Физиология цветового зрения

Содержание:
Описание

Первая физиологическая информация на клеточном уровне была получена спустя 250 лет после Ньютона в исследованиях шведско-финско-венесуэльского физиолога Гуннара Светихина, который в 1956 году на костистой рыбе осуществил внутриклеточную регистрацию активности нейтронов сетчатки — сначала он принял их за колбочки, но они оказались горизонтальными клетками.
На освещение сетчатки эти клетки отвечали только медленными потенциалами (потенциалов действия не наблюдалось). Как показано на рис. 125, Светихин обнаружил три типа клеток: первый тип, названный им L-клетками, гиперполяризовался при световой стимуляции независимо от спектрального состава света; второй тип, названной r-g-клетками, гиперполяризовался волнами малой длины с максимумом ответа на зеленый свет и деполяризовался волнами большой длины с максимумом ответа на красный свет; третий тип, названный с учетом теории Геринга у-клетками, отвечал по типу клеток r-g, но с максимумом гиперполяризации на синий и максимумом деполяризации на желтый свет.
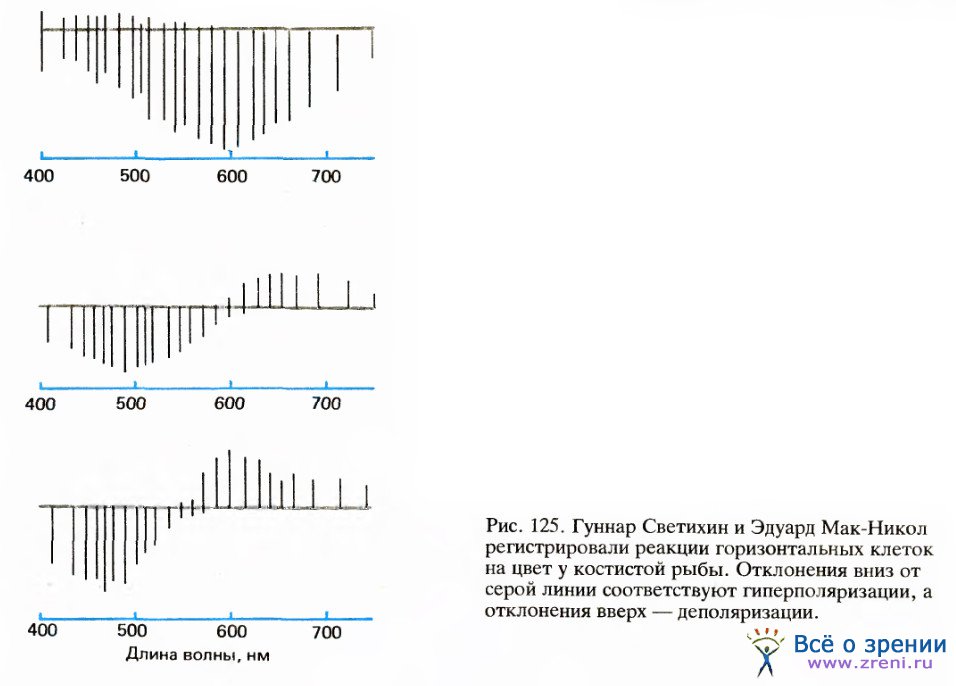
У клеток r-g и у-b белый свет вызывал лишь слабые и быстро затухающие ответы, как и следовало ожидать ввиду широкополосного спектрально-энергетического состава белого света. Кроме того, у клеток обоих этих типов, которые мы можем назвать оппонентно-цветовыми клетками, свет с некоторой промежуточной длиной волны, называющейся тонкой перекреста, не вызывал никакой реакции. Поскольку эти клетки реагируют на окрашенный, но не на белый свет, они, вероятно, связаны с цветовыми ощущениями.
В 1958 году Рассел де Валуа и его сотрудники зарегистрировали ответы, поразительно сходные со светихинскими, от клеток наружного коленчатого тела макака. Ранее с помощью поведенческих тестов де Валуа показал, что цветовое зрение макака и человека почти одинаково; например, соотношение, в котором нужно смешать два цветных луча, чтобы получить третий цвет, почти идентично у обоих видов. Поэтому можно думать, что макаки и люди обладают сходными механизмами на нижних уровнях зрительной системы, и мы, по-видимому, вправе сравнивать психофизику цвета у человека с физиологией у макака. Де Валуа обнаружил, что многие клетки коленчатого тела активировались рассеянным монохроматическим светом в диапазоне от одного конца спектра до точки перекреста, где реакция отсутствовала, и подавлялись светом во втором диапазоне — от точки перекреста до другого конца спектра.
