Дирекциональная избирательность

Содержание:
Описание

Многие сложные клетки лучше реагируют на движение стимула в одном направлении, чем в противоположном. Различие в реакции часто бывает весьма резким — при одном направлении движения возникает энергичный ответ, а при обратном направлении клетка вообще не отвечает (рис. 48). Как выяснилось, примерно 10—20% клеток в верхних слоях стриарной коры проявляют заметную дирекциональную избирательность. Остальные клетки, по-видимому, не обладают такой избирательностью — мы тщательно исследовали ответы клеток с помощью ЭВМ, пытаясь обнаружить хотя бы небольшое различие в ответах на движение стимула в противоположных направлениях. Таким образом, существуют, видимо, два разных класса клеток — один из них четко проявляет дирекциональную избирательность, другому она не свойственна.
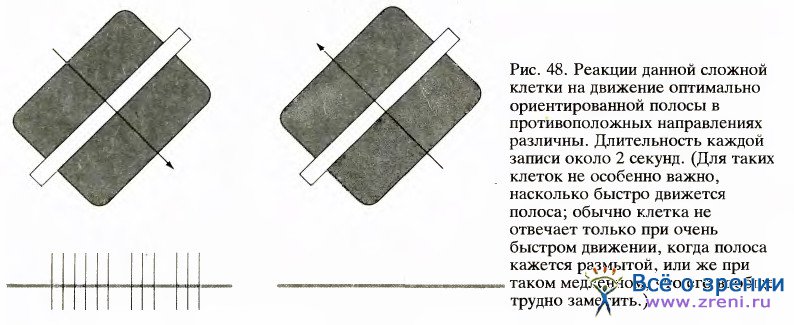
Если прослушивать импульсную реакцию клетки с сильно выраженной дирекциональной избирательностью, то создается впечатление, что при движении линии в одном направлении стимул как бы резко подталкивает клетку и заставляет ее разряжаться, а при движении в противоположном направлении происходит как бы сбой и стимул становится неэффективным (это напоминает ситуацию, когда вы, заводя часы, поворачиваете головку заводного механизма назад и раздается характерный треск).
Мы не знаем, как устроены входные сети таких клеток с дирекциональной избирательностью. Возможно, что на вход такой клетки подключены простые клетки, реакции которых на движение стимула в противоположных направлениях неравнозначны, асимметричны. Рецептивные поля таких простых клеток асимметричны.
Другую возможную схему предложили в 1965 году Г. Барлоу и У. Левик для объяснения дирекциональной избирательности некоторых клеток в сетчатке кролика — клеток, которых, по-видимому, нет у обезьяны. Если применить их схему к сложным клеткам коры, то нужно предположить наличие промежуточного слоя клеток, расположенных между простыми и сложными клетками (как на рис. 49).
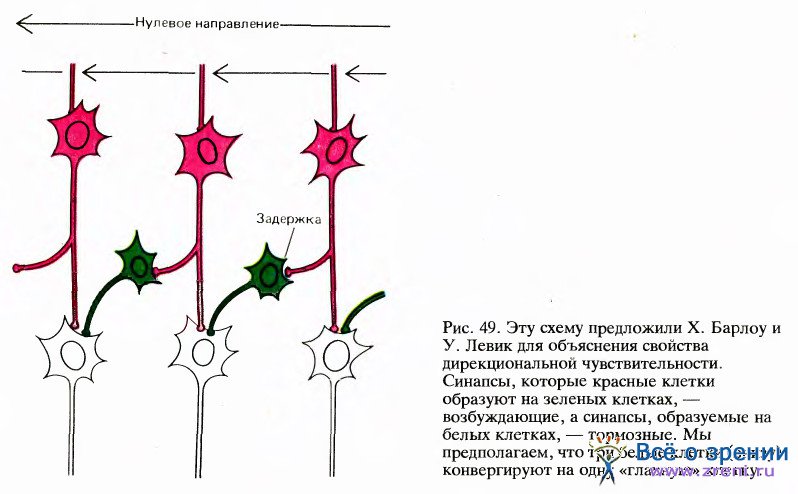
Представим себе, что клетка из промежуточного слоя имеет возбуждающий вход от одной простой клетки и тормозный вход — от другой клетки, рецептивное поле которой непосредственно примыкает к предыдущему, и притом всегда с одной стороны. Предположим далее, что тормозный путь включает некоторую задержку, возможно, за счет включения еще одной промежуточной клетки. В таком случае, если стимул движется в одном направлении, скажем справа налево (как на рис. 49, иллюстрирующем модель Барлоу и Левика), то промежуточная клетка возбуждается сигналом с одного из своих входов как раз в тот момент, когда действует торможение, приходящее от другой клетки, рецептивное поле которой только что пересек стимул. Возбуждающий и тормозный эффекты нейтрализуют друг друга, и в результате клетка не дает импульсного разряда. Если же стимул движется в обратном направлении, торможение возникает слишком поздно для того, чтобы предотвратить импульсный разряд. Если множество таких промежуточных клеток конвергирует на какую-то клетку, расположенную на следующем, третьем, уровне, то эта клетка будет обладать свойствами сложной клетки с дирекциональной избирательностью.
У нас нет прямых доказательств правильности тех или иных иерархических схем, объясняющих наблюдаемые реакции клеток, т. е. схем, в которых клетки на каждом следующем уровне организуются на основе элементов предыдущего уровня. Тем не менее у нас есть веские основания считать, что нервная система организована по иерархическому принципу. Одно из самых серьезных оснований для такого заключения связано с морфологией — например, у кошек простые клетки сосредоточены в четвертом слое стриарной коры, т. е. в том слое, который получает входы от клеток наружных коленчатых тел, тогда как сложные клетки располагаются в слоях коры выше и ниже — на одно или два синаптических переключения дальше от входа. Таким образом, хотя мы не можем дать точную схему связей на каждом уровне, у нас есть достаточные основания полагать, что такая схема существует.
Главная причина того, что в сложных клетках мы видим результат определенной организации клеток с круглыми рецептивными полями с центром и периферией, — это очевидная необходимость производить переработку информации логически в два этапа. Я хотел бы подчеркнуть слово логически, поскольку в целом нужное преобразование, по-видимому, могло бы быть физически осуществлено и в один этап — если суммировать на отдельных разветвлениях дендритов сложных клеток входные сигналы от клеток с круглыми рецептивными полями. В этом случае каждое разветвление выполняло бы функции отдельной простой клетки, посылая к телу клетки (и соответственно к аксону) сигналы электротоническим путем (с помощью пассивного электрического процесса) всякий раз, когда какая-то линия попадет в определенную зону соответствующего рецептивного поля. В этом случае такая клетка сама проявляла бы свойства сложной клетки. Однако само наличие простых клеток заставляет думать, что мы не должны строить таких сложных воображаемых конструкций.
↑ Значение клеток, чувствительных к движению (включая некоторые комментарии к тому, как мы «видим»)
Почему столь многочисленны клетки, чувствительные к движению? Прежде всего приходит в голову, что такие клетки сигнализируют о том, имеется ли в поле зрения движущийся объект. Для животных, в том числе и для нас с вами, изменения внешнего мира гораздо важнее, чем подтверждения его неизменности, идет ли речь о выживании хищника или жертвы. Поэтому не удивительно, что большинство корковых клеток лучше реагирует на движущиеся объекты. Следуя этой логике, вы должны теперь усомниться — как же тогда мы проводим анализ неподвижных зрительных сцен, если в интересах чувствительности к движению так много клеток, настроенных на определенную ориентацию стимула, нечувствительно к неподвижным контурам. Чтобы получить ответ на этот вопрос, нужно сделать небольшое отступление и остановиться на некоторых важных, но противоречащих интуиции данных относительно того, как мы «видим».
Прежде всего естественно было бы ожидать, что при обследовании окружающего мира наши глаза будут плавно сканировать всю сцену непрерывными движениями. На самом же деле при фиксации какого-то объекта происходит вот что: сначала мы устанавливаем глаза так, чтобы изображение этого объекта попало в область центральной ямки обоих глаз, затем мы удерживаем глаза в таком положении в течение короткого времени, скажем полсекунды, потом глаза скачком перемещаются в новую позицию и фиксируют новую мишень, которая находится где-то в другом месте зрительного поля и привлекает к себе внимание тем, что несколько сдвигается относительно фона или имеет какую-то интересную форму. Во время такого скачка, или саккады, скорость движения глаз столь велика, что зрительная система не успевает отреагировать на перемещение изображения по сетчатке и мы его просто не замечаем. (Возможно, что в некотором смысле зрение отключается на период скачка с помощью какой-то сложной нейронной схемы, которая связывает глазодвигательные центры с главным зрительным путем.) Таким образом, процесс обследования зрительного поля при чтении или просто при рассматривании окружающего пространства состоит из серии быстрых скачков с одной точки на другую.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Специальная запись движений глаз позволяет наглядно продемонстрировать, насколько незаметны для нас эти скачки. Чтобы записывать движения глаз, берут крошечное зеркальце и прикрепляют его к контактной линзе сбоку — так, чтобы оно не мешало видеть. После этого на зеркальце направляют пучок света, который, отражаясь, создает на экране небольшое световое пятнышко. Есть еще один способ регистрации движений глаз, более современный; он разработан Д. Робинсоном в Институте Уилмера (университета Джонса Гопкинса). При этой методике по краю контактной линзы на глазу испытуемого монтируется крохотная катушка индуктивности, а самого испытуемого помещают между двумя взаимно перпендикулярными обручами величиной с велосипедное колесо, на которых расположены другие катушки индуктивности. Протекание тока в этих катушках ведет к появлению тока в катушке на контактной линзе. После калибровки такой системы можно регистрировать движения глаз с большой точностью. Для самого бедняги испытуемого ни тот, ни другой способ регистрации движений глаз нельзя назвать приятным!
В 1957 году советский психофизик A. Л. Ярбус записал движения глаз испытуемых при рассматривании различных изображений, в частности таких, как сцена в лесу и женское лицо (рис. 50).
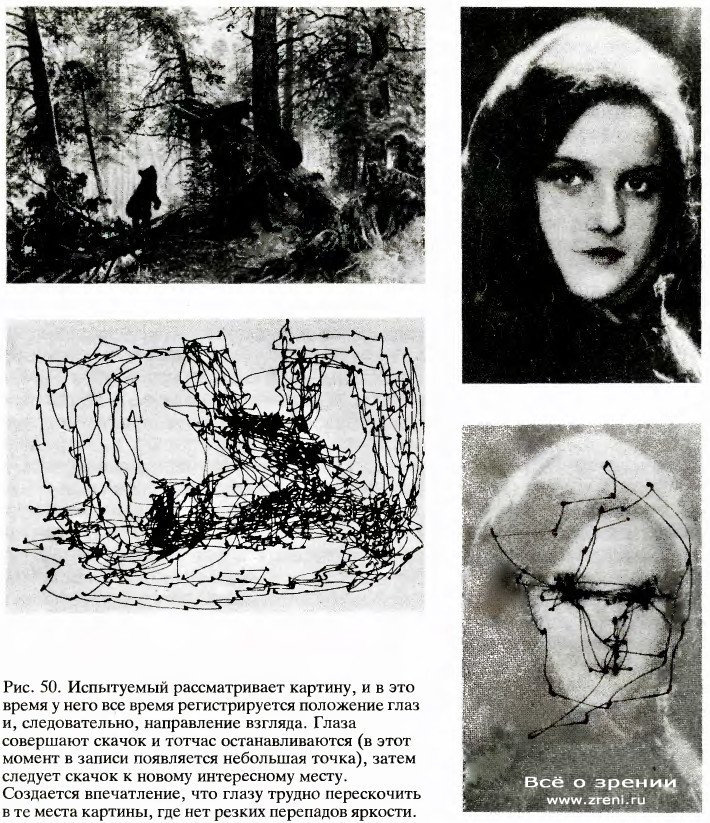
На этих записях периоды фиксации взора представлены точками, а точки соединены линиями, показывающими траекторию движения глаз во время скачков. Даже беглое ознакомление с этими удивительными записями дает много сведений о работе нашей зрительной системы — даже о том, какие предметы и детали окружающего мира нас больше интересуют.
Итак, первый факт, противоречащий нашей интуиции, состоит в том, что при зрительном осмотре глаза перескакивают с одной интересной точки на другую и что нельзя осматривать неподвижную сцену плавными движениями глаз. Задача глазодвигательной системы, видимо, заключается не в том, чтобы удерживать изображение на сетчатках неподвижным, а в том, чтобы предотвращать его плавное смещение. Если же движется вся зрительная сцена, как бывает, когда мы смотрим из окна поезда, то мы прослеживаем эту сцену, фиксируя взором какой-нибудь объект и поддерживая его фиксацию путем плавного движения глаз до тех пор, пока объект не выйдет из зоны видимости, после чего делаем скачок и фиксируем новый объект. Такая последовательность движений глаз — плавное прослеживающее движение, скажем вправо, а затем саккада влево — называется нистагмом. Вы сможете сами увидеть такого рода движения, когда окажетесь в поезде или в трамвае: понаблюдайте за движениями глаз ваших попутчиков, когда они будут рассматривать через окно окружающий пейзаж (будьте только осторожны, чтобы ваше внимание не было неверно истолковано!). Управление скачками глаз при рассматривании интересных элементов сцены с целью перевода их изображений в центральную ямку осуществляется из верхних бугорков четверохолмия. Это показал в 1978 году в серии впечатляющих работ П. Шиллер из Массачусетсского технологического института.
Вторая группа фактов относительно того, как мы видим, еще больше противоречит нашему субъективному впечатлению. Когда мы рассматриваем неподвижную сцену и наш взгляд фиксирует какую-то точку, привлекающую внимание, то эта фиксация не бывает абсолютно неподвижной. Несмотря на все наши попытки жестко фиксировать точку, глаза не остаются в полном покое, а совершают непрерывные микродвижения, называемые микросаккадами. Они совершаются несколько раз в секунду и направлены более или менее случайно, достигая амплитуды 1—2 угловых минут. В 1952 году Л. Риггс и Ф. Рэтлифф из Университета Брауна и Р. Дитчберн и Б. Гинсборг из Редингского университета одновременно и независимо обнаружили, что если изображение на сетчатке искусственно стабилизировать (специальными методами), устранив его смещения относительно сетчатки, то зрительный образ спустя примерно секунду как бы «выцветает» и поле зрения становится совершенно пустым! (Простейший способ стабилизации состоит в том, что точечный источник света прикрепляют к контактной линзе; когда глаза движутся, источник света тоже движется и световое пятно быстро становится невидимым.) Если после стабилизации хотя бы чуть-чуть сместить изображение на сетчатке, световое пятно тотчас появляется снова. Очевидно, микросаккады необходимы для того, чтобы непрерывно видеть неподвижные объекты. Как будто бы Природа, создавая зрительную систему, особенно заботилась о восприятии движения и поэтому постаралась обеспечить нечувствительность клеток к неподвижным объектам, однако потом ей пришлось изобрести микросаккады, для того чтобы сделать и неподвижные объекты видимыми.
Можно предположить, что в этом процессе участвуют сложные клетки коры, особо чувствительные к движению стимула, но, вероятно, не участвуют клетки с дирекциональной избирательностью, так как микросаккады явно случайно распределяются по направлениям. С другой стороны, механизм дирекциональной избирательности должен, по-видимому, быть полезным для выявления движений объектов относительно неподвижного фона, сигнализируя о наличии движения и о его направлении. Для того чтобы следить за движущимся объектом на неподвижном фоне, нужно фиксировать объект и перемещать взор вместе с ним. В этом случае изображение всего остального будет передвигаться по сетчатке (такая ситуация в других случаях встречается редко). Перемещение всех деталей неподвижного фона по сетчатке должно приводить к бурной активности клеток коры.
↑ Концы линий как зрительные стимулы
В стриарной коре встречается еще один вид клеток. Обычно для простых и сложных клеток характерна пространственная суммация — чем длиннее стимульная линия, тем лучше ответ. Однако реакция усиливается лишь до тех пор, пока длина линии не достигнет размеров рецептивного поля: дальнейшее удлинение линии не приводит к более энергичному ответу. В отличие от этого у клеток, реагирующих на концы линий (end stopped cells), удлинение линии до известного предела продолжает улучшать ответ, а если линия выходит за этот предел (в одном или в обоих направлениях), то ответ ослабевает (рис. 51, Б).

Некоторые клетки, которые мы называем «реагирующими исключительно на конец линии» (completely end stopped cells), вообще не отвечают на предъявление стимула в виде длинной линии. Ту зону, с которой можно вызвать ответ клетки, мы называем зоной активации (или возбуждающей зоной), а зоны, расположенные с одного или с двух концов, — зонами торможения (или тормозными зонами). Таким образом, все рецептивное поле подобной клетки состоит из возбуждающей зоны и тормозной зоны (или зон) по краям. Стимул оптимальной ориентации, активирующий клетку с возбуждающей зоны, вызывает максимальное торможение за пределами этой зоны (с одной или с двух сторон). Это можно показать путем повторной стимуляции возбуждающей зоны линией оптимальной длины с оптимальной ориентацией при одновременном тестировании внешней зоны линиями разной ориентации (как показано на рис. 52).

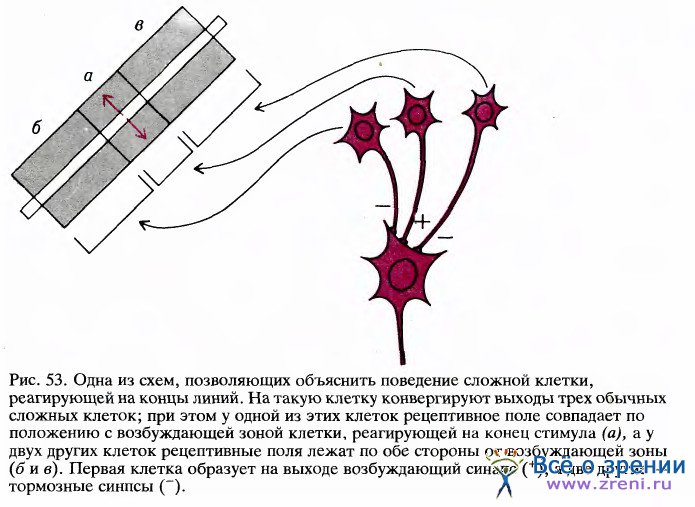
Вначале мы думали, что такие клетки относятся к следующему уровню иерархии корковых нейронов, на уровень выше сложных клеток. Согласно простейшей схеме возможной организации таких клеток, они могли бы иметь один или несколько возбуждающих входов от обычных сложных клеток с рецептивными полями, находящимися в возбуждающей зоне, и тормозные входы от сложных клеток с такой же ориентацией рецептивного поля, расположенных вне зоны возбуждения (эту схему поясняет рис. 53).
Согласно другой возможной схеме (рис. 54), клетка имеет возбуждающий вход от клеток с небольшим рецептивным полем (а) и тормозный вход от клеток с большим рецептивным полем (б).
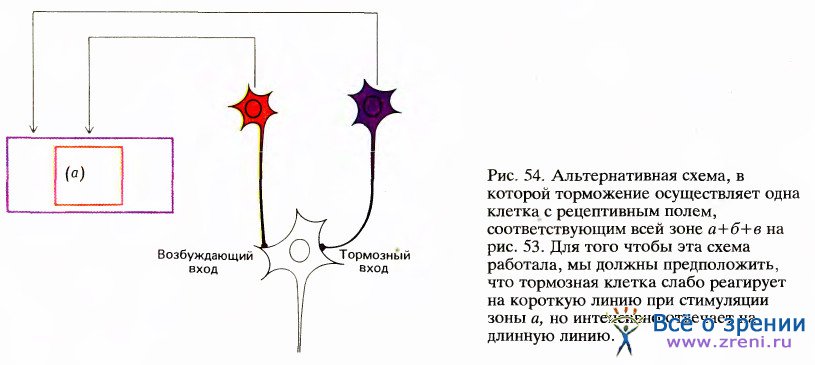
Предполагается, что клетки, подающие тормозные сигналы, максимально чувствительны к длинным линиям, но слабо возбуждаются короткими линиями. Эта вторая схема (аналогичная модели клеток, имеющих рецептивные поля с центром и периферией) — одна из немногих схем, получивших частичное подтверждение. Чарлз Гилберт из Рокфеллеровского университета в Нью-Йорке установил, что сложные клетки в слое 6 стриарной коры обезьяны имеют как раз те свойства, которые нужны для торможения, предполагаемого в данной схеме. Он также показал, что если инактивировать эти клетки путем локальных инъекций, то расположенные в верхних слоях коры клетки, реагирующие на концы линий, теряют способность отвечать торможением на конец линии.
Уже после того как были предложены описанные выше модели, Дж. Генри (Канберра, Австралия) обнаружил простые клетки, реагирующие на конец линии, схема рецептивных полей которых представлена на рис. 55.
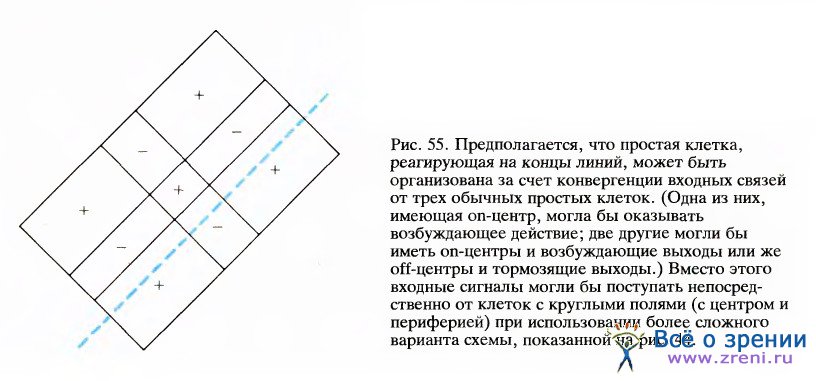
Схема организации подобных клеток аналогична предложенной выше первой схеме, за исключением того, что входы здесь не от сложных клеток, а от простых. Таким образом, ответы сложных клеток, реагирующих на концы линий, могли бы определяться сочетанием возбуждающего входа от одного набора сложных клеток и тормозного входа от другого набора (как в схемах, представленных на рис. 53 и 54) или же конвергенцией входов от нескольких простых клеток, реагирующих на концы линий.
Оптимальным стимулом для клетки, настроенной на конец линии, служит отрезок линии определенной длины. Для такой клетки, которая реагирует на границу, причем с одной стороны отвечает только на ее конец, идеальным стимулом будет угол; для клетки, реагирующей на оба конца светлых или темных линий, наилучшим стимулом окажется короткая белая или черная линия, а также такая кривая линия, которая попадает в возбуждающую зону и не попадает из-за своей кривизны в тормозную зону (так будет, если ориентация ее концов отличается не менее чем на 20—30е от ориентации ее средней части; подобный случай показан на рис. 56). Таким образом, клетки, реагирующие на концы, можно рассматривать как клетки, чувствительные к углам, к кривизне или к резким изломам линий.

↑ Нейрофизиология одиночных клеток и зрительное восприятие
Сам по себе факт, что какая-то клетка мозга реагирует на зрительные стимулы, еще не означает, что она непосредственно участвует в восприятии. Например, многие нейронные структуры в стволе мозга, относящиеся главным образом к зрительной системе, предназначены только для вспомогательных функций — для управления движениями глаз или сужением зрачка, для фокусировки изображения с помощью хрусталика. Что касается тех клеток, которые я описывал в этой главе, то очевидно, что они имеют самое прямое отношение к зрительному восприятию. Как я уже упоминал вначале, разрушение любого небольшого участка стриарной коры приводит к слепоте в каком-то небольшом участке поля зрения. У обезьян повреждение стриарной коры дает такой же эффект. Однако у кошек дело обстоит сложнее — кошка с удаленной стриарной корой может видеть, хотя и не так хорошо, как раньше. Другие отделы мозга, такие как верхние бугорки четверохолмия, возможно, играют у нее более важную роль в зрительном восприятии, чем у приматов. У таких низших позвоночных, как лягушки и черепахи, нет ничего похожего на нашу кору большого мозга, однако никто не станет утверждать, что они слепы.
Теперь мы с достаточной уверенностью можем сказать, что именно любая из описанных выше корковых клеток делает в ответ на стимуляцию изображением видимой сцены. Большинство корковых клеток плохо отвечает на диффузное освещение и хорошо — на линии с нужной ориентацией. Так, при показе фигуры, напоминающей по форме почку (рис. 57), такого рода клетка будет отвечать тогда и только тогда, когда участок границы с определенной ориентацией будет пересекать ее рецептивное поле. Те же клетки, рецептивные поля которых расположены внутри границ фигуры, никак не отреагируют — они будут продолжать давать спонтанный импульсный разряд независимо от присутствия или отсутствия данной фигуры.

Так ведут себя клетки с ориентационной избирательностью. Однако для возбуждения простой клетки недостаточно того, чтобы участок контура соответствовал оптимальной ориентации, — контур должен еще почти точно попадать на границу тормозной и возбуждающей зон рецептивного поля, ведь для ответа нужно, чтобы свет падал на возбуждающую зону, но не распространялся на тормозную. Если хотя бы немного сдвинуть участок контура, не меняя его ориентации, стимуляция данной клетки окажется недостаточной, и теперь уже начнет возбуждаться другая популяция простых клеток. Для сложных клеток условия возбуждения не столь жестки, поскольку популяция клеток, активированная стимулом в какой-то момент времени, не изменится при небольшом сдвиге границ фигуры без изменения их ориентации. Чтобы заметно изменить популяцию возбужденных сложных клеток, нужно достаточно сильно сдвинуть границу — так, чтобы она полностью вышла из рецептивных полей одних клеток и попала в поля других. Таким образом, популяция возбужденных сложных клеток — в отличие от простых — в целом мало меняется при небольших поступательных смещениях объекта.
Если, наконец, обратиться к клеткам, реагирующим на концы линий, то и здесь мы обнаружим менее жесткие ограничения на точное положение стимула (хотя каждая данная конфигурация будет активировать гораздо меньше клеток). Для нейронов этого типа ориентация контура должна всегда совпадать с оптимальной ориентацией возбуждающей зоны рецептивного поля, но должна заметно меняться за пределами этой зоны, чтобы торможение не уравновесило возбуждение. Короче говоря, участок контура должен иметь достаточную кривизну или резко прерываться, чтобы были соблюдены все условия для возбуждения данной клетки (см. рис. 56).
Эти жесткие требования повышают избирательность реакций коры, так как каждый видимый объект возбуждает лишь очень малую долю клеток, на рецептивные поля которых падает его изображение. Такая специализация клеток, вероятно, продолжает возрастать и при дальнейшем переходе к еще более высоким уровням за пределами стриарной коры. На палочки и колбочки воздействует просто свет как таковой. Ганглиозные клетки сетчатки, клетки НКТ и корковые нейроны с центром и периферией сравнивают соответствующие участки поля зрения с окружающим фоном, поэтому они, вероятно, будут реагировать на любой отрезок контура, попавшего в их рецептивное поле, но не будет давать ответа на общее изменение освещенности сетчатки.
Клетки, избирательно чувствительные к ориентации, отмечают не только наличие контура, но и его ориентацию и даже быстроту ее изменения, т. е. кривизну линии. Если такие клетки относятся к категории сложных, то они чувствительны также к движению. Как говорилось в одном из предыдущих разделов книги, возможны два предположения относительно роли чувствительности к движению: возможно, она позволяет легче обращать внимание на движущиеся объекты, а может быть, этот механизм в сочетании с микросаккадами поддерживает реакцию сложных клеток на неподвижные объекты.
Мне кажется, что границы между светлым и темным — самый важный компонент наших зрительных восприятий, но, без сомнения, далеко не единственный. Цвет различных объектов, безусловно, помогает при выделении их (хотя нужно сказать, что наши последние работы указывают на меньшую роль цвета при определении формы). При зрительной оценке формы и глубины используются также распределение светотени на поверхности предметов и текстура их поверхности. Хотя клетки, работу которых мы обсуждали, в принципе могли бы участвовать в восприятии светотеневых переходов и текстуры, не следует ожидать, что они смогут четко реагировать на оба этих признака. Еще предстоит выяснить, каков механизм восприятия текстур в зрительной системе.
Одна из возможных гипотез состоит в том, что сложные клетки обрабатывают информацию о полутонах и текстурах самостоятельно, без помощи каких бы то ни было специализированных групп клеток. Возможно, что такие стимулы не способны сильно активировать большое число клеток, однако текстурам и полутонам свойственна пространственная протяженность, и они могли бы вызывать реакцию сразу многих клеток, хотя каждая клетка будет отвечать умеренно или слабо. Быть может, вялые ответы множества клеток были бы достаточны для передачи информации на более высокие уровни.
Многие люди, в том числе и я, все еще с трудом воспринимают то, что внутренняя часть любой фигуры (например, область внутри почковидного пятна на рис. 57) не является стимулом для клеток нашего мозга и что восприятие однородного внутреннего поля как белого или черного (или окрашенного) зависит только от возбуждения клеток, чувствительных к границам. Логическим доводом в пользу этого служит следующее соображение: если бы при восприятии внутренней области активировались клетки, рецептивные поля которых лежат в ее пределах, то она должна была бы, наоборот, восприниматься как неоднородная!
Таким образом, если мы видим все занятое фигурой пространство как равномерно черное, белое, серое или зеленое, то клетки с рецептивными полями внутри границ фигуры не имеют к этому никакого отношения. С этой мыслью трудно смириться, не так ли? Однако с точки зрения инженера, конструирующего машину для кодирования формы, такой механизм, я думаю, был бы очень подходящим. Единственная информация, которую необходимо в этом случае получать, — это информация о внешнем контуре фигуры; анализ ее внутренности становится излишним. Нужно ли удивляться, что эволюция мозга пошла по пути, позволяющему перерабатывать информацию с помощью минимального количества клеток?
Услышав о свойствах простых и сложных клеток, люди часто делают вывод, что для полного анализа всех мельчайших элементов зрительного поля, включая темные и светлые линии и края различных участков во всех возможных ориентациях, требуется астрономическое число клеток. Разумеется, так оно и есть. Но все дело в том, что кора как раз и содержит астрономическое число клеток. Сегодня мы уже можем сказать, как работают клетки в данном участке мозга, по крайней мере как они отвечают на множество простых зрительных стимулов, встречающихся в повседневной жизни. Я подозреваю, что в стриарной коре нет двух клеток, которые работали бы совершенно одинаково. Действительно, всякий раз, когда с помощью микроэлектрода удавалось одновременно регистрировать активность двух клеток, оказывалось, что эти клетки хоть немного, но отличаются друг от друга — по положению рецептивных полей, дирекциональной чувствительности, интенсивности ответов или каким-то другим параметрам. Короче говоря, складывается впечатление, что в данном отделе мозга если и есть избыточность, то она невелика.
Можем ли мы быть уверены в том, что описанные клетки на самом деле предназначены именно для выявления отрезков прямых линий, а не каких-то других стимулов? Нельзя сказать, что мы и другие исследователи не пытались применять множество других стимулов, включая лица, карту мира и пассы руками. Как показывает опыт, было бы глупо думать, что мы исчерпали все возможности. В начале 1960-х годов, когда мы удовлетворились результатами, полученными с клетками стриарной коры, и решили перейти (и даже фактически уже перешли) к следующей области, нам случайно удалось записать реакцию одной слабо отвечавшей клетки стриарной коры.
Однако, сделав светлую линию более короткой, мы убедились, что эта клетка способна давать весьма энергичную реакцию. Именно тогда мы наткнулись на класс клеток, отвечающих на концы линий. После этого мы еще почти двадцать лет работали с корковыми клетками обезьян, прежде чем обнаружили «пузырьки» — скопления клеток, специфически реагирующих на цвет (они описаны в главе 8). Сделав эти оговорки, я должен добавить, что некоторые из свойств, обнаруженных у клеток стриарной зоны, например ориентационная чувствительность, являются, несомненно, истинными свойствами этих клеток. В пользу этого можно привести много дополнительных данных, например данные по функциональной анатомии.
↑ Бинокулярная конвергенция
До сих пор я почти не упоминал о существовании двух глаз. Очевидный интерес представляет вопрос: получает ли та или иная корковая клетка входные сигналы от обоих глаз, и если да, то одинаковы ли эти входы в количественном и качественном отношении?
Для того чтобы получить ответ, мы должны вернуться на некоторое время к наружному коленчатому телу (НКТ) и выяснить, имеют ли какие-то из его клеток входы от обоих глаз. НКТ — это самый низкий уровень, на котором уже возможно было бы объединение сигналов от двух глаз на одной клетке. Однако возможность эта, по-видимому, здесь не реализована — два разных пучка входных волокон распределяются по разным клеточным слоям, между которыми нет или почти нет взаимодействия. Как можно было бы ожидать, учитывая раздельность проекций от двух глаз, отдельная клетка НКТ должна отвечать на стимуляцию одного какого-то глаза и совсем не реагировать на стимуляцию другого. Судя по результатам некоторых экспериментов, стимулы, подаваемые на «чужой» для данной клетки НКТ глаз, могут оказывать слабое воздействие на реакцию, вызываемую со «своего» глаза. Однако практически можно считать, что каждая клетка находится под контролем только одного глаза.
Интуитивно ясно, что пути, идущие от каждого глаза, должны рано или поздно сойтись вместе, так как мы, глядя на что-либо, видим одну целостную картину. Тем не менее повседневный опыт говорит нам, что если закрыть один глаз, то большого изменения не произойдет — предметы будут казаться такими же четкими, такими же реальными и такими же яркими. Разумеется, общее поле зрения для двух глаз будет несколько шире, поскольку каждый глаз видит на своей стороне большую область пространства, чем другой глаз, хотя эта разница составляет всего лишь около 20—30°. Существенное отличие бинокулярного зрения от монокулярного состоит в ощущении глубины, о чём я Вам расскажу позднее.
В коре обезьяны те клетки, которые получают входные сигналы от НКТ и имеют рецептивные поля с круговой симметрией, сходны с клетками НКТ в том, что они тоже монокулярные. На этом корковом уровне мы находим примерно одинаковое число клеток, возбуждающихся от левого и от правого глаза, по крайней мере в тех участках коры, которые обслуживают область поля зрения в радиусе около 20° от центральной точки фиксации. Однако на следующем уровне коры выявляются уже бинокулярные клетки, простые и сложные, причем у макаков больше половины таких клеток могут реагировать независимо на сигналы от каждого из глаз.
Как только мы нашли бинокулярную клетку, можно тщательно сравнить ее рецептивные поля в обеих сетчатках. Сначала закрываем правый глаз животного и картируем рецептивное поле клетки в левом глазу, отмечая его точное положение на экране или на сетчатке, а также его сложность, ориентацию и расположение возбуждающих и тормозных зон; выясняем также, простая это клетка или сложная, исследуем ее способность реагировать на конец линии и дерекциональную чувствительность; после этого закрываем левый глаз животного, открываем правый и повторяем всю процедуру измерений сначала.
Оказалось, что у большинства бинокулярных клеток все свойства, выявляемые в опытах с левым глазом, обнаруживаются и при стимуляции правого глаза — то же положение на сетчатке, та же дирекциональная чувствительность и т. д. Это позволяет заключить, что все связи, идущие к данной клетке от левого глаза, совпадают по структуре со связями, идущими от правого глаза.
Говоря о таком дублировании связей, нужно сделать одно уточнение. Если, определив для клетки оптимальный стимул, его положение, ориентацию, направление движения и т. д., мы сравним ее ответы при стимуляции одного глаза и при стимуляции другого, интенсивность реакции не всегда окажется одинаковой.
Некоторые клетки действительно одинаково хорошо активируются от обоих глаз, однако другие явно дают более сильный разряд при стимуляции определенного глаза. В целом, за исключением той части корковых клеток, которые обслуживают периферию поля зрения, мы не находим никакого особого преимущества того или другого глаза — в каждом полушарии число клеток, лучше активируемых с противоположной стороны (от контралатерального глаза) и с той же стороны (от ипсилатерального глаза), примерно одинаково. При этом встречаются все степени относительного доминирования глаза, начиная от клеток, возбуждаемых исключительно с левого глаза, и кончая клетками, отвечающими только на стимуляцию правого глаза.
Теперь можно оценить численность различных популяций клеток. Разобьем все изученные клетки, скажем 1000 штук, произвольно на семь классов по относительной эффективности воздействия на них того или другого глаза. Затем подсчитаем число клеток в каждом классе. На рис. 58 представлены соответствующие гистограммы для кошки и макака.
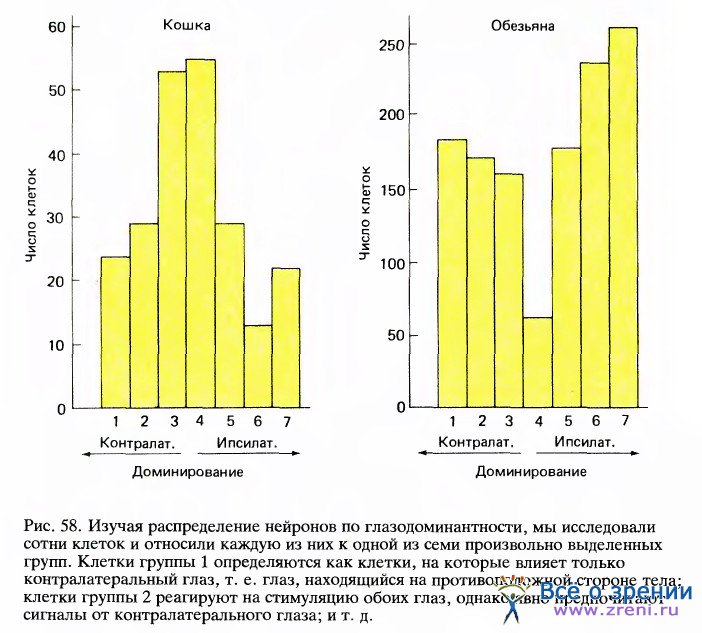
Здесь сразу видны сходства и различия в распределении клеток у этих животных. Видно, что у обоих видов бинокулярные клетки встречаются достаточно часто, причем среди клеток с односторонним доминированием хорошо представлены оба глаза (у макака примерно поровну), что у кошек бинокулярных клеток очень много, что у макаков численность монокулярных и бинокулярных клеток примерно одинакова, причем у бинокулярных клеток зачастую сильно выражено доминирование одного глаза (группы 2 и 5), и что реже всего встречаются клетки, одинаково хорошо активируемые с любого глаза.
Теперь мы можем продвинуться дальше и задать следующий вопрос: отвечают ли бинокулярные клетки при стимуляции обоих глаз лучше, чем при стимуляции одного глаза? Оказывается, многие клетки именно таковы — от одного глаза они активируются слабо или не возбуждаются совсем а от двух глаз дают сильный импульсный разряд, особенно в том случае, когда оба глаза стимулируются одновременно и совершенно одинаково. На рис. 59 показаны записи ответов трех клеток (1, 2 и 3), демонстрирующих выраженную синергию.

Одна из этих клеток вообще не отвечает на стимуляцию только одного глаза, так что мы вообще не смогли бы ее обнаружить, если бы не стимулировали оба глаза одновременно. Эффект синергии у многих клеток выражен слабо или не отмечается вовсе — такие клетки отвечают на стимуляцию обоих глаз примерно так же, как и на стимуляцию каждого глаза по отдельности.
Подобные связи одиночных клеток с двумя глазами еще раз указывают на высокую степень специфичности соединений в мозгу. Мало того, что входные системы связей данной клетки позволяют ей отвечать только на линию определенной ориентации и лишь на одно направление движения, — оказывается к тому же, что эти системы представлены двумя копиями, по одной от каждого глаза. Но и этого еще мало: как мы узнаем из главы 9, большинство связей, по-видимому, должно быть сформировано и готово к работе уже к моменту рождения животного. Все это поистине удивительно.
---
Статья из книги: Глаз, мозг, зрение | Хьюбел Д.

Комментариев 0