Экологические вариации сетчатки и зрительных центров


Описание
Общеизвестно, что существует общий план строения сетчатки у низших позвоночных и млекопитающих, включая приматов и человека. В данной главе будет сделана попытка выделить особенности, свойственные некоторым отдельным видам и рассмотреть их как в экологическом, так и эволюционном аспекте.Сетчатка отражает те особенности, которые выражены в строении глаза в целом, его форме, величине, строении аккомодационного аппарата. В труде Walls [1942] соотношение экологии с глазом и сетчаткой у позвоночных превосходно обобщено. По Е. А. Бабуриной, строение сетчатки отражает видовую экологическую специфику. В работах Е. А. Бабуриной, представленных в монографии [1972] показано также, что строение глаза и сетчатки специфично для каждого этапа развития также в связи с экологией. Например, иредличинки сельдей обладают колбочковой сетчаткой. В мальковом периоде сетчатка обогащается палочками в связи с возникновением сумеречного зрения. У взрослых сельдей полное развитие получают как колбочковый, так и палочковый аппараты. У некоторых видов рыб молодые особи обладают областью острого зрения, полностью редуцирующейся у взрослых особей [Бабурина, 1961].
Как яркий пример различий между двумя экологическими группами Е. А. Бабурина [1964] рассматривает формирование сетчатки у карповых рыб. Глаза взрослых карповых рыб хорошо развиты, в сетчатке имеются и колбочки, и палочки; зрительная ориентация имеет решающее значение для активного добывания пищи. При этих общих чертах формирования глаз существенные различия отмечаются в особенностях развития сетчатки у литофильных и пелагофильных карповых рыб.
У литофильных (икра которых приклеивается к камням) карповых (рыбца и шемаи) рано появляется способность к отрицательной фотокинетической реакции при уже образовавшемся зрительном нерве и зрелом состоянии пигментного эпителия. Палочки и колбочки, одпако, еще не развиты. К началу личиночного периода у литофильных рыб в сетчатке формируются фоторецепторы, у личинок имеются палочки и колбочки, сетчатка способна к ретиномоторной реакции, а глаз созревает для восприятия оформленного раздражителя.
Совершенно иначе происходит развитие сетчатки у пелагофильных карповых рыб (откладывающих икру, плывущую по течению в воде), например, востробрюшки и чехонки. Пигментация клеток пигментного эпителия завершается значительно позже, когда в сетчатке уже сформированы все слои и образованы фоторецепторы — колбочки, но сетчатка еще не способна к ретиномоторной реакции. Лишь в конце предличиночного периода у предличинок возникает положительная фотокинетическая реакция. У взрослых особей в сетчатке, кроме колбочек, имеется много палочек и развивается способность к выраженной ретиномоторной реакции [Бабурина, 1972].
Далее в трудах Е. А. Бабуриной приведены примеры сходного развития сетчатки у рыб разных семейств и отрядов, но со сходной экологией в сравнимых периодах жизни. В частности,, обнаруживается сходство у судака и сельди-черноспинки с карповыми пелагофильными рыбами в сроках формирования пигментного эпителия, отстающего от развития других слоев сетчатки, и во времени появления ретиномоторной реакции.
Яркие отличия фоторецепторов у различных позвоночных демонстрировал Kolmer [1936]. Наличие или отсутствие колбочек двойные колбочки различной конфигурации, жировые капли хорошо видны на многочисленных рисунках в его трудах. Современные ультрамикроскопические исследования позволили проанализировать строение эллипсоида, параболоида, миоида, дисков рецепторов и показать связь структурных параметров с функциональными особенностями [Винников, 1971, 1979; Лычаков, 1977; Говардовский, 1978, 1983; Островский, 1971; Бочкин,. Островский, 1984; Бызов, 1984; и др.].