И вновь аналогия с цветовыми процессами Геринга была полной: де Валуа выявил оппонентно-цветовые клетки двух типов, красно-зеленые и желтосиние; у каждого типа смешение двух световых волн, длины которых на волновой шкале располагались симметрично относительно точки перекреста, приводило к взаимному погашению реакций точно так же, как в восприятии добавление синего к желтому или зеленого к красному порождает белый. Результаты де Валуа особенно хорошо согласовались с формулировками Геринга, поскольку две группы цветовых клеток имели максимумы реакции и точки перекреста точно в тех местах спектральной шкалы, чтобы одна группа могла отражать «желто-синие» свойства падающего света, а другая группа — «красно-зеленые».

Следующий шаг состоял в том, чтобы рассмотреть рецептивные поля этих клеток, используя вместо рассеянного света маленькие цветные пятнышки; это мы и сделали в 1966 году совместно с Торстеном Визелом. У рецептивных полей большинства оппонентно-цветовых клеток де Валуа обнаружилась удивительная организация, которая до сих пор ставит нас в тупик. Клетки, как и у кошек по данным Куффлера, имели поля, разделенные на два антагонистических участка — центр и периферию; центр мог быть типа on или off. В типичном случае центр поля представлен исключительно красными, а тормозящая периферия — исключительно зелеными колбочками.
Поэтому при красном свете как маленькое, так и большое пятно вызывает энергичную реакцию, поскольку центр избирательно чувствителен к длинноволновому свету, а периферия на него почти не реагирует; при коротковолновом свете маленькие пятна дают лишь очень слабую реакцию или не вызывают ее вовсе, а большие порождают сильное торможение с off-реакциями. При белом свете, содержащем короткие и длинные волны, малые пятна вызывают on-реакции, а большие не вызывают ответа.
Хотя нашим первым впечатлением было, что такая клетка из центра должна получать входные сигналы от красных, а с периферии — от зеленых колбочек, теперь кажется вероятным, что полное рецептивное поле включает две перекрывающиеся системы, как это показано на рис. 127.
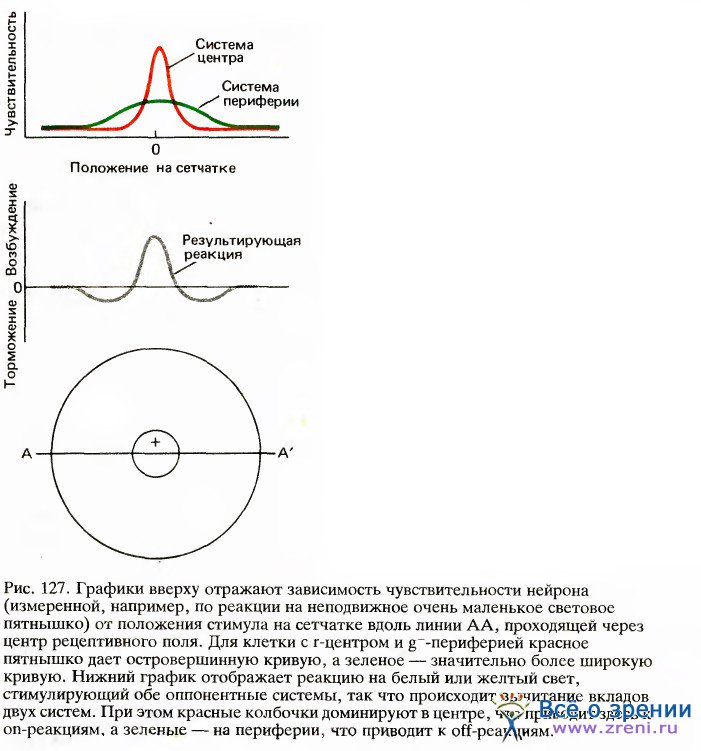
Как красные, так и зеленые колбочки распределены по довольно широкой круговой области, причем их количество максимально в центре и убывает по мере удаления от него. В центре красные колбочки сильно преобладают, но к периферии число их падает значительно быстрее числа зеленых колбочек. Поэтому небольшое пятнышко, вспыхивающее в центре и содержащее длинноволновую компоненту, будет весьма мощным стимулом для красной системы; даже если оно стимулирует и зеленые колбочки, их число по сравнению с общим числом связанных с клеткой зеленых колбочек будет слишком мало, чтобы составить какую-либо конкуренцию красной системе.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Те же соображения приложимы и к клеткам с центром и периферией, описанным в главе 3, рецептивные поля которых тоже должны состоять из двух оппонентных круговых перекрывающихся зон с разными формами кривых зависимости чувствительности от координаты. Таким образом, периферия, вероятно, имеет не кольцеобразную, как думали вначале, а круговую форму. В отношении этих оппонентно-цветовых клеток обезьян предполагается (правда, без достаточных данных), что периферические области отражают вклад горизонтальных клеток.
