (нарушение аккомодации и реакции на свет) или овальные зрачки; X — сдавливание дорзомедиальных волокон глазодвигательного нерва — внутренняя и наружная офтальмоплегия; XI — каудальное смещение ствола мозга, смещающее глазодвигательный нерв в направлении верхней мозжечковой артерии — полная офтальмоплегия; XII аневризма задней cоединительной артерии — болезненная офтальмоплегия; XIII — аневризма или спазм внутрипещеристой части сонной артерии — болезненная полная офтальмоплегия; XIV — синдром верхней глазничной щели — паралич симпатической и парасимпатической иннервации и раздражением ветвей глазного нерва и болью; XV — преганглионарные ветви ресничного ганлия - внутренняя офтальмоплегия; XVI — ресничные постганглионарные волокна — секторный паралич и/или тонический зрачок; XVII -фармакологическая блокада нейромышечных соединений (1 - кортикотектальный тракт: 2 — задняя спайка; 3 — верхние бугорки; 4 — претектальная область; 5 — наружное коленчатое тело; 6 — верхние ручки; 7 — ядро Якубовича—Эдингера—Вестфаля; 8 — зрительный тракт; 9— глазодвигательный нерв; 10— верхняя мозжечковая артерия; 11—задняя артерия мозга; 12—базилярная артерия; 13— задняя соединительная артерия; 14 — нижние ветви; 15 — ветвь к нижней косой мышце; 16 ресничный ганглий; 17 - диафрагма турецкого седла; 18 — внутренняя сонная артерия; 19 — средняя артерия мозга; 20 — передняя артерия мозга; 21 — зрительный нерв)
↑ Парасимпатическая система
Сетчатка и зрительный нерв. При поражении зрительного нерва и сетчатки (неврит, амблиопия) определяется амавротическая неподвижность зрачка. Зрачок слепой стороны слегка шире, чем другой. Отсутствует прямая и сохраняется непрямая реакция на свет. На видящем глазу сохраняется прямая, но отсутствует непрямая реакция на свет. Конвергенция сохранена на обоих глазах.
В тех случаях, когда на одном глазу развивается папиллит или ретробульбарный неврит, нарушение реакции зрачка на освещение является одним из наиболее важных признаков заболевания, даже при нормальной остроте зрения.
Зрительный перекрест. При поражении зрительного перекреста отмечается наличие гемианоптической неподвижности зрачков при наличии битемпоральной гемианопсии. При освещении носовых половин сетчаток выявляется двустороннее отсутствие прямой и непрямой реакции. При освещении височных половин сетчаток — живая прямая и непрямая реакция. Конвергенция сохранена.
Зрительный тракт. При поражении правого зрительного тракта у больных развивается гемианопическая левосторонняя неподвижность зрачков на свет при гомонимной левосторонней гемианопсии. Отсутствует прямая и непрямая реакция на свет при освещении слева. При этом сохраняется прямая и непрямая реакция при освещении справа.
Поражение зрительного тракта кпереди верхнего соединительного плеча приводит к развитию гемианоптического зрачка Вернике. Повреждение зрительного тракта при выходе из верхнего соединительного плеча сопровождается только гемианопсией.
Внутренняя капсула, или зрительная лучистость. При правостороннем поражении у больных обнаруживается гомонимная левосторонняя гемианопсия без гемианоптической неподвижности зрачков.
Ядро сфинктера. При повреждении ядра сфинктера справа развивается абсолютная неподвижность зрачка. Наряду с отсутствием прямой и непрямой реакций на свет, отмечается нарушение конвегренции справа. На левом глазу рефлексы сохранены.
Корковопокрышечный путь. Повреждение коры мозга и промежуточного мозга приводит к нарушению зрачкового рефлекса и аккомодации. При этом может развиться как возбуждение, так и торможение любой из этих функций.
Претектальная область. Специфическое нарушение зрачкового рефлекса при сифилисе описано Арджиллем—Робертсоном. При этом зрачок плохо реагирует на освещение глаза, но хорошо сохранена конвергенция и аккомодация. Зрачок обычно сужен и плохо расширяется при атропинизации. Наличие миоза связывают с разрушением кортикоспинальных волокон, которые ингибируют деятельность ядра Якубовича—Эдингера—Вестфаля. В результате этого повышается суживающий зрачок тонус. При этом синдроме обнаруживается и атрофия стромы радужной оболочки.
