Нуклеотидрегулируемые каналы сенсорных клеток (ЧАСТЬ І)


Описание
Основные свойства нуклеотидрегулируемых каналов
Изучению свойств ЦН-каналов посвящено множество публикаций. Систематически современные представления о свойствах этих каналов изложены в обзоре. Можно без преувеличения сказать, что этот тип каналов является одним из наиболее изученных. Структурно и филогенетически ЦН-каналы могут быть отнесены к большой группе потенциал-зависимых каналов, и многие их свойства могут оказаться сходными. Целью настоящего раздела является изложение накопившихся сегодня знаний о структуре и функциях ЦН-каналов.
Как уже было сказано выше, впервые прямая активация ионных каналов циклическими нуклеотидами была показана в плазматической мембране наружных сегментов фоторецепторных клеток сетчатки, где эти каналы участвуют в преобразовании света в электрический потенциал. С тех пор ЦН-каналы были найдены в разных видах клеток: обонятельных, ганглиозных и биполярных клетках сетчатки, слуховых волосковых клетках, вкусовых рецепторах, в печени, в шишковидной железе, сердце, центральной нервной системе и др.
ЦН-канал является гетеротетрамерным мембранным белком, состоящим из четырех субъединиц, по меньшей мере, двух типов, структурно гомологичных субъединицам потенциал-зависимых каналов. Он активируется в результате прямого связывания с молекулами циклических нуклеотидов. Было найдено, что большинство ЦН-каналов являются катионнеселективными, и их активность слабо зависит от величины мембранного потенциала. Они блокируются двухвалентными катионами, причем некоторые из них проникают сквозь сам канал.
Большинство исследований, посвященных работе ЦН-каналов, выполнены на палочках, колбочках и обонятельных нейронах. Структура и принципы работы ЦН-каналов в других типах клеток менее изучены, поэтому мы рассмотрим в основном только эти три типа клеток.
Установлено, что в наружном сегменте фоторецепторов ЦН- проводимость является единственным типом проводимости. В темноте ЦН-каналы фоторецепторных клеток активированы, и через них течет так называемый темновой ток. При освещении, в результате понижения концентрации цГМФ, ЦН-каналы закрываются, вызывая гиперполяризацию клеточной мембраны и понижение внутриклеточной концентрации Са2+. В обонятельных нейронах, напротив, ЦН-каналы закрыты в покое и активируются в результате стимуляции ароматическими молекулами, вызывая увеличение внутриклеточной концентрации Са2+, а электрическим сигналом в обонятельном нейроне является деполяризация клеточной мембраны.
Функциональные свойства ЦН-каналов. Все ЦН-каналы отличаются относительно высоким сродством к циклическим нуклеотидам, избирательной проводимостью положительных ионов, низкой единичной проводимостью в присутствии двухвалентных катионов. Однако в разных типах клеток эти свойства ЦН-каналов могут существенно различаться своей ионной селективностью, избирательным сродством и чувствительностью к циклическим нуклеотидам, характером взаимодействия с ионами Са2+. ЦН-каналы бывают катионнеселективными, как в фоторецепторных клетках, или селективными к К+, как в мышцах дрозофилы или в обонятельных нейронах лобстера. Некоторые каналы, например, ЦН-каналы в мышцах дрозофилы или ЦН-каналы волооковых клеток, более специфичны к цАМФ, чем к цГМФ. Величина чувствительности к циклическим нуклеотидам ЦН-каналов фоторецепторных клеток на порядок меньше, чем у каналов в обонятельных сенсорных нейронах.
Важнейшим функциональным свойством ЦН-каналов является осуществление и регуляция диффузии ионов Na+, Са2+, К сквозь липидный мембранный барьер. В наружном сегменте фоторецепторных клеток эти каналы осуществляют единственный тип проводимости и, следовательно, определяют величину мембранного потенциала фоторецептора. В результате активации ЦН-канала в ответ на увеличение концентрации цГМФ (в случае фоторецепторных клеток) или цАМФ (в случае обонятельных нейронов) происходят увеличение внутриклеточной концентрации Са2+ и деполяризация клеточной мембраны.
Важной характеристикой всех ЦН- каналов является их крайне низкая единичная проводимость в присутствии двухвалентных катионов. Так, если проводимость ЦН-каналов палочек в бескальциевой среде составляет 20-45 пС, то в присутствии двухвалентных катионов она уменьшается до 0,1-0,01 пС. Столь низкая проводимость имеет принципиальное значение для функционирования сенсорной клетки. Фоторецептор и обонятельная клетка могут работать в условиях так называемой предельной чувствительности. Важно понимать, что при очень низких значениях порога чувствительности критичным становится не высокий коэффициент усиления, а возможность эффективного выделения сигнала из шума. Ясно, что соотношение сигнал/шум для 1000 флуктуирующих каналов небольшой проводимости выше, чем для одного флуктуирующего канала с большой проводимостью.
Второй важной функциональном особенностью ЦН каналов является их способности регулировать внутриклеточную концентрацию ионов кальция. Фракция ионов Ca2+составляет в среднем от 12 до 21%. Поскольку внутриклеточная концентрация кальция относительно невелика, его регуляция может осуществляться ЦН-каналами даже в том случае, когда их плотность, как в фоторецепторе или цилии обонятельного нейрона, не так высока. Более того, есть основания полагать, что в этом случае регуляция внутриклеточного кальция становится основной функцией ЦН-каналов.
В фотооецепторных клетках концентрация внутриклеточного Са2+ определяется балансом постоянного входящего темново- го тока Са2+ черес открытый ЦН-канал и выходом ионов Са2+ через Na+ /Ca2+K+-обменник и равна -500 нМ. Когда ток ионов Са2+ внутрь клетки блокируется в результате световой инактивации ЦН-каналов, происходит понижение концентрации внутриклеточного Са2+ до ~50 нМ, что является началом фазы восстановления темнового потенциала. Снижение внутриклеточной концентрации кальция приводит к активации гуанилатциклазы и увеличению концентрации цГМФ в клетке Таким образом, проникновение ионов Ca2+ внутрь клетки через ЦН каналы вместе с регулированием остальных клеточных функций осуществляет обратную связь в механизме сигнальной транедукции.
Известно, что в сетчатке колбочки в 30-100 раз менее чувствительны к свету, чем палочки. Фотоответ в колбочках протекает быстрее, чем в палочках. Одной из причин, видимо, является разная скорость и величина изменений внутриклеточной концентрации Са2+, так как относительная проницаемость ионов Са2+различается в ЦН-каналах разных типов фоторецепторов. Так, в колбочках величина PСa/РВа равна 7,6 ± 0,8, а в палочках - 3,1 ± 1,0. Блокада ионами Са2+ имеет потенциал-зависимый характер - она максимальна в палочках и колбочках при равновесном потенциале и ослабевает при гипер- или деполяризации. Таким образом, более слабое взаимодействие с Са2+ в колбочках приводит к более быстрому току ионов Ca2+. Подобные результаты были получены при изучении свойств гомомерных каналов, образованных ?-субъединицами разных типов. Было обнаружено, что часть тока, переносимая ионами Са2+ (Рг), значительно больше в колбочках и обонятельных каналах, чем в палочковых. Так, фракция Са2+ от полного тока при концентрации Са2+ 0.3 мМ составляет 5,8% в колбочках, 3,9% - в обонятельных клетках и 0,25% - в палочках, а величина РСа/Ркравна 8,0, 4,6 и 1,7 соответственно.
В противоположность фоторецепторным клеткам ЦН-каналы обонятельных рецепторов закрыты в состоянии покоя и открываются в результате стимуляции ароматическими молекулами, что ведет к увеличению внутриклеточной концентрации Са2+. Повышение концентрации внутриклеточного Са2+ одновременно способствует и терминирует ответ обонятельных рецепторов. Са2+ также активирует деполяризующий ток ионов Cl-, таким образом существенно усиливая ответ. Повышение концентпацми Са2+стимулирует гидролиз цАМФ Са2+/кальмодулинактивируемой фосфидиэстеразой и запускает механизм инактивации ЦН-каналов, также включающий кальмодулин.
Преобладающая роль ЦН-каналов как регуляторов интегрального тока проявляется только в двух случаях: в наружных сегментах фоторецепторных и в цилиях обонятельных клеток, т.е. когда эти каналы являются важнейшими элементами механизма сенсорной трансдукции. Во всех других случаях можно считать, что основная функция этих каналов состоит в регуляции концентрации внутриклеточного кальция. Такова роль ЦН-каналов в нейронах сетчатки второго и третьего уровней: в биполярных и ганглиозных клетка, а также в других, нерецепторных тканях.
В последнее время появитесь данные, свидетельствующие о широком распространении ЦН-каналов в ЦНС. Например, Шиллс с соавторами обнаружили, что ЦН-каналы on-биполярных клеток сетчатки участвуют в регуляции мембранного ответа на входящий синаптический сигнал; при этом используется трансдукционный каскад, подобны» фоторецепторному. Найдены ЦН-каналы в ганглиозных клетках сетчатки крысы показали, что в нейронах гиппокампа экспрессируются ?-субъединицы обонятельного и палочкового LJH-каналов. С помощью ПЦР (полимеразная цепная реакция) показана экспрессия ЦН-каналов в разных отделах мозга. Интересно отметить, что в ЦНС ЦН-каналы являются новым, третьим по счету, типом каналов, осуществляющим Са2+-проводимость; первые два - это потенциал-зависимые Са2+-каналы и каналы, взаимодействующие с нейромедиаторами, такими как глутамат и ацетилхолин. Похоже, что в ЦНС, подобно сенсорным нейронам, в механизм регуляции работы ЦН-каналов включены: Са2+, каскад реакций, в котором участвуют G-белки, гуанилат- и аденилатциклазы, газовые месенджеры NO и СО. ЦН-каналы играют важную роль в нейронной пластичности, так как ток ионов Са2+ через каналы, расположенные в пресинаптической области, влияет на процесс высвобождения нейротрансмиттепа. В частности, важную роль в пресинаптической области играют цАМФ, цГМФ и NO, так как Са2+, проникающий внутрь через ЦН-каналы, влияет на процесс экзоцитоза, и этот эффект регулируется NO. Кроме того, ЦН-каналы экспрессированы на ранних стадиях развития нейронов еще до формирования синапса и участвуют в процессе узнавания клетки-мишени конусом роста аксона.
Изменение внутриклеточной концентрации кальция существенно не только для механизма трансдукции обонятельного и зрительного сигналов, но и для процессов экзоцитоза в колбочках, хемотаксиса сперматозоидов и т.д. Однако в большинстве нерецепторных тканей физиологическая роль ЦН каналов и особенности их работы остаются не вполне ясными.
Молекулярная структура нуклеотидрегулируемых каналов. ЦН-канал формирует гетеритетрамерный комплекс, состоящий из четырех разных, хотя и гомологичных субъединиц, разделяемых обычно на два типа - ? и ?, и структурно относится к семейству потенциал-зависимых каналов. Эти два типа субъединиц имеют общие структурные особенности: они состоят из основной структурной части, состоящей из шести трансмембранных сегментов, части, образующей ионную пору, домена, связывающего циклические нуклеотиды, и части, с которой связывается модулятор канала Са2+/кальмодулин (рис. 4.1).
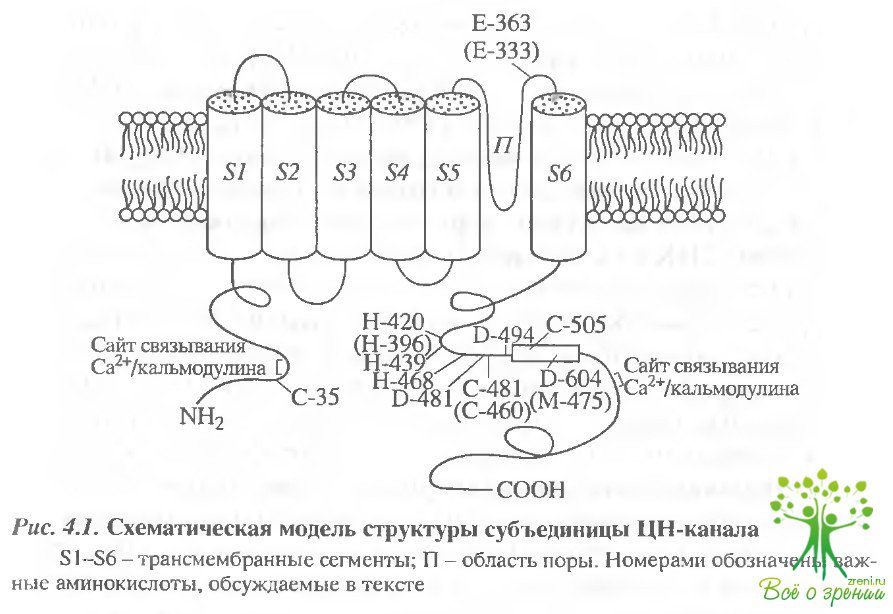
?-субъединица. В каждом из трех видов рецепторных клеток: палочках и колбочках сетчатки, а также в обонятельных сенсорных нейронах формируется уникальная форма ?- субъединицы канала (ЦНК?), называемая (в разных источниках по-разному) ЦНК?1 (пЦНК, ЦНК1) в палочках, ЦНК?З (ЦНКЗ, кЦНК) в колбочках и ЦНК?2 (ЦНК2, оЦНК) в обонятельных клетках. Она является основной субъединицей (молекулярная масса -63 кДа). которая формирует функциональные свойства и ключевые характеристики ЦН-канала, так как из ?-субъединиц, в отличие от ?-субъединиц, может быть сформирован функциональный гомомерный канал, свойства которого отличны от нативного.
Во всех типах клеток ?-субъгдиницы (ЦНК?1, ЦНК? и ЦНК?3) обладают общими структурными свойствами, которые также присущи субъединицам потенциал-зависимых каналов, и характеризуются наличием шести трансмембранных ос-спиралей (S1-S6) и шпилькоподобной области, формирующей ионопроницаемую пору. По своим функциональным свойствам ЦН-каналы относятся к лиганд-зависимым каналам.
Во всех тканях, где были найдены ЦН-каналы, обнаружены только эти три (ЦНК?1, ЦНК?2 и ЦНК?З) типа ?-субъединиц. Так, клон ДНК, кодирующий ЦНК?, был выделен из клеток печени, ганглиозных клеток сетчатки, биполярных клеток и клеток шишковидной железы; ДНК, кодирующая ЦНК?, была выделена из клеток печени, яичек, клеток шишковидной железы. В некоторых из этих тканей соответствующие ?-субъединицы были идентифицированы путем наращивания коротких последовательностей ДНК из колбочек. Таким образом, эти субъединицы могут представлять собой сплайс-вариант генов, экспрессивных в фоторецепторах позвоночных или обонятельных нейронах. Помимо сплайс-вариантов, функциональное разнообразие ЦН-каналов может быть расширено в результате использования различных субъединиц и с помощью посттрансляционного процес-синга полипептида, образующего канал.
Дополнительные (модуляторные) субъединицы. Дополнительные субъединицы, отличие от основных ?-субъединиц, не могут формировать функциональный канал сами по себе. Однако, будучи коэкспрессированы с ?-субъединицами, они образуют гетеро-олигомерный канал, свойства которого характерны для нативного ЦН-канала.
В настоящее время известны следующие дополнительные субъединицы: ЦНК?1а обнаружена в палочках сетчатки, ЦНК?4 и ЦНК найдены в обонятельных рецепторах Эти субъединицы гомологичны ?-субъединицам, они модулируют активность канала, сообщая ему специфические свойства, которые собственно и формируют природный ЦН- канал.
В фоторецепторных колбочках дополнительные субъединицы не найдены; в них экспрессирована только одна ЦНК?3 субъединица.
Хотя принципиальные структурные особенности обоих типов дополнительных субъединиц схожи с особенностями ?-субъединиц, включал шесть трансмембранных сегментов, ионопроницаемую пору и область связывания ЦН, они не экспрессируются как функциональные каналы. Однако, будучи коэксприссирована с ?-субъединицей, ?-субъединица фоторецепторной палочки, ЦНК(31а, образует гетероолигомерный канал, подобный природному, обладающий специфическими свойствами, такими как чувствительность к Са2+/кальмодуину или мерцательный характер работы канала (single- channel flickering). Эти индуцируемые ?-субъециницей свойства, также характерные для нативного ЦН-канала, могут служить доказательством того, что природный ЦН канал состоит из ?- и ?-субъединиц. Это наблюдение подтверждается при биохимическом выделении ЦН-кэнала фоторецепторных палочек, который состоит из ЦНК?1 субъединицы (молекулярный вес 63 кДа) и ЦНК?1а субъединицы (молекулярный вес 240 кДа). По своей основной последовательности эти субъединицы отличаются друг от друга намного более существенно, чем разные ?-субъединицы, что подтверждает их функциональную неэквивалентность.
В ЦН-канал обонятельного нейрона, в отличие от палочкового канала, включены две разные дополнительные субъединицы: ЦНК?4 и ЦНК?1b. Они структурно несколько отличаются от ?-субъединицы канала палочки. Так, вторая субъединица обонятельного ЦН-канала - ЦНК?4 имеет намного более короткий N-конец, чем субъединица фоторецепторной палочки ЦНК?1а и филогенетически намного ближе к ?-субъединицам, чем ЦНК?1а. Подобно ЦНХР1а из фоторецепторной палочки, собственно ЦНК?4 не образует функционального канала, но при коэкспрессии с ЦНК?З сообщает образовавшемуся гетерослигомерному каналу большую чувствительность к сАМФ и мерцательный (спонтанный) характер открывания канала. Однако чувствительность гетеромера ЦНК?3, ЦНК?4 более чем в два раза ниже чувствительности природного канала. В настоящее время существуют данные, свидетельствующие о том, что ЦН-канал из обонятельных нейронов состоит по меньшей мере из трех различных, хотя и гомологичных субъединиц. Третья субъединица обонятельного ЦН-канала, ЦНК4.3 или ЦНК?1b , представляет собой альтернативную сплайс-форму палочковой ЦНК?1а субъединицы. Хорошее соответствие таких свойств природного обонятельного ЦН-канала и канала, образованного в результате коэкспрессии трех разных субъединиц, как чувствительность к цАМФ, кинетика и проводимость одиночного ЦН-канала позволяют предположить, что природный ЦН-канал из обонятельного нейрона состоит именно из этих субъединиц - ЦНК?lb, ЦНК?3 и ЦНК?4.
Согласно последним данным, по своей структуре ЦН-канал фоторецепторных палочек формируется четырьмя субъединицами, причем одинаковые субъединицы расположены по диагонали напротив друг друга: ?-?-?-?. Возможно, четыре субъединицы, формирующие канал, могут быть ассоциированы и активируются как два независимых канальных димера. Для обонятельного ЦН-канала была предложена другая конформация: ?-?-?-?. Однако, учитывая последние данные о существовании в обонятельных клетках двух разных ?-субъединиц, следует предположить возможность другой конформации обонятельного канала.
Область связывания циклических нуклеотидов. Областью связывания циклических нуклеотидов называется домен, расположенный в С-концевой части каждой из субъединиц, формирующих ЦН-канал.
Области связывания ЦН обонятельных и палочковых ЦН-каналов отличаются высокой степенью подобия (более 80% аминокислот идентичны), хотя они и имеют разную селективность по отношению к циклическим нуклеотидам. Канал палочки наиболее эффективно активируется цГМФ, тогда как на обонятельный канал цАМФ и цГМФ влияют примерно одинаково.
Область связывания ЦН формируется последовательностью примерно 130 аминокислот из С концевой последовательности белка канала. Эта последовательность в основном гомологична САР-белку Е. с oil и аналогичной последовательности аминокислот протеинкиназы и структурно состоит из двух ?-спиралей (В и С) и восьми цепей в ?-конформации (?-roll) (?1 —?8), причем молекула ЦН связывается внутри (?-roll), а его пуриновое кольцо взаимодействует с Т-127 С спирали САР белка.
Varnum et al. [1995] исследовали молекулярный механизм способности ЦН-каналов различать цАМФ и цГМФ. Они обнаружили? что фоторецепторные и обонятельные ЦН-каналы по-разному активируются только лигандами, отличающимися по структуре их пуринового кольца. Было найдено, что селективность ЦН-канала (цГМФ цАМФ) палочки из куриной сетчатки существенно изменяется в результате нейтрализации остатка аспарагиновой кислоты (D-604) в домене, связывающем ЦН (цГМФ ? цАМФ). D-604 аналогичен позиции треонина Т-127 в САР-белке. Замена в этой позиции (D-604) на неполярный остаток инвертирует селективность канала к агонисту (цАМФ цГМФ). Этот эффект связан не с модификацией сродства канала с лигандом, а является резульатом изменения относительной способности агониста вызывать изменение аллостерической конформации, связанное с активацией канала Таким образом, этот аминокислотный остаток (D-604) играет ключевую роль в процессе избирательной активации ЦН- канала палочки в результате связывания с молекулой цГМФ
По всей видимости, специфичность канала к циклическим нуклеотидам (цАМФ и цГМФ) во многом определяется наличием в ?С-спирали отрицательно заряженных остатков D-604 и Y-468
Замена аспарагиновой кислоты D-604 на метионин вызывает существенное уменьшение эффективности цГМФ и увеличение эффективности цАМФ. Электрическое взаимодействие между карбоксильной группой и пуриновым кольцом цАМФ энергетически невыгодно, что объясняет столь редкую в норме активацию канала цАМФ. Протонирование в этом месте замещает это невыгодное взаимодействие и позволяет цАМФ действовать как почти полному агонисту.
Протонирование второй аминокислоты - гистидина Н-468 - способствует увеличению нуклеотид-неспецифичной селективности ЦН-канала и, похоже, является элементом механизма открывания канала. Этот эффект протонного взаимодействия проявляется для нативных каналов фоторецепторных палочек в меньшей степени, чем для каналов, состоящих из ?-субъединиц. В гетеромультимерных каналах, сформированных в результате коэкспрессии субъединиц аир, наблюдалось подобное уменьшение селективности ЦН- канала. Уменьшение влияния рН на нативные каналы может быть объяснено отсутствием поотонируемых аминокислот в ?-субъединице в позициях, соответствующих Н 468 и D-604.
Для обонятельных каналов в работе Shapiro, Zagotta [2000] показано, что замена единственного аминокислотного остатка ?-субъединицы, метионина в позиции 475 на глутамат (М475Е) вызывает изменение селективности канала к циклическим нуклеотидам: увеличивает сродство к цГМФ и уменьшает сродство к цАМФ. Метионин М-475 аналогичен аминокислотному остатку D-604 ?-субъединицы палочкового канала
Таким образом, можно считать, что аспарагиновая кислота D-604 из ?-субъединицы палочковых каналов и метионин М-475 из ?-субъединицы обонятельных каналов, расположенные в области связывания ЦН, являются определяющими для селективности канала к циклическим нуклеотидам.
Ц-cвязывающая область.Ц-связывающей областью называется цепочка, состоятцая примерно из 90 аминокислот, которая соединяет сайт связывания ЦН с последним трансмембранным участком канала.
Установлено, что Ц-связывающая область играет решающую роль в процессе активации канала, происходящем после связывания с циклическим нуклеотидом. Она влияет на сродство агониста и эффективность, с которой, циклические нуклеотиды открывают канал.
Gordon et al. [1996] обнаружили, что скоординированное связывание Ni2+между дзумя гистидиновыми остатками (Н-420) субъединиц канала фоторецепторной палочки, экспрессированного в ооцигах Xenopus, находящегося в открытом состоянии, вызывает увеличение сродства к цГМФ и как следствие увеличение цГМФ-зависимого тока этого канала. Связывание Ni2+ с аналогичными гистидиновыми остатками (Н-396) в обонятельных нейронах вызывает обратный эффект — ингибировачие. Таким образом, эта особая Ц-связывающая область, возможно, участвует в движении, происходящем при открывании канала.
Как выяснилось, различие в сродстве и чувствительности к циклическим нуклеотидам определяется не только областью связывания ЦН, но и Ц-связывающей областью. Так, в результате замены грех аминокислот в Ц-связывающей области канала колбочки на соответствующие аминокислоты из обонятельного канала (изолейцина 1439V, аспарагиновой кислоты D481А и D494S) существенно увеличилась эффективность цАМФ и повысилось сродство к цАМФ и цГМФ, а кинетика стала подобна кинетике ЦИ-канала из обонятельного нейрона, что объясняет повышение эффективности цАМФ.
Необходимо отметить также ключевую роль нескольких цистеиновых остатков, расположенных около или внутри сайта связывания ЦН. Существует мнение, что модификация этих остатков влияет на реакцию открывания канала. Это происходит либо в результате воздействия на каждую из субъединиц, либо в результате влияния на взаимодействие между cубъединицами. Так, модификация цистечна C-460 из Ц-области обонятельного ЦН-канала вызывает активацию канала в отсутствие циклических нуклеотидов.
Модификация SH-реагентами ?-субъединицы канала из се-чатки цыпленка приводила к значительному увеличению чувствительности канала и к цГМФ. и к цАМФ, и. следова гельно. к увеличению активации канала. Этот эффект отсутствовал при замене цистеинового остатка С-481. соответствующего цистеину С-460 из канала обонятельного нейрона, на неактивный (нейтральный) аланиновый остаток. Увеличение сродства к агонисту происходило быстрее при насыщающей концентрации цГМФ. что свидетельствует о большей доступности этой области, когда канал открыт.
Все эти результаты подтверждают, что Ц-связывающая область подвержена существенному сдвигу во время активации канала и играет ключевую роль в осуществлении опосредованной связи между двумя событиями — связыванием лиганда и открыванием поры.
Статья из книги: Молекулярные механизмы зрительной рецепции | Каламкаров Г.Р., Островский М.А..

Комментариев 0