Реакция на диффузный свет — в данном случае on-реакции на красный, off-реакции на синий и зеленый и отсутствие реакции на белый свет — ясно показывают, что такая клетка должна регистрировать информацию о цвете. Но реакции на определенного рода белые края и отсутствие реакций на диффузный свет убеждают в том, что эта клетка связана также и с восприятием формы. Мы называем эти цветооппонентные клетки с центром и периферией клетками «типа 1».
Наружное коленчатое тело обезьяны, содержит шесть слоев, причем четыре верхних слоя содержат множество мелких клеток, а два нижних — меньшее число крупных клеток. Мы находим клетки описанного выше типа 1 в верхних, или мелко клеточных, слоях. Они различаются по типу колбочек, входящих в их центральную и периферийную системы, и по характеру центра, который может быть возбуждающим или тормозящим. Пример, представленный на рис. 126, мы можем обозначить как r+g“ (с красным центром и зеленой периферией). Среди подтипов клеток, получающих входы от красных и зеленых колбочек, мы находим все четыре возможных варианта: r+ g-, r“g+, g+r-, g~r+. Вторая группа клеток получает входы от синих колбочек из центра поля и от комбинации красных и зеленых (или, быть может, только от зеленых) — с периферии. Мы называем такие клетки «сине-желтыми», причем слово «желтые здесь употреблено для краткости вместо «красные плюс зеленые».
В четырех верхних слоях мы находим клетки еще двух типов. Клетки типа 2 составляют около 10 процентов нейронной популяции и имеют рецептивные поля, состоящие только из центра. Повсюду в этом центре у некоторых клеток выявляется красно-зеленая, а у остальных — сине-желтая оппонентность. Рецептивные поля еще примерно у 15 процентов клеток в четырех верхних слоях коленчатого тела и у всех клеток в двух нижних (крупноклеточных) слоях имеют центр и периферию, но эти клетки не проявляют цветовых предпочтений; создается впечатление, что центр и периферия их полей получают одинаковый относительный вклад от всех трех видов колбочек. Мы считаем эти клетки широкополосными и в верхних слоях называем их клетками типа 3.
Все эти данные поразительно согласуются с моделью Геринга: кроме двух классов клеток с цветовой оппонентностью мы имеем также третий класс, вовсе не обладающий этим свойством, но с широкополосной пространственной оппонентностью. Что как будто не согласуется ни с какой теорией, так это пространственная организация полей у оппонентно-цветовых клеток — клеток типа 1. На первый взгляд можно подумать, что эта организация каким-то образом связана с цветовым контрастом, т. е. с тенденцией одного цвета, например синего, казаться более ярким в окружении другого цвета, например зеленого, или с тем, что кусок серой бумаги кажется желтоватым, если мы видим его на синем фоне. Но минутное размышление убедит вас, что клетки коленчатого тела едва ли могут быть полезны для этого вида цветового контраста: описанная выше клетка с г+-центром и ^-периферией не будет сильно возбуждена красным пятном на зеленом фоне — она ответит очень слабо или совсем не ответит, так как один эффект уничтожится другим; произойдет обратное тому, что нужно для усиления цветового контраста.
