Эволюционный аспект


Описание
Не все структурные вариации сетчатки и других отделов зрительной системы удается объяснить экологией видов. Обратимся снова к выполненному современным электронно-микроскопическим методом исследованию синаптической организации сетчатки у позвоночных различного уровня эволюционного развития [Dowling, 1968]. Как оказалось, количество синапсов во внутреннем плексиформном слое сетчатки у лягушки— 10 400 000 на 1 мм2 тогда как в сетчатке человека (2 900 000 на 1 мм2). Обнаружены и весьма существенные качественные различия: во внутреннем плексиформном слое сетчатки лягушки отростки очень малы (1,5—2 мкм в диаметре), они образуют в 4 раза больше амакриновых синапсов, среди которых много сериальных, чем в сетчатках приматов и кошки.Принимая количество и особенности синапсов во внутреннем плексиформном слое за показатель уровня эволюционного развития, автор предлагает следующий ряд позвоночных, в котором они расположены по степени сложности структурной организации сетчатки: простая сетчатка—у приматов и кошки, промежуточная— у кролика и белки, сложная — у лягушки и голубя. По Dowling, тенденция к сложности сетчатки коррелирует с меньшей энцефализацией функции нервной системы, так что у человека и обезьяны, у которых кора мозга панболее развита, сетчатка самая простая. У лягушек и голубей, у которых кора мозга наименее развита, сетчатка самая сложная.
Наблюдается также корреляция со сложностью физиологических ответов ганглиозных клеток: например, в сетчатке лягушки отдельные клетки отвечают на очень специфические черты зрительного образа [Lettvin et al., 1961; Пигарев, 1970; Gordon, Hood, 1976; Grusser, Grusser-Cornehls, 1976; Мантейфель, 1977]. По схеме, предложенной Dowling [1968], основные особенности связей в сетчатке лягушки — преобладание амакриново-амакриновых сериальных синапсов и связь ганглиозных клеток с биполярными через вставочный амакриновый нейрон. Основные выводы Dowling подтверждает Dubin [1970] в работе, проведенной на большем числе различных по уровню
эволюционного развития позвоночных также методом количественной электронной микроскопии внутреннего плексиформного слоя.
Физиологами обнаружены принципиальные различия в функциональной организации зрительной системы у крайних представителей эволюционного ряда позвоночных [Праздникова, 1966; Глезер, 1975]. У низших позвоночных (рыб, лягушек) система детекторных нейронов хорошо развита уже в сетчатке, отсутствует формирование абстрактного зрительного образа; в отличие от высших животных они оперируют конкретными изображениями. Возможности мозга низших представителей эволюционного ряда жестко лимитированы, у них отсутствует сенсорное обучение и осуществляется прямой выход из сенсорной системы на поведенческий уровень.
У высших позвоночных существует мир абстрактных образов, велика инвариантность образа, т. е. опознание устойчиво к ряду его преобразований.
Рассмотрим полученные нами данные о нейронной организации зрительной системы в свете этих исследований. Сетчатка лягушки (Rana temporaria), несомненно, сложнее, чем сетчатка кошки, не только по особенностям синапсов, но также по конструкции нейронов. Сложность особенно проявляется в дробной дифференцировке внутреннего плексиформного слоя. Весьма разнообразны диффузные ганглиозные клетки — можно выделить пе менее пяти их разновидностей, различающихся по размаху, густоте, тонкости дендритных ветвлений. Диффузные ганглиозные клетки в сетчатке кошки наблюдаются значительно реже.
Сложной по архитектонике и нейронному строению можно считать и сетчатку многих рыб. Так, у костистых рыб обнаружено до 5 типов фоторецепторов, часто расположенных с четкой регулярностью и образующих мозаику [Бабурина, 1972; Подугольника и Максимов, 1977]. У некоторых рыб найдено не менее 10 типов биполярных клеток [Scholes, 1975]; число рядов и соответственно типов горизонтальных клеток у многих рыб также велико, достигая 4 [Parthe, 1972]. Выявлена дробная дифференцировка внутреннего сетчатого слоя Подугольникова [1981]). Особенно следует подчеркнуть многослойные амакриновые клетки, наблюдавшиеся нами в сетчатке щуки [Школьник-Яррос, Подугольникова, Дюбина, 1975].
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
В соответствии со сложной морфологической организацией в сетчатке костистых рыб обнаружены избирательно реагирующие на специфические стимулы ганглиозные клетки; например, на направление движения [Зенкин, Ппгарев, 1969], цвет раздражителя [Орлов, 1971]. Еще большее разнообразие нейронов найдено в сетчатке рептилий и птиц [Ramon у Cajal, 1972; Kolb,.1982].
Изучение нейронного строения сетчатки суслика [West, 1976} и белки (Школьник-Яррос, 1974) позволяет видеть значительную диффереицированность и многообразие типов нейронов. Если по количеству синапсов внутреннего сетчатого слоя, по Dowling, беличьи занимают промежуточное положение между лягушкой и приматами, то по нейронному строению эта сетчатка ближе к сложным. Как пример типичного двухслойного нейрона с расположением дендритных ветвлений на двух уровнях внутреннего сетчатого слоя может служить рисунок ганглиозной клетки сетчатки белки. Факт весьма дробной дифферепиировки внутреннего плексиформного слоя подчеркивал Walls [1942] и у другого вида, близкого к семейству беличьих,луговой собачки (Cynomys ludovicianus).
Сравнение данных о нейронном строении сетчатки различных позвоночных с данными Polyak [1941, 1957] и Boycott, Dowling [1969] о нейронном строении сетчатки приматов позволяет выделить специфические черты, свойственные зрительной системе приматов. Прежде всего—это карликово-колбочковая система, включающая карликовые биполярные клетки, карликовые ганглиозные клетки [Polyak, 1941, 1957; Dowling, Boycott, 1966; Boycott, Dowling, 1969; Kolb, Boycott, Dowling, 1969; Kolb, 1970], карликовые элементы наружного коленчатого тела и зрительной коры приматов [Школьник-Яррос, 1962, 1965а]. Эта система отсутствует у кошки. Хотя метод Гольджи не дает права ввиду избирательности импрегнации окончательно утверждать отсутствие любого элемента, тем не менее полученные в данном исследовании факты, а также литературные данные о биполярных клетках сетчатки кошки [Boycott, Kolb, 1973b] позволяют думать, что карликовые биполяры, а также карликовые ганглиозные клетки в сетчатке кошки отсутствуют.
Вторым важнейшим для понимания конструкции зрительной системы приматов признаком является особое развитие корковых зон, окружающих первичное зрительное поле 17, area striata, у человека [Филимонов, 1949а] и иной характер связей этих полей в зрительной системе. Так, если у грызунов, хищных волокна из наружного коленчатого тела приходят не только в area striata (поле 17, зона VI), но и в поле 18 (зона VII), то у приматов волокна из наружного коленчатого тела прослеживаются только в поле 17; поле 18 (зона VII) пе получает прямых путей из этого ядра и связано с полем 17 кортико-кортикальными волокнами [Супин, 1981].
Третий признак, свидетельствующий о чрезвычайном значении зрительного восприятия для приматов,— еще большее, чем у других млекопитающих, распространение представительства зрения по многим зонам коры мозга [Van Essen, 1979]. Если же включить в функцию зрительное восприятие, внимание, необходимое для сознательного его выполнения, последующее программирование действий, включающих зрительный акт, то морфологическим базисом явятся лобные отделы мозга [Лурия, 1966].Так как любое действие, сопровождаемое зрением, включает также глазодвигательные реакции [Ярбус, 1965; Леушина, 1971], то не будет преувеличением сказать, что значительная часть гигантского мозгового механизма участвует в зрительном процессе. У человека этот механизм, входящий в систему сознательных, творческих, логических операций, достигает наивысшего расцвета.
Таким образом, по некоторым весьма существенным признакам структурной организации зрительная система кошки и человека существенно отличаются. Если объединение кошки и приматов в одну группу по характеру синаптических связей сетчатки, по Dowling, возможно, то рассмотрение зрительной системы целиком заставляет склониться к мнению о качественных их отличиях, причем употребление терминов «простота» и «сложность» нежелательно тем более, что физиологи неоднократно демонстрировали более сложные и разнообразные рецептивные поля ганглиозных клеток в сетчатке кошки [Stone, Fabian, 1966; Spinelli, 1967].
Вернемся к сравнению зрительной системы лягушки и кошки в плане развития зрительных центров. Как уже упоминалось, неокортекс у лягушки имеется лишь в виде крошечного зачатка латерального паллиума [Карамян, 1976]. Основная масса зрительных волокон, проецируясь «точка в точку», оканчивается в крыше среднего мозга. Немногочисленные зрительные волокна оканчиваются в ядрах ствола и межуточного мозга: претектуме, латеральном коленчатом теле, ядре Беллопчи, заднеталамическом ядре, крючковидном ядре, преоптическом ядре, тегментуме. По А. И. Карамяну [1976], у амфибий впервые появляются два пути проведения зрительной информации: ретино-таламо-телепцефалический и ретино-текто- таламо-теленцефалический. Однако зрительные проекции к коре мозга у амфибий оканчиваются в гиппокампе.
У кошки, как известно, по многочисленным исследованиям морфологии, физиологии зрительных центров и путей, в мощно развитой зрительной коре мозга обнаруживаются нейроны с простыми, сложными и сверхсложными рецептивными полями, выраженное колончатое строение [Hubei, Wiesel, 1962, 1974, 1977; и др.]. Зрительные проекции широко представлены в цитоархи- тектонических полях 17, 18, 19, а также в соседних, височной, сплениальный, теменной областях коры мозга. Основная масса зрительных волокон из сетчатки оканчиваются у кошки в наружном коленчатом теле, весьма высокоразвитом, многослойном ядре, образуя синаптические гломерулярные комплексы [Szentagothai, 1963]. Другие зрительные сетчаточные волокна прослеживаются в верхнем двухолмии, многих
подкорковых ядрах и стволе мозга.
Следовательно, наиболее перспективный подход для понимания относительного упрощения структуры сетчатки у многих млекопитающих по сравнению с сетчаткой многих рыб, некоторых амфибий, рептилий, птиц — рассмотрение зрительной системы в целом. С полным основанием Maturana et al. [1960], Dowling [1968], Shepherd [1974] и многие другие подчеркивают сложность интегрированных реакций на зрительные стимулы в зрительных центрах у высших позвоночных.
Относительное упрощение и перестройку структуры и функций можно наблюдать от tectum opticum низших позвоночных к верхнему двухолмию млекопитающих. Так, tectum opticum амфибий, рептилий, птиц характеризуется чрезвычайно ярко выраженной многослойностыо (до 15 слоев у птиц), густотой расположения нейронов в клеточных слоях, сложностью и многообразием (рис. 105)
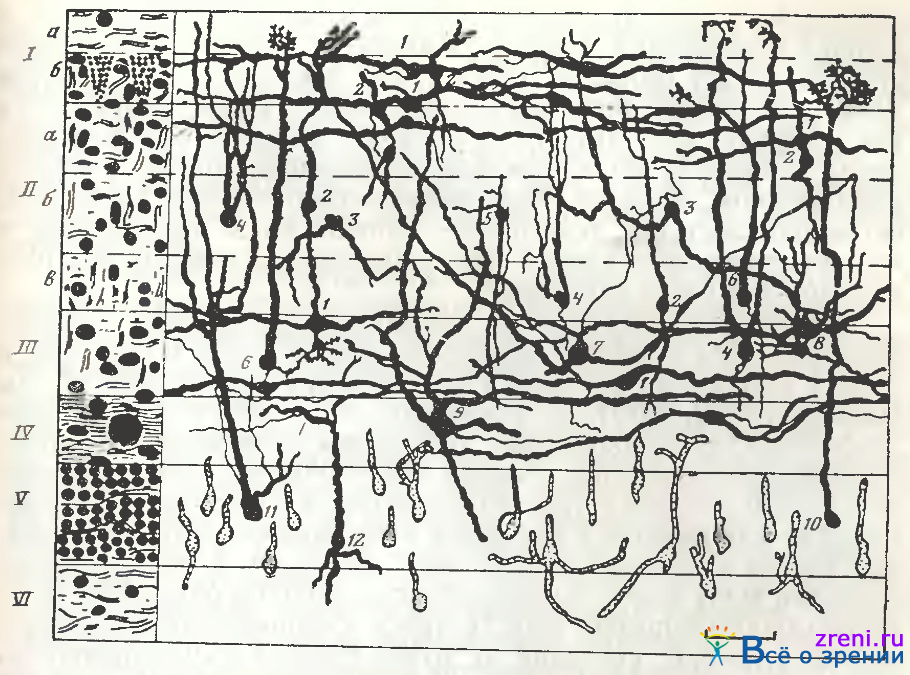
Рис. 105. Схема нейронной организации крыши среднего мозга степной черепахи [Гончарова, 1976]
Слева — метод Ниссля; справа — метод Гольджи, рисовальный аппарат; I—VI—слои; а, б, в — подслои: 1, 2 — горизонтальные и вертикальные веретена; 3, 4, 5, 6, 7, 12 — нейроны с различной морфологией дендритных ветвлений; 8 — звездчатые клетки; 9 — гигантские клетки; 10 —клетка с концевым ветвлением дендрита в виде кисточки; И —клетка без концевой кисточки
Слева — метод Ниссля; справа — метод Гольджи, рисовальный аппарат; I—VI—слои; а, б, в — подслои: 1, 2 — горизонтальные и вертикальные веретена; 3, 4, 5, 6, 7, 12 — нейроны с различной морфологией дендритных ветвлений; 8 — звездчатые клетки; 9 — гигантские клетки; 10 —клетка с концевым ветвлением дендрита в виде кисточки; И —клетка без концевой кисточки
нейронов [Ramon, 1890; Ramon у Cajal, 1911; Сепп, 1949; Гончарова, 1976, 1979]. У млекопитающих в верхнем двухолмии часть слоев полностью исчезает, особенно это касается верхних отделов двухолмия. Это изменение связано с развитием ретино-таламо-кортикальной системы и приходом значительной части зрительных афферентов в наружное коленчатое тело и далее в кору мозга. Высокодифференцированные верхние слои tectum opticum, явчяясь основным зрительным центром у низших позвоночных, у млекопитающих в значительной степени утрачивают эту функцию. Однако, как показал И. В. Викторов?[1969], об упрощении в истинном смысле нельзя говорить, так как в верхние слои двухолмия приходят корково-тектальные волокна и таким путем осуществляется корковый контро ib многих рефлекторных функций. Чем выше млекопитающее по филогенетической лестнице, тем больше влияние коры на подкорку, и соответственно появляются новые признаки в нейронной структуре верхних слоев двухолмия.
Подчеркнем несомненную важность внутреннего плексиформного слоя сетчатки для определения степени сложности и уровня интеграции функций в данном звене зрительной системы [Dowling, 1968, 1979]. Во внутреннем плексиформиом слое сходятся отростки амакриновых, биполярных, ганглиозных, иитерплексиформных клеток, а также центробежные волокна сетчатки. Именно эта структура характеризуется образованием синаптических комплексов, а входящие в них многослойные ганглиозные клетки обладают наибольшим сходством с пирамидными клетками коры мозга. Многослойность, наблюдающаяся во внутреннем плексиформиом слое сетчатки и особенно выраженная у многих амфибий, рептилий и птиц, придает ему сходство со строением tectum opticum низших позвоночных и со зрительной корой дневных приматов. Современные электронномикроскопические данные позволяют расшифровать структурную организацию экранных центров. Для сетчатки, например характерны многочисленные и многократно повторяющиеся микрокомплексы, в которые входят непременной частью сериальные синапсы [Kidd, 1962], реципрокпые синапсы [Dowling. Boycott, 1965], триады [Dowling, Boycott, 1966].
Рассуждая о направлениях эволюции зрения, Walls [1942] приходит к выводу о сходстве глаза плацентарных животных с глазом амфибий (не связывая этот факт с линией развития от амфибий к плацентарным, а объясняя его значительным упрощением глаза во время длительно наблюдавшегося ночного периода его функционирования). Walls считает, что значительное упрощение глаза млекопитающих по сравнению с глазом их непосредственного предка — рептилий привело как бы к возврату эволюции и внешнему подобию с глазом амфибий. Рассмотренные выше данные о синапсах внутреннего плексиформного слоя [Dowling, 1968; Dubin, 1970], а также о нейронном строении сетчатки лягушки и кошки не позволяют полностью согласиться с гипотезой Walls о сходстве глаз (и сетчатки) амфибий и млекопитающих. Если принять сетчатку домашней кошки за исходную для сравнения, то приведенные рисунки демонстрируют отчетливые отличия ее от сетчатки травяной лягушки, начиная с весьма существенных количественных и качественных отличий фоторецепторов и кончая ганглиозными клетками и внутренним плексиформным слоем. Однако наше возражение не относится к структуре сетчатки всех без исключения млекопитающих и сравнение ее структурной организации у лягушки и белки действительно выявляет сходство, особенно в комплексе: амакриновые, ганглиозные клетки и внутренний плексиформный слой. Нельзя согласиться с Walls, что глаз амфибий прост, в этом еще одна его ошибка, и совершенно ясно кроме того, что нельзя переносить данные по глазу одного вида млекопитающих на другие виды этого класса.
Положение Я. Сентаготаи и М. Арбиб [1976], считающих, что в сетчатке нейроны получают в основном однородный входной сигнал на все дендритные ветви, следует рассматривать также в свете различий между строением сетчатки у низших и высших позвоночных. Так, ганглиозные клетки с однослойными ветвлениями дендритов, преимущественно наблюдающиеся в сетчатках у высших млекопитающих и человека, могут действительно получать однородный сигнал, проходящий через биполяры и сходные типы амакриновых клеток. Но многослойные ганглиозные клетки, чаще встречающиеся в сетчатках амфибий, рептилий, птиц [Ramon у Cajal, 1972; Kolb, 1982], по-видимому приспособлены к восприятию разнородных сигналов, идущих через биполярные клетки разных типов, однослойные амакриновые клетки разных подслоев внутреннего плексиформного слоя, многослойные амакриновые клетки.
Можно отметить корреляцию простоты или сложности в структуре сетчатки не только со степенью развития мозга в целом, но и со способностью к выполнению задач по экстраполяции [Крушинский, 1977]. Л. В. Крушинский при помощи оригинального и вполне адекватного для изучения элементарной рассудочной деятельности метода установил огромные различия в успехе решения экстраполяционных задач животными разных таксономических групп. Так, весьма показательно с этой точки зрения полное отсутствие способности к экстраполяции у бесхвостых амфибий— травяной лягушки и жабы — при наличии весьма сложной сетчатки и слабом развитии мозга. Кошки же вполне удовлетворительно справляются с экстраполяционными задачами; сетчатка у них проще, а мозг велик. Представляет интерес, что у голубей, при высокой дифференпированности сетчатки [Boycott, Dowling, 1969; Бекчанов, 1980], так же как у травяной лягушки, полностью отсутствует способность к выполнению экстраполяционных проб.
Однако такая закономерность — сочетание сложно дифференцированной сетчатки с отсутствием способности к экстраполяции обнаруживается не всегда. Так, у врановых птиц элементарная рассудочная деятельность развита очень хорошо. По способности к экстраполяции вороны не уступают хищным млекопитающим [Крушинский, 1977].
Морфологические корреляции таких различий между голубями и врановыми обнаруживаются при изучении структуры их мозга. Нейронная структура неостриатума ворон отличается чрезвычайно разнообразным нейронным составом, включая длинноаксонные, короткоаксонные клетки разных типов, малые, крупные, различной формы и ветвистости. Среди длинноаксонпых клеток в неостриатуме ворон выделяются нейроны, дендриты которых чрезвычайно богаты протоплазматическими выростами — шипиками [Доброхотова, 1969, 1971; Крушииский, Доброхотова,. Школьник-Яррос, 1985]. Подобных нейронов в стриатуме голубей не обнаружено.
В заключение подчеркнем еще раз важность рассмотрения не только зрительной системы в целом, но и уровня организации всего мозга. Исследование эволюции зрительной системы и других систем у отдельных представителей различных классов позвоночных дает основополагающие представления о закономерностях и общих направлениях изменений структурно-функциональной организации. Однако лишь изучение структуры и функции органа у родственных форм в сравнительном аспекте выявляет значение экологии для формирования адаптивных свойств зрительной и других систем [Матвеев, 1967]. Наиболее убедительно значение экологии выявляется при систематическом изучении онтогенеза видов; яркий пример такого подхода по отношению к сетчатке, строение которой тесно связано с образом жизни, приведен в начале главы по работам Е. А. Бабуриной [ 1972].
Упомянутые выше, обнаруженные физиологами [Праздникова, 1966; Глезер, 1975] принципиальные отличия в зрительном восприятии у крайних представителей эволюционного ряда коррелируют со структурной организацией зрительной системы. Так, у высших животных и человека формируется абстрактный зрительный образ, они способны к значительному сенсорному обучению, велика инвариантность образа, в то время как у низших позвоночных все эти свойства резко уменьшены или отсутствуют. Морфологически в этом плане важнее всего соотношение развития структур глаза (сетчатки) и мозга. У низших представителей позвоночных отсутствует корковое звено зрительной системы и преобладает периферическое, у высших относительно уменьшается значение и сложность сетчатки. Наибольшего развития центральные отделы зрительной системы достигают у человека; в зрительном процессе — восприятии сигналов, переработке полученной информации и последующих действиях, включающих и акты с участием зрения и глазодвигательные реакции, участвует почти весь гигантский мозговой механизм.
----
Статья из книги: Нейроны сетчатки | Е.Г. Школьник-Яррос, А.В. Калинина

Комментариев 0