Синдром Арджилля—Робертсона также развивается при опухолевых (пинеалома), сифилитических и других поражениях претектальной области. Аккомодация сохранена по той причине, что надъядерные волокна, обслуживающие эту функцию, не представлены в претектальной области.
Чаще синдром Арджилля—Робертсона характеризуется нарушением зрачкового рефлекса на обоих глазах. В случаях одностороннего поражения полагают, что дефект волокон локализуется вне претектальной области, а именно в пучке волокон, уже направляющихся к ядру Якубовича—Эдингера—Вестфаля.
Средний мозг. Повреждения среднего мозга характеризуются расширением зрачка. Подобные нарушения могут возникнуть при повреждении ядра глазодвигательного нерва или нервных волокон нерва в месте выхода его на основании мозга. При этом зрачок плохо реагирует на свет. Выявляются различные дефекты подвижности глаза и птоз. Центростремительные дефекты зрачка возникают тогда, когда пупилломоторные тракты поражены на отрезке между зрительным трактом и ядром Якубовича—Эдингера—Вестфаля. При этом наибольшее практическое значение имеют три синдрома. Это синдром Арджилля—Робертсона, синдром Паринауда и «претектальные центростремительные дефекты зрачка».
Синдром Арджилля—Робертсона возникает у части больных тритичным сифилисом. У них зрачки значительно, но неодинаково, сужены и не реагируют на свет. Нередко развивается атрофия радужной оболочки. Необходимо отметить, что синдром Арджилля—Робертсона описан также при диабете, хроническом алкоголизме, энцефалите и некоторых других дегенеративных заболеваниях.
При синдроме Паринауда зрачки слабо реагируют на освещение. При этом обнаруживается также паралич взгляда кверху , нистагм, наличие приподнятого века. Наиболее частой причиной синдрома является возникновение опухоли шишковидной железы, рассеянный склероз и гидроцефалия.
Массивные патологические процессы (опухоли), приводящие к сдавлению среднего мозга, могут вызвать паралич мидриаза и конвергенции. Незначительные повреждения, вовлекающие глазодвигательные волокна в пределах среднего мозга, приводят к развитию эктопического зрачка (corectopia).
Ресничный ганглий. Воспалительные заболевания или травма ресничного ганглия иногда сопровождаются временным или постоянным параличом аккомодации, односторонним нарушением зрачкового рефлекса (тонический зрачок). Предполагают, что тонический зрачок, также известный как синдром Ади, складывается из патологической реакции зрачка, наиболее важным компонентом которого является миотоническое состояние аккомодации. Зрачок пораженной стороны реагирует на раздражитель медленнее, чем зрачок противоположной стороны, и расширяется более медленно. Пораженный зрачок обычно не реагирует на прямое и непрямое освещение. Сухожильный рефлекс отсутствует, но не обнаруживается нарушение двигательных функций и чувствительности.
Синдром чаще односторонний, но нередко становится двухсторонним. При этом зрачок плохо сужается при освещении. В щелевой лампе можно выявить сегментный паралич радужки и сегментный паралич ресничной мышцы, что связано с наличием асимметричной реиннервации.
Синдром Ади описан при опоясывающем герпетическом лишае, ветряной оспе, височном артериите, сифилисе, травме глазницы. Двухсторонние тонические зрачки описаны при диабете, хроническом алкоголизме, амилоидозе.
Повреждение глазодвигательного нерва. Развивается нарушение зрачкового рефлекса в результате повреждения эфферентной парасимпатичекой иннервации сфинктера. Сопровождаются эти изменения параличом глазодвигательных нервов. Поражение зрачковой реакции при параличе глазодвигательного нерва почти всегда сопровождается птозом и ограничением подвижности глаза. Расширение зрачка может быть единственным признаком паралича нерва при базальном менингите.
↑ Симпатическая система
Центральные симпатические пути. Симпатический тракт может быть поврежден на различных уровнях. При этом независимо от уровня поражения развивается синдром Горнера.
Характерными чертами этого синдрома являются:
- Миоз со стороны поражения, являющийся следствием паралича дилятатора.
- Нарушение расширения зрачка в темноте.
- Частичный птоз, развивающийся в результате паралича мюллеровской части леватора верхнего века.
- Ангидроз лица вследствие отсутствия стимуляции потовых желез.
- Энофтальм в результате пареза гладких мышц глазницы, включая мышцу Мюллера.
Деструктивные патологические процессы, разрушающие структуры моста, спинного мозга, вызывают односторонний синдром Горнера. В то же время процессы, «раздражающие» эти структуры, приводят к развитию синдрома Бернарда—Горнера (Bernard—Homer), который своими клиническими проявлениями противоположен предыдущему. При этом зрачок и глазные щели расширены, увеличено потоотделение, просвет сосудов на стороне повреждения сужен.
Врожденный синдром Горнера часто обнаруживается одновременно с гетерохромией радужной оболочки. При этом снижена пигментация радужки со стороны поражения. Гипопигментация связана с тем, что стромальные меланоциты содержат небольшое количество зерен меланина. Предполагают, что развитие гетерохромии связано с недостатком тирозиназы, наступающий в результате адренергической денервации. При этом депигментация не распространяется на невусные клетки радужки. Паралич Клумпа развивается после родовой травмы в результате повреждения двигательных корешков.
Центральные поражения симпатической системы наиболее часто связаны с окклюзией задней нижней мозжечковой артерии (синдром Валленберга). При этом выявляются:
- Синдром Горнера со стороны поражения.
- Дисфагия (паралич IX, X нервов).
- Аналгезия кожи лица со стороны поражения (спинной путь и ядра тройничного нерва) и аналгезия туловища и конечностей с противоположной стороны (восходящий спинно-таламический путь).
- Мозжечковая атаксия со стороны поражения и ротаторный нистагм (вестибулярные ядра). Центральные тракты, локализованные на уровне спинного мозга, наиболее часто повреждаются в результате травмы, развития опухолей, сирингомиелии и при демиелинизирующих заболеваниях.
Преганглионарные поражения. Поражение преганглионарных волокон (корешковые поражения) может развиться в результате родовой травмы и ассоциируется с параличом руки, развивающемся со стороны поражения (синдром Клюмпке). Опухоли верхушки легкого или верхней части средостения также приводят к повреждению преганглионарных волокон на уровне Т, (синдром Панкоаста; Pancoast). В области шеи волокна поражаются при развитии опухолевых, воспалительных, травматических заболеваний, увеличении лимфатических узлов. Описаны и случаи повреждения волокон при проведении хирургических вмешательств и каротидной ангиографии. Характеризуется синдром развитием сильных болей в плече, пареза и атрофии мышц руки в сочетании с легким энофтальмом, птозом и миозом. Нередко возникают эрозии в области третьего ребра.
Постганглионарные поражения. Постганглионарные поражения не сопровождаются ангидрозом кожи лица.
Внутренняя сонная артерия. Аневризмы основного ствола внутренней сонной артерии и ее ветвей вызывают расширение зрачка. При этом могут быть вовлечены оба компонента автономной нервной системы. Паралич симпатической системы приводит к миозу, увеличению слезоотделения, головным болям (синдром Нортона). Головная боль может сопровождаться птозом.
Парасимпатическая и симпатическая иннервация нередко нарушена и при воспалительных процессах в области верхней глазничной щели. Врожденный синдром Горнера развивается наиболее часто при родовой травме плечевого нервного сплетения. При этом нарушение иннервации радужки приводит к возникновению гетерохромии в результате нарушения пигментации пораженного глаза.
В заключение необходимо отметить, что выявить уровень поражения симпатической системы, приводящий к развитию синдрома Горнера, возможно на основании использования кокаина и гидроксиамфетамина. Приблизительно 90% норэпинефрина, выделяющегося в нервно-мышечных соединениях дилятатора радужки, заново поглощается пресинаптическими окончаниями. Кокаин блокирует механизм обратного поглощения норэпинефрина. Сохраняющаяся высокая концентрация норэпинефрина стимулирует мышечные клетки. В результате этого механизма, у пациента без поражения симпатического тракта зрачок расширится. В то же время у больного синдромом Горнера зрачок расширится незначительно, поскольку норэпинефрин вообще не высвобождается в пресинаптическую щель. Как указано выше, уточнить уровень поражения возможно при использовании гидроксиамфетамина (1% раствор). Гидроксиамфетамин способствует высвобождению в пресинаптическую щель норэпинефрина. По этой причине у здоровых пациентов этот препарат расширяет зрачок. При постганглионарном синдроме Горнера отмечается дегенерация нервных окончаний. По этой причине зрачок расширится незначительно. При преганглионарном синдроме Горнера постганглионарный нейрон сохраняется, в связи с чем при введении гидроксиамфетамина зрачок расширяется. Использование анализа реакции зрачка при проведении исследования с использованием кокаина и гидроксиамфетамина в достаточно большом числе случаев может уточнить уровень поражения, что имеет большое клиническое значение.
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0