О клетках типа 1 можно лишь утверждать, что по своей численности они должны быть наиболее важным для мозга источником цветовой информации, хотя эта информация представлена каким-то странным образом. Их возможная работа совместно с клетками типа 3 очень хорошо согласуется с представлениями Геринга о двух оппонентных цветовых системах и одной оппонентной пространственной системы. Вероятно, они играют также важную роль в восприятии тонких деталей формы объектов, поскольку единственные другие клетки коленчатого тела с маленькими центрами полей — это широкополосные клетки типа 3, которых в десять раз меньше.
Как мы видели раньше, остроту зрения, т. е. способность различать малые объекты, можно измерять несколькими способами; можно, например, определять наименьшее расстояние между двумя точками, при котором они еще не сливаются, или наименьший видимый разрыв в окружности (так называемом кольце Ландольта). Острота, измеренная по любому из этих способов, оказывается для центральной ямки порядка 0,5 угловой минуты, или около 1 миллиметра с расстояния 8 метров. Этот показатель хорошо согласуется с расстоянием между двумя колбочками в центральной ямке. Клетки коленчатого тела типа 1, получающие входные сигналы от зоны центральной ямки, имеют рецептивные поля с центрами порядка 2 угловых минут в диаметре. Кажется вероятным, что центр поля составляет здесь всего одна колбочка. Таким образом, мы находим разумное соответствие между остротой зрения и наименьшими размерами центра рецептивного поля у клеток наружного коленчатого тела.
Вентральная (нижняя) пара слоев коленчатого тела отличается от четырех дорсальных слоев тем, что состоит исключительно из клеток с широкополосными центрами полей. Эти клетки проявляют любопытную форму цветовой оппонентности, смысл которой никто не понимает и о которой я не будут говорить подробнее. Большинство считает эти клетки «слепыми» к цвету. Центры их полей в несколько раз крупнее, чем центры нейронов мелкоклеточных слоев, и у них есть ряд других интересных особенностей. В настоящее время мы подозреваем, что эти клетки обслуживают отделы мозга, играющие важную роль в восприятии формы, глубины и движения. Развитие этой темы увело бы нас далеко в сторону от цвета, и потребовалось бы написать еще одну книгу.
Большинство типов клеток, описанных для наружного коленчатого тела, встречается также и в сетчатке. В коленчатом теле они больше обособлены, что облегчает их изучение. Вклад коленчатого тела в анализ зрительной информации у обезьян до сих пор не известен.
↑ Нейронная основа константности цвета
Поскольку клетки типа 1 наружного коленчатого тела как будто не приспособлены для цвето-пространственных сравнений, нам, по-видимому, следует выйти на более высокие уровни зрительной системы. Для проверки идеи, согласно которой такие сравнения могли бы происходить в коре головного мозга, группа Лэнда, Маргарет Ливингстон и я изучали человека, у которого была произведена перерезка мозолистого тела с целью лечения эпилепсии. Между областями, разделенными средней вертикалью поля зрения, пространственно цветовые взаимодействия у него отсутствовали: на цвет пятна, расположенного чуть левее точки, на которую смотрел этот испытуемый, не влияли даже резкие цветовые изменения в правом зрительном поле, тогда как нормальные испытуемые сообщали в таких случаях о заметных изменениях. Это означает, что цвето-пространственные взаимодействия, по-видимому, не могут происходить в самой сетчатке. Хотя никто всерьез и не утверждал обратного, вопрос продолжал дискутироваться, и было приятно получить какие-то экспериментальные данные. Результаты этого эксперимента согласуются с тем фактом, что нам не удалось обнаружить в сетчатке такие ганглиозные клетки, которые вполне могли бы участвовать в цвето-пространственных взаимодействиях.
У золотой рыбки, осуществляющей пространственные сравнения не хуже нас с вами, коры головного мозга практически нет. Возможно, рыбка в отличие от нас выполняет эти сравнения с помощью своей сетчатки. Открытие в сетчатке рыб двойных оппонентных клеток (Н. Доу, 1968) как будто подтверждает это. У обезьян (см. следующую статью) мы находим такие клетки в коре, но их нет ни в сетчатке, ни в наружном коленчатом теле.
↑ Пузырьки
Примерно к 1978 году первичная зрительная кора обезьян с ее простыми и сложными клетками, с нейронами, реагирующими на концы линий, с глазодоминантными и ориентационными колонками представлялась достаточно изученной. Но неожиданной особенностью ее физиологии казалось то, что как будто лишь немногие клетки здесь небезразличны к цвету. Когда мы картировали рецептивное поле простой или сложной клетки, используя белый свет, а затем повторяли опыт с окрашенными пятнами или полосками, то результаты, как правило, были одинаковыми. Однако у некоторых клеток, составляющих, наверное, лишь десятую долю всех нейронов верхнего слоя коры, обнаруживались явные цветовые предпочтения — энергичные ответы на ориентированную полоску, скажем, красного цвета при полном отсутствии ответов на другие цвета или даже на белый цвет. Ориентационная избирательность этих клеток была не ниже, чем у клеток, нечувствительных к цвету, но большинство нейронов к цвету были безразличны. Все это казалось особенно странным ввиду того, что в наружном коленчатом теле очень большая доля клеток кодирует цвет, а коленчатое тело служит главным источником информации для зрительной коры. Трудно было понять, что могло происходить с этой цветовой информацией в коре.
Неожиданно в 1978 году все изменилось. Нейроанатом из Сиэтла Маргарет Уонг-Райли обнаружила, что при окрашивании коры на фермент цитохромок-сидазу в верхних слоях выявляется никогда не наблюдавшаяся ранее неоднородность — периодические темноокрашенные участки шириной около четверти миллиметра, разделенные промежутками около полумиллиметра. Цитохромоксидазу — участвующий в метаболизме фермент — содержат все клетки, и никто не мог даже подумать, что гистохимическая реакция на этот фермент позволит увидеть в коре что-либо интересное. Когда Уонг-Райли прислала нам свои микрофотографии, Торстен Визел и я заподозрили, что мы видим колонки глазодоминантности в поперечном разрезе и что большинство монокулярных клеток по каким-то причинам метаболически более активны, чем бинокулярные. Мы положили снимки в ящик стола и постарались забыть о них.
Прошло несколько лет, прежде чем нам или кому-либо другому довелось изучать с помощью той же реакции срезы первичной зрительной коры, параллельные ее поверхности. Когда это приблизительно одновременно было, наконец, сделано двумя группами (Анитой Хендриксон и А. Хамфри в Сиэтле и Дж. Хортоном и мной в Бостоне), выявился — ко всеобщему полному изумлению — узор, напоминающий материал в горошек. Пример показан на рис. 129.


Вместо полосок мы увидели похожие на пузырьки образования, которые ни с чем из известного ранее не связывались. Неоднородности Уонг-Райли называли всеми мыслимыми именами: точки, пуфы, мушки, пятнышки. Мы назвали их «пузырьками» [blobs — это слово наглядно, узаконено (оно есть даже в оксфордском словаре английского языка) и как будто раздражает наших конкурентов].
Следующая задача была очевидной: мы должны были снова регистрировать ответы клеток в стриарной коре, гистологически контролируя эксперимент окраской на цитохромоксидазу, и попытаться выявить нечто особенное у клеток, находящихся в пузырьках. В 1981 году Маргарет Ливингстон и я принялись за эту работу. Результат оказался совершенно неожиданным. Проходя расстояние в четверть миллиметра, равное диаметру пузырька, можно исследовать примерно пять или шесть клеток. Всякий раз, когда мы пересекали пузырек, попадавшиеся на пути электрода клетки были полностью лишены ориентационной избирательности, что заметно контрастировало с высокой ориентационной избирательностью клеток, расположенных вне пузырьков.
Этому отсутствию ориентационной специфичности можно было бы дать два объяснения. Во-первых, эти клетки могли неизбирательно получать входные сигналы от соседних ориентационных клеток, лежащих вне пузырьков, и поэтому еще способны были отвечать специфической реакцией на линии (полоски и т. п.), но после объединения всех возможных ориентаций всякое предпочтение какой-либо из них исчезало. Во-вторых, они могли быть похожими на клетки коленчатого тела или на корковые клетки слоя 4С и, таким образом, быть проще внепузырьковых ориентационно-избирательных клеток. Вскоре вопрос был разрешен: оказалось, что большинство этих клеток имеет рецептивные поля с центром и периферией. Несколько дополнительных экспериментов убедили нас в том, что многие из них участвуют в кодировании цвета.
Больше половины пузырьковых клеток имели оппонентно-цветовые рецептивные поля с центром и периферией, но вели себя явно более сложным образом, чем клетки типа 1 наружного коленчатого тела. Они практически не отвечали на белые пятна любых размеров и формы. Но на небольшие цветные пятна, вспыхивающие в центре рецептивного поля, они энергично реагировали в одном диапазоне длин волны и затормаживались в другом диапазоне; некоторые активировались длинными волнами (красным светом) и подавлялись короткими (зеленым и синим светом), другие вели себя противоположным образом. Как и среди клеток коленчатого тела, мы могли, в зависимости от положения максимальных реакций на спектральной шкале, выделить два класса — красно-зеленые и сине-желтые клетки. (Здесь, как и ранее, слова «красный», «зеленый» и «синий» указывают на соответствующие типы колбочек, а слово «желтый» — на параллельные входы от красных и зеленых колбочек.)
Таким образом, эти клетки весьма напоминали оппонентно-цветовые клетки коленчатого тела, обладающие только центром (тип 2). Но в отличие от клеток типа 2 эти цветокодирующие пузырьковые клетки чаще всего не отвечали на большие белые или цветные пятна, каков бы ни был спектральный состав света. Они вели себя так, как если бы центральная система рецептивного поля была у них окружена кольцом оппонентности. Если говорить о наиболее распространенном типе клеток, то центр типа r+g~, казалось, был окружен у них кольцом типа r-g+.
Мы с Маргарет Ливингстон назвали эти клетки двойными оппонентными из-за их красно-зеленой или желто-синей оппонентности в центре и антагонизма периферии по отношению к любой реакции в центре, будь она типа on или off. Поэтому они не отвечают не только на белый свет в любой геометрической конфигурации, но и на большие пятна, независимо от их спектрального состава. Центры рецептивных полей, как и у клеток типа 2, были в несколько раз больше, чем у клеток коленчатого тела типа 1. Как уже упоминалось, Н. Доу ввел термин двойные оппонентные для клеток, обнаруженных им в сетчатке золотой рыбки. Он предполагал, что сходные клетки могли бы участвовать в цвето-пространственных взаимодействиях у человека, и несколько лет спустя совместно с А. Пирлменом он старательно, хотя и безуспешно, искал такие клетки в наружном коленчатом теле макака.
В конце 60-х годов и позже двойные оппонентные клетки изредка находили в коре обезьян, но их не связывали четко с какими-либо анатомическими структурами. Нам до сих пор непонятны некоторые из их особенностей. Например, у только что описанных клеток типа r+g_ красное пятно, окруженное зеленым, часто вызывает слабую реакцию вместо энергичной, которой можно было бы ожидать.
Вперемешку с двойными оппонентными клетками обоих классов (краснозелеными и желто-синими) встречались и обычные широкополосные клетки с центром и периферией. Эти широкополосные клетки отличались от клеток верхних слоев коленчатого тела и от клеток слоя 4СР коры более крупными размерами своих центров.
Мы с Маргарет Ливингстон предположили, что пузырьки представляют собой ветвь зрительного пути, имеющую дело с «цветом» в широком смысле, включая оттенки черного, белого и серого. Эта система, видимо, обособляется от остальной части зрительного пути либо в наружном коленчатом теле, либо в слое 4 стриарной коры. Коленчатое тело, возможно, имеет прямую, хотя и слабую проекцию на пузырьки. Кажется вероятной и проекция на них слоя 4С(3 — может быть, для них это даже главный источник входных сигналов. Проецируется ли на них слой 4Са , неясно. Обычно для реакции пузырьковых клеток необходим граничный контраст — нужны либо границы яркости, на которые отвечают широкополосные клетки с центром и периферией, либо цветоконтрастные границы, на которые реагируют двойные оппонентные клетки. Как уже говорилось, это равнозначно тому, что такие клетки играют роль в создании константности цвета.
Если пузырьковые клетки участвуют в константности цвета, они не могут осуществлять в точности те вычисления, которые рассматривались Лэндом, а именно сравнения между каким-то участком и его окружением для каждого из колбочковых спектральных поддиапазонов. Вместо этого они, по-видимому, осуществляют сравнение по типу Геринга: сопоставляют красно-зеленое в одном участке с красно-зеленым в окружающем фоне и делают то же самое для желто-синего и для интенсивности света. Но оба способа трактовки цвета — г, g и b в одном случае и b-w, r-g, и y-b в другом — по существу, эквивалентны. Цвет требует задания трех переменных: любому цвету соответствует тройка чисел, и мы можем представлять любой цвет точкой в трехмерном пространстве. Точки в таком пространстве можно представлять не единственным способом. Система координат может быть декартовой с осями, расположенными по нашему усмотрению, либо мы можем использовать полярные или цилиндрические координаты. В теории Геринга (в сетчатке и мозгу, наверное, тоже) просто используется иная совокупность осей для описания того же пространства.

Эта теория, несомненно, упрощает действительность, так как пузырьковые клетки, принадлежащие к трем классам, вовсе не так сходны между собой, как горошины в стручке, — они значительно варьируют по относительной силе периферии и центра, по совершенству баланса между оппонентными цветами и по другим характеристикам, отчасти еще не вполне понятым. Сейчас мы можем лишь сказать, что физиология удивительно хорошо согласуется с психофизикой.
Может возникнуть вопрос: зачем мозгу понадобились для отображения цвета такие на первый взгляд странные оси вместо более простой системы осей г, g и Ь, используемой в рецепторном слое сетчатки? По-видимому, цветовое зрение добавилось в ходе эволюции к «бесцветному» восприятию низших млекопитающих. Цветовое пространство у этих животных было одномерным, а колбочки разных типов (если животное обладало не одним типом колбочек) были объединены в общий пул. В ходе эволюции цветового зрения к уже существующей оси добавились еще две. Это было разумнее, чем отбросить уже сложившуюся черно-белую систему, а потом создавать три новые системы. Когда мы адаптируемся к темноте и используем только палочки, наше зрение лишается цвета и вновь отображается на единственную ось, в создании которой, очевидно, участвуют палочки. Это было бы не так просто сделать с осями г, g и Ь.
В настоящее время мы можем лишь гадать, какова схема связей двойных оппонентных клеток. На рис. 131 показан один из возможных вариантов их соединения с клетками верхних слоев коленчатого тела или с клетками слоя 4Сb коры.

Излишне говорить, что все это чисто умозрительно; существенно лишь то, что двойная оппонентность каким-то образом должна достигаться: либо ее источник находится на нижних уровнях (сетчатки или коленчатое тело), и в этом случае мы просто еще не обнаружили клеток этого типа, либо это должно происходить в коре. Наличие таких клеток в сетчатке рыб не доказывает их существования у млекопитающих. Схема на рис. 131 всего лишь показывает, как могли бы быть сформированы центры рецептивных полей: для периферии мы всегда можем представить себе обратную процедуру с использованием клеток г —g+ коленчатого тела, поля которых широко перекрываются.
Наше стремление рассматривать цвет и форму как раздельные аспекты восприятия находит, таким образом, подтверждение в виде физического обособления пузырьковых и непузырьковых участков первичной зрительной коры. В областях выше стриарной коры — в зрительной зоне 2 и даже еще выше — эта обособленность сохраняется. Мы не знаем, где происходит их объединение, если оно вообще где-нибудь происходит.
↑ Заключение
Самое интересное в цветовом зрении то, что здесь удается, объединив психофизические и нейрофизиологические методы, понять такие явления, как результаты смешения цветов или константность цвета, которые могли бы представляться совершенно загадочными. Проблемы, связанные с цветом, при всей их сложности, вероятно, проще тех, которые связаны с формой. Несмотря на открытие ориентационно-специфических клеток или клеток, реагирующих на концы линий, мы еще далеки от понимания нашей способности распознавать формы, отличать формы от их фона или воссоздавать трехмерную картину по плоским изображениям на сетчатке каждого глаза.
Сопоставление этих двух модальностей — цвета и формы — вообще может ввести в заблуждение: вспомним, что одни лишь различия цвета на границах в отсутствие каких-либо различий освещенности могут приводить к восприятию формы. Цвет в этом смысле ничем не отличается от черного и белого и служит лишь одним из механизмов проявления формы.
---
Статья из книги: Глаз, мозг, зрение | Хьюбел Д.

Комментариев 0