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Тщательно изучается пространственное расположение фоторецепторов сетчатки (рис. 103, 104),
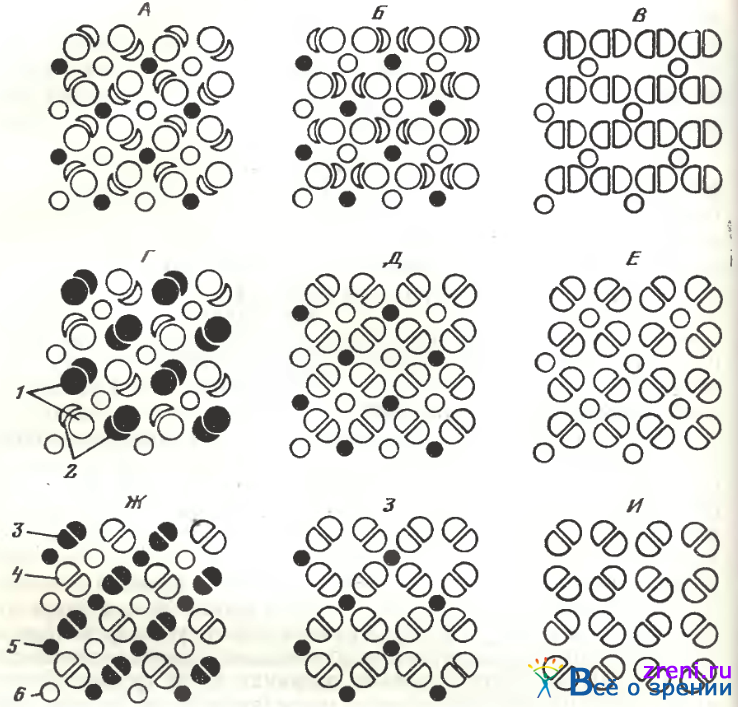
Рис. 103. Схемы различных модификаций рисунка мозаики колбочек в сетчатке костистых рыб [Подугольникова, Максимов, 1977]
1 — главные элементы неравночленных двойных колбочек разных типов; 2 — добавочные элементы; 3, 4 — равночленные двойные колбочки разных типов; 5 — дополнительная (короткая) 6 — центральная (длинная) единичные колбочки; А — И —различные модификации мозаик, характерные для представителей различных видов рыб (карповых) (Б). кефалеобразных (Д). окунеобразных (Е) и др.
1 — главные элементы неравночленных двойных колбочек разных типов; 2 — добавочные элементы; 3, 4 — равночленные двойные колбочки разных типов; 5 — дополнительная (короткая) 6 — центральная (длинная) единичные колбочки; А — И —различные модификации мозаик, характерные для представителей различных видов рыб (карповых) (Б). кефалеобразных (Д). окунеобразных (Е) и др.
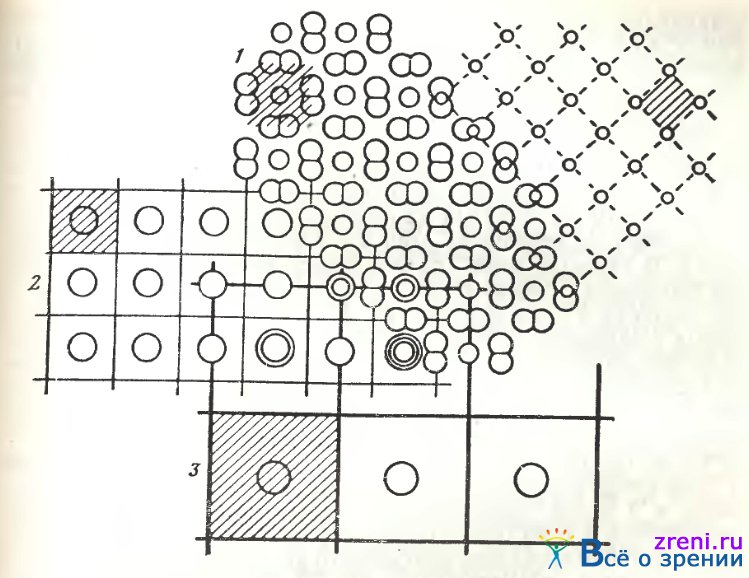
Рис. 104. Схема соотношений регулярных решеток, образованных различными элементами сетчатки костистых рыб [Подугольникова, Максимов, 1977]
1 — мозаика колбочек; 2—решетка горизонтальных клеток наружного слоя; 3 — решетка горизонтальных клеток среднего слоя; 4 — решетка булавовидных окончаний биполярных клеток; кружки — положения ядер клеток; штриховка — отдельные элементы каждой из решеток
1 — мозаика колбочек; 2—решетка горизонтальных клеток наружного слоя; 3 — решетка горизонтальных клеток среднего слоя; 4 — решетка булавовидных окончаний биполярных клеток; кружки — положения ядер клеток; штриховка — отдельные элементы каждой из решеток
образующих выраженную мозаику [Engstrom, 1963; Бабурина, 1972; Подугольникова, Максимов, 1973, 1977; Steinberg, Reid, Lacy, 1973; АН, Antil, 1976; и др.]. По Вагнеру [Wagner, 1975], экологические факторы играют важнейшую роль в определении типа колбочковой мозаики в сетчатке рыб. Наиболее сложные высокоорганизованные колбочковые мозаики обнаруживаются у пелагических с дневной активностью рыб, зрение которых определяет их жизнедеятельность. По мнению Wagner, регулярные узоры мозаик фоторецепторов могут представлять структурную основу для высокой разрешающей способности как во времени, так и пространственной. По мнению Г. Н. Симкипа [1980], формирование мозаики рецепторов — следствие специализации системы регистрации и анализа движущейся светотени. В сетчатке четырехглазой рыбы — Anableps anableps — мозаика рецепторов существенно отличается в центре и на периферии сетчатки [Borwein, Hollenberg, 1973]. Сетчатка четырехглазой рыбы — один из демонстративных примеров связи строения с экологией [Walls, 1942]. В детальном исследовании Borwein, Hollenberg [1972] смогли весьма убедительно показать, что вентральная и дорсальная сетчатки этой рыбы существенно отличны. Вентральная сетчатка, контролирующая верхнюю часть поля зрения, примерно вдвое толще дорсальной сетчатки, смотрящей вниз и отражающей водную среду. Внутренний плексиформный слой значительно шире, более дробно подразделен, внутренний ядерный слой содержит 10—12 рядов ядер, тогда как в дорсальной сетчатке всего 3—5 рядов. Ганглиозных клеток и фоторецепторов также больше в вентральной сетчатке. Авторы этой ценной работы привели данные о различиях в структуре сетчатки, зависящих от той среды, в которой та или иная часть глаза функционирует наряду с другими фактами о структуре всего глаза в целом. Так, роговица верхнего отдела глаза, смотрящего в воздух, толще и имеет большую кривизну, то же касается и хрусталика.
Замечательный пример приспособления структуры к среде — сетчатка глубоководных рыб [Locket, 1969, 1971, 1977]. Из фоторецепторов у них наблюдаются только палочки с очень длинными наружными сегментами. У глубоководной рыбы Poromitra nigrofulvus (Garman) наружные сегменты палочек образуют два слоя — наружный и внутренний, составляющие вместе 68% всей толщи сетчатки [Locket, 1969]. Большому числу очень длинных палочек соответствует и большое число дисков в каждой палочке, до 6000—15 000 [Винников, 1971; Говардовский, 1978; обзор Бабурина, Белтадзе, 1983]. Для сравнения напомним, чго в сетчатке лягушки в каждой палочке около 1100 дисков, в колбочке— около 750. Внутренний ядерный слой и слой ганглиозных клеток отличаются очень малым количеством клеточных тел. Locket [1977], а ранее Duke-Elder [1958] считали, что такие особенности конструкции связаны с высокой степенью суммации фоторецепторных сигналов, что обусловливает высокий уровень светочувствительности при низкой остроте зрения.
У птиц, многие из которых, как известно, обладают высокой остротой зрения, отмечены и уникальные особенности в структурной организации зрительной системы, многие из которых могут быть связаны с условиями их существования [Walls, 1942; Авилова, 1980]. У небольших птиц вес глаз часто превосходит вес мозга. В сетчатке птиц, отличающихся быстротой и маневренностью полета, например сокола, ласточки и других, две ямки — центральная и темпоральная. Из 13 видов птиц семейства чайковых у полярной, пестроносой и чайконосой крачки, преследующих мелкую подвижную добычу, обнаруживается очень глубокая центральная ямка [Авилова, 1976, 1980]. Развитие внутреннего ядерного слоя сетчатки птиц достигает величин, не наблюдающихся ни у каких других позвоночных [Rochon-Duvigneaud, 1943; Walls, 1942; Detwiler, 1943]. В частности, у малиновки (Turdus migratorius) клетки наружного ядерного слоя расположены в 3 рядах, внутреннего ядерного слоя — в 28 рядах, у ястреба — в 4 и 20 рядах соответственно [W'alls, 1942]. У серебристой чайки в области фовеального валика клетки внутреннего ядерного слоя расположены в 20 —25 рядах [Авилова, 1980]. Для сравнения можно обратиться к рис. 1, где в сетчатке кошки имеется противоположное соотношение; многие примеры можно также почерпнуть у Detwiler [1943], где автор приводит фотографии сетчатки многих позвоночных, ведущих самый различный образ жизни.
Интерес представляет рассмотрение особенностей морфологии горизонтальных клеток, в частности особая их величина у некоторых позвоночных. У некоторых осетровых рыб эти клетки настолько велики, что другие нейроны внутреннего ядерного слоя занимают очень небольшую его часть [Walls, 1942]. У многих костистых рыб также отмечены весьма большие горизонтальные клетки [Бабурина, 1972; Подугольникова, Максимов, 1973; и др.].
Связать большую величину горизонтальных нейронов с водной средой обитания не удается, так как у многих костистых рыб горизонтальные клетки обычной величины [Walls, 1942; Бабурина, 1972]. Объяснение Ramon у Cajal [1972], считающего, что большой объем этих клеток связан с большим количеством палочек, также не удовлетворителен; в палочковой сетчатке многих представителей позвоночных нет крупных горизонтальных клеток. Закономерность связи величины нейрона с диаметром аксона и быстротой проведения по нему возбуждения, подтверждающаяся в других отделах нервной системы, в данном случае вряд, ли соответствует истине. Аксоны горизонтальных клеток коротки,, а у некоторых форм этих нейронов аксоны отсутствуют [Parthe„ 1972; Gallego, 1971а; Boycott, 1974].
Как известно, горизонтальные клетки одного и того же ряда у рыб связаны друг с другом многими щелевыми контактами [Yamada, Ishikawa, 1965; Бызов, 1984; и др.]. Вероятно, большая поверхность слияния очень крупных тел друг с другом может способствовать более синхронному вовлечению их в деятельность, соответственно мнению Bennet [1972] о роли электрических синаптических контактов.
Детально исследовалось строение сетчатки и экология у различных видов грызунов. Весьма четкие различия по строению и соотношению толщины слоев сетчатки у белок, сусликов и крысы нашла Tansley [1961], сопоставлявшая структуру с электроретинограммой. По ее данным, у белок и сусликов наружный ядерный слой на поперечных срезах узок и состоит лишь из двух рядов ядер колбочек; внутренний ядерный слой широкий, ядра в нем расположены плотно. Слой ганглиозных клеток содержит несколько рядов тел нейронов, особенно плотно расположенных в центральном поле. Между сетчатками чисто дневных белок, белок с ночной активностью и сусликов имеются четкие различия. У крысы в сетчатке преобладают палочки и соотношение слоев соответственно существенно иное. Строение сетчатки и глаза в целом у белки, сурка и морской свинки сравнивала Ж. М. Кухтина [1963]. У белки сетчатка на поперечных срезах широкая, с преобладанием колбочек в отношении 4—5 к одной палочке. В среднем на 1 ганглиозную клетку приходится 5—7 клеток внутреннего зернистого слоя и 4—6 палочек и колбочек. Центральное поле весьма многоклеточно. Сетчатки сурка и морской свинки качественно отличаются от сетчатки белки, причем различия рецепторного и нервного отдела глаза связаны со структурой глаза в целом (размеры и форма хрусталика и т. д.).
Морфология сетчатки белки представляет большой интерес ввиду высоко развитой у нее зрительной функции в сочетании с прекрасной моторикой. Сложный зрительный акт белки включает распознавание множества компонентов стимула: его цвета [Орлов, 1971], направления движения [Cooper, Robson, 1966], мелких его деталей, глубины расположения, величины и др. Изучение нейронов сетчатки подтвердило сложную ее дифференцировку: большое количество подслоев во внутреннем плексиформном слое, множество двух- и трехслойных ганглиозных клеток, особое разнообразие диффузных клеток.
Значительное своеобразие отмечено и в отношении зрительных центров у белки. Так, физиологами выявлен примечательный факт: для многих нейронов зрительной коры белки оптимальными являются скорости движения значительно более высокие, чем для нейронов коры кролика [Масс, Полкошников, Супин, 1973]. Нейроны, реагирующие на направление движения, специфически чувствительны к высоким скоростям движения порядка сотен градусов в секунду [Супин, 1975, 1981]. По данным А. Я. Супина, в зрительной коре белки можно выделить две системы нейронов, одна из которых ведает определением направления и скорости движения стимула, т. е. его динамических характеристик, другая — ориентацией контуров и линий, т. е. его статических характеристик. Способность анализировать быстро движущиеся изображения возникла, по мнению автора, как необходимое приспособление к особенностям движения белки, в частности для корректировки полета в прыжке, так виртуозно осуществляемого этим грызуном.
Хорошо развитая ассоциативная деятельность, точно рассчитанные прыжки и хорошее зреште соответствуют как сложности конструкции сетчатки, так и сложности и «приматному» типу первичного коркового зрительного поля 17. Поле 17 в коре мозга белки обнаруживает весьма высокую степень диффереицировки; еще М. О. Гуревич, Г. X. Быховская, Я. А Ураповский [1929] отметили его «приматный тип», характеризующийся расслоением слоя IV на 3 подслоя. Это наблюдение было подтверждено при изучении цитоархитектоники [Кухтина, 1963] и нейронов поля 17 [Школьник-Яррос, 1968 б]. В подслое IV — множество крупных и средних звездчатых клеток Кахаля, сходных с таковыми в поле 17 низшей обезьяны [Школьник-Яррос, 1955]. Поле 17 кошки качественно отличается; слой IV широк и клетки в нем расположены плотно, но без четкого подразделения на 3 подслоя. Звездчатые клетки Кахаля также отличны по характеру дендритного ветвления [Ramon у Cajal, 1923; Школьник-Яррос, 1965а]. Подразделение слоя IV на 3 подслоя в коре мозга приматов рядом авторов связывалось с механизмом бинокулярного зрения. Нет сомнения, что образу жизни белки, ее совершенной моторно-зрительной координации должна соответствовать высокая степень диффереицировки коркового зрительного центра, чем и может объясняться «приматный» тип поля 17.
Особенности в структуре различных звеньев зрительной системы в связи с образохм жизни обнаружены и у представителей отряда млекопитающих — насекомоядных [Шалагина, 1974а, б; и др.]. У землеройки, выхухоли, крота и ежа, ведущих разный образ жизни и отличающихся свойствами зрения, заметно отличаются и зрительные центры. Как пример приводим такие отличия у крота и ежа. У крота отмечено отсутствие слоистости в передних бугорках четверохолмия, узкая кора зрительной области мозга, малая дифференцированность слоев коры. Значительно шире и более дифференцирована зрительная кора ежа, в передних бугорках четверохолмия намечается слоистость.
Можно наблюдать весьма заметные различия в строении зрительных центров у обезьян в соответствии с их различным образом жизни и своеобразием в свойствах зрения. Многократно отмечены в литературе особенности наружного коленчатого тела и area striata у ночных и дневных обезьян [Walls, 1942; Школьник-Яррос, 1962; и др.], выражающиеся в характере и степени расслоенности этих образований, в качественном своеобразии нейронов.
Как обнаружено В. П. Зворыкиным [1960, 1971], существует зависимость между структурными особенностями подкорковых образований и экологией каждого вида. По В. П. Зворыкину, такая зависимость особенно ясно выражена на уровне ядер зрительной и слуховой систем, а па корковом уровне выявляется мало. Не удивительно с этой точки зрения, что у приматов существует несколько вариантов строения наружного коленчатого тела. Так, если закономерно наличие 6 слоев в этом ядре у большинства приматов, включая человека, то у обезьян Aotus, Callithris, Tarsius лишь 4 слоя в наружном коленчатом теле [Naback,1975]. Структура проекций зрительных волокон на наружное коленчатое тело также различается. У Haplorhini контралатеральные зрительные волокна оканчиваются в 1, 4, 6 слоях, ипсилатеральные — в 2, 3, 5 слоях. У других приматов — в 1, 5, 6 и 2, 3, 4 соответственно. У насекомоядного тупайи (как считают некоторые авторы — предшественника приматов) контралатеральные волокна проецируются на 1, 3, 4, 5 слои, ипсилатеральиые — на 2, 6 слои [Noback, 1975]. Автор предполагает, что эти три типа проекций возникли в филогенезе независимо; функциональное значение таких существенных различий пока неизвестно.
Связь экологии с морфологией зрительных центров можно наблюдать не только у млекопитающих, но и у низших позвоночных. Так, существенные отличия в нейронной организации крыши среднего мозга — основном зрительном центре — обнаружены у сухопутных черепах, греческой (Testudo Graeca), степной (Testudo Horsfieldi) и болотной черепахи (Ernys orbicularis) [Гончарова, 1976, 1979, 1980; Давыдова и Гончарова, 1970]. В частности, у сухопутных черепах, греческой и степной, горизонтальные нейроны формируют широкие системы в верхних слоях tectum; плотность клеток на уровне подслоя «б» слоя I в 2—2,5 раза выше; в слое II наблюдаются нейроны с короткими дендритами, оканчивающимися в исходном слое; у некоторых клеток глубокого слоя V апикальные дендриты не достигают слоя I. Все перечисленные морфологические особенности и некоторые другие присущи сухопутным черепахам и отсутствуют у болотной. Авторы предполагают, что такие различия в нейронной организации крыши среднего мозга трех изученных видов черепах связаны с особенностями среды обитания и характером питания, т. е. экологией. Существенные различия между сухопутной, степной и болотной черепахами имеются также в синапсоархитектонике крыши среднего мозга [Давыдова, Гончарова,, 1977].
----
Статья из книги: Нейроны сетчатки | Е.Г. Школьник-Яррос, А.В. Калинина

Комментариев 0