Архитектура зрительной коры

Содержание:
Описание
По сравнению с наружными коленчатыми телами (НКТ) и сетчаткой первичная зрительная кора, или стриарная кора, — структура гораздо более сложная. Как мы уже видели, резкое увеличение структурной сложности отделов мозга сопровождается таким же усложнением физиологической организации. В стриарной коре мы находим большее разнообразие функциональных типов клеток.
Нейроны стриарной коры отвечают на более сложные стимулы, т. е. стимулы с большим числом параметров, причем эти параметры должны быть вполне определенными. Если при исследовании клеток сетчатки и НКТ нам достаточно было изменять только местоположение и размеры стимула в виде простого пятна, то теперь мы внезапно столкнулись с необходимостью учитывать такие параметры, как ориентация линии, направление движения, длина линии и ее кривизна, а также выбор глаза, на который подается стимул. Какая связь существует между этими параметрами и структурной организацией коры (если она вообще есть)? Для того чтобы подойти к рассмотрению этого вопроса, сначала необходимо сообщить кое-что о строении стриарной коры.
↑ Анатомия зрительной коры
Кора представляет собой слой нервной ткани толщиной около 2 мм, который почти полностью покрывает большие полушария головного мозга. Площадь поверхности коры у человека составляет больше квадратного фута (около 900 см в квадрате). Общая площадь коры у макаков примерно в 10 раз меньше, чем у человека. Уже более столетия известно, что кора полушарий подразделяется на множество различных корковых полей. Среди всех этих полей первой удалось выделить зону первичной зрительной коры, которая на поперечных срезах выглядит слоистой или полосатой (отсюда и ее давнее название — стриарная кора: лат. stria — полоса). Было время, когда все устремления нейроморфологов сводились к тому, чтобы выявить как можно больше корковых полей на основании подчас весьма тонких гистологических различий. В одной из принятых систем такой классификации стриарной коре был присвоен номер 17. По одной из новейших оценок (Д. ван Эссен, Калифорнийский технологический институт) у макаков первичная зрительная кора занимает около 1200 мм в квадрате (немногим меньше одной трети кредитной карточки); это примерно 15% всей площади коры, т. е. довольно существенная ее часть.
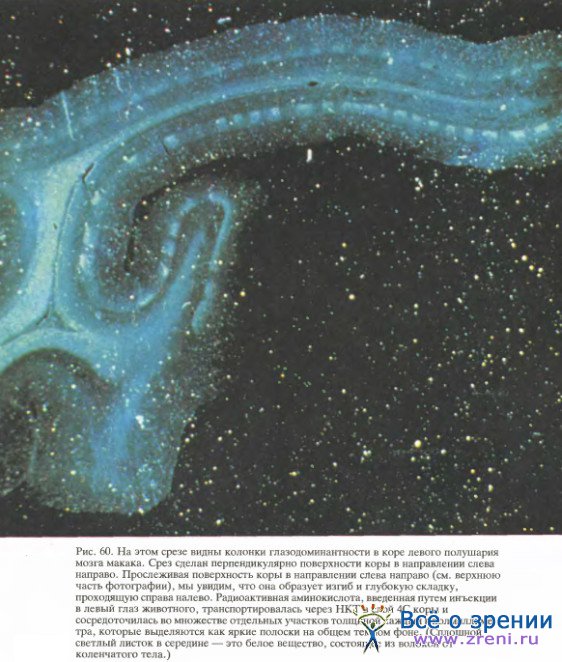
На рис. 62 представлена фотография мозга макака (вид сзади).
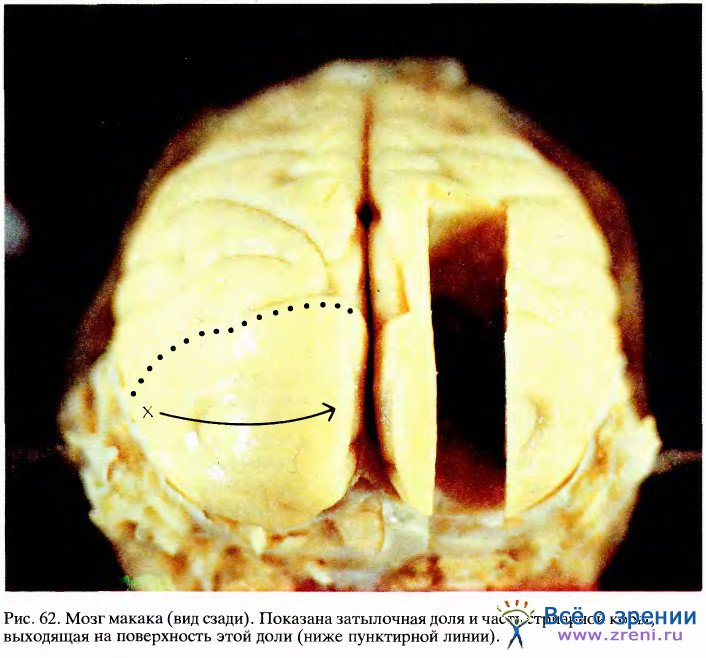
Черепная коробка была удалена, а мозг с целью консервации перфузировали разбавленным раствором формальдегида, что придало ему желтую окраску. Обычно на поверхности коры бросается в глаза сеть кровеносных сосудов, однако здесь они находятся в спавшемся состоянии и поэтому незаметны. На этой фотографии видна главным образом поверхность затылочной доли коры, т. е. та область, которая имеет прямое отношение к зрению и включает не только стриарную кору, но также и одну-две дюжины (а может быть, и больше) престриарных зон. Для того чтобы поместить лоскут нервной ткани толщиной 0,5 мм, равный по площади большой каталожной карточке, в коробку величиной с череп макака, пришлось бы скомкать этот лоскут, наподобие того как мы комкаем лист бумаги, прежде чем выбросить его в мусорную корзину. В результате аналогичного сжимания в коре мозга образуются щели, или борозды, между которыми проходят выпуклые участки, называемые извилинами.
Область на фотографии, лежащая позади (ниже) пунктирной линии, — это открытая, видимая часть стриарной коры. Хотя стриарная кора занимает большую часть поверхности затылочной доли, на фото мы можем увидеть только часть ее (0,3 — 0,5 всей площади); остальная часть скрыта в борозде.
Стриарная кора (поле 17) имеет множество выходных волокон, значительная часть которых идет в следующую корковую область — зрительное поле 2, часто также называемое полем 18, так как оно примыкает к полю 17. Поле 18 представляет собой полоску коры шириной примерно 6—8 мм, почти полностью окружающую поле 17. На фото (рис. 62) можно видеть только часть поля 18, расположенную над пунктирной линией, которая соответствует границе между полями 17 и 18; большая часть поля 18 уходит вниз, в глубокую борозду, расположенную как раз перед пунктирной линией. Поле 17 упорядоченно, точка в точку, проецируется на поле 18, а последнее в свою очередь образует проекции по крайней мере в трех затылочных зонах величиной каждая с почтовый штемпель.
Эти зоны известны как поле МТ (сокращение от medialis temporalis, т. е. срединное височное) и зрительные поля 3 и 4 (часто обозначаемые как V3 и V4). Как правило, каждая корковая зона имеет проекции в нескольких других зонах, расположенных на более высоком уровне. Кроме того, для каждой зоны коры можно найти обратную проекцию — в ту зону или те зоны, откуда идут входные волокна. И как будто стремясь к еще большей сложности, каждая из этих зон образует проекции в глубинных структурах мозга, например в верхних бугорках четверохолмия и в различных участках таламуса (сложного клеточного образования величиной с мяч для гольфа, небольшую часть которого составляет НКТ). И все эти зрительные зоны получают входные сигналы от разных участков таламуса — так же, как НКТ проецируется в первичную зрительную кору, так различные области таламуса образуют проекции в других корковых зонах.
На той же фотографии буквой X обозначен тот участок поля 17, который получает входные сигналы от центральных ямок обеих сетчаток. Если двигаться по левому полушарию из точки X в направлении, указанном стрелкой, то в поле зрения это будет соответствовать движению объекта по горизонтали от точки фиксации. Если же, выйдя снова из точки X, двигаться прямо вдоль границы между полями 17 и 18, то это будет соответствовать движению объекта от точки фиксации вниз, а перемещение в обратном направлении — движению объекта вверх. Самой стрелке в поле зрения соответствует горизонтальная линия длиной около 6°. Области поля зрения, расположенные дальше 9° от центра взора, отображаются в ту часть поля 17, которая лежит в складке коркового слоя под наружной поверхностью мозга, параллельно ей.
Для того чтобы посмотреть, как выглядит кора в поперечном разрезе, мы вырезали участок зрительной коры из правой половины мозга, изображенного на рис. 62. Полученный срез был обработан крезиловым фиолетовым, который окрашивает тела нейронов в синий цвет, но не окрашивает аксоны и дендриты. На этой микрофотографии, сделанной при малом увеличении, нельзя различить отдельные клетки, но можно видеть темные слои плотно упакованных клеток и более светлые слои, где клеток гораздо меньше. Под наружной поверхностью коры видна сложная структура из складок, напоминающая по форме гриб. В действительности эта структура и поверхностная часть коры без перерыва переходят друг в друга. Светлоокрашенное пространство — это белое вещество, оно заполняет области между наружными частями коры и ее глубже расположенными складками. Белое вещество состоит в основном из миелинизированных нервных волокон, которые не окрашиваются красителем. Часть коры, содержащая тела нервных клеток, их аксоны и дендриты, а также синапсы, образует серое вещество.
По сложности своей слоистой структуры поле 17 превосходит все другие зоны коры. Эту разницу можно заметить даже при малом увеличении (рис. 63), если сравнить поле 17 и соседнюю область справа — поле 18 (граница между ними отмечена буквой г).

Еще одна деталь: если сравнить на рис. 63 области а, б и в, куда проецируются разные участки поля зрения (расстояния их от точки фиксации < 5, 6 и 80—90 градусов соответственно), мы не заметим существенного различия ни в толщине коры, ни в ее слоистой структуре. Как выяснилось, такая однородность имеет важное значение. К этому вопросу я еще вернусь в этой главе.
↑ Слои зрительной коры
На рис. 64 представлен небольшой участок поля 17 при более сильном увеличении.
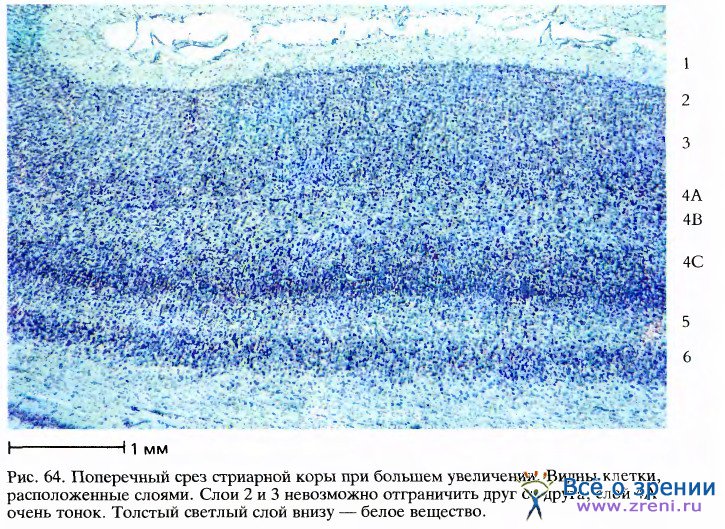
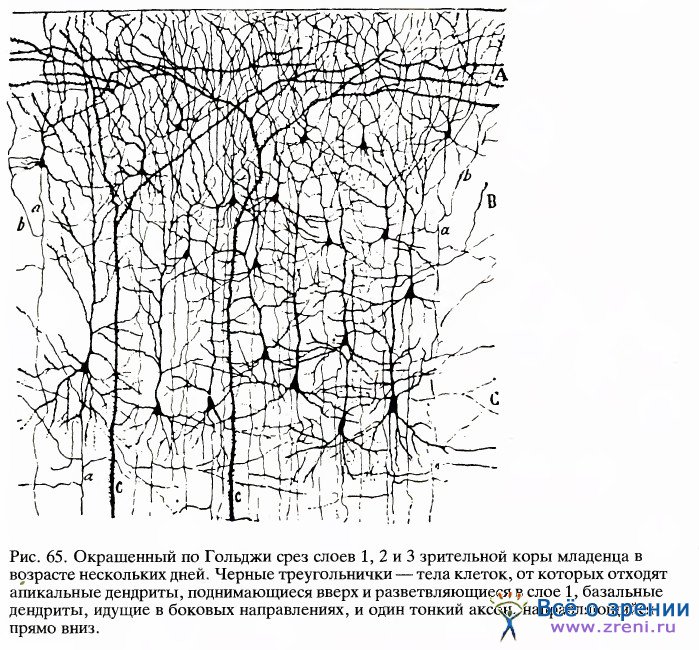
Здесь уже можно различить в виде точек тела отдельных клеток и получить некоторое представление об их размерах, количестве и распределении. Видимая картина слоев частично определяется интенсивностью окрашивания клеток и частично—плотностью их упаковки. Слои 4С и 6 - самые плотные и темные. В слоях 1, 4В и 5 клетки расположены наиболее рыхло. Слой 1 едва ли содержит хотя бы одну нервную клетку, но здесь множество аксонов, дендритов и синапсов. Для того чтобы показать, что разные слои включают разные типы клеток, нужно применить какой-либо вариант методики, изобретенной в 1900 году К.Гольджи. При окраске по Гольджи выявляются лишь немногие клетки, зато на срезах клетка видна вся целиком, включая ее аксон и дендриты. Два главных класса корковых клеток — это пирамидные клетки, имеющиеся во всех слоях коры, кроме слоев 1 и 4, и звездчатые клетки, которые есть во всех слоях. Примеры пирамидной и звездчатой клеток представлены на рис. 4. Типичное распределение пирамидных клеток в коре показано на рисунке Рамон-и-Кахала, помещенном в его книге «Гистология» (рис. 65); здесь изображена лишь небольшая доля всех пирамидных клеток, имеющихся в этом участке, наверное около 1 процента.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
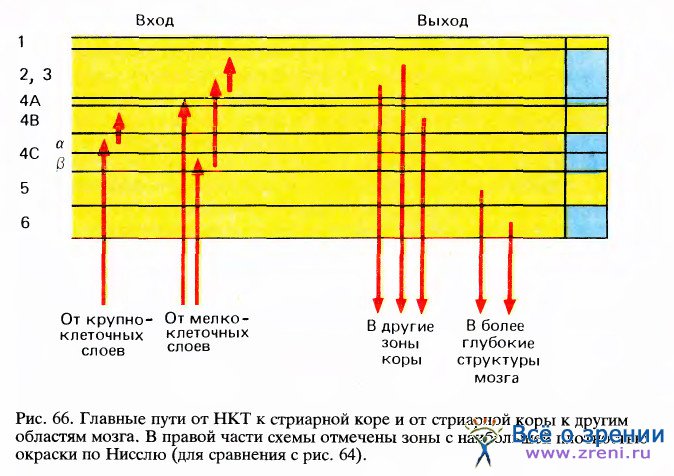
Волокна, идущие из НКТ, приходят в кору из белого вещества мозга. Пройдя в диагональном направлении, большая часть волокон доходит до слоя 4С, многократно разветвляется и, наконец, достигает конечного пункта, образуя синапсы со звездчатыми клетками, которые в основном заполняют этот слой. Аксоны, выходящие их двух вентральных (крупноклеточных) слоев НКТ, оканчиваются в верхней половине слоя 4С, называемой подслоем 4Са, a те, которые выходят из четырех дорсальных (мелкоклеточных) слоев НКТ, оканчиваются в нижней половине слоя 4С (подслой 4Сb). Как видно из схемы на рис. 66, эти два подслоя образуют разные проекции в верхних слоях — 4Са посылает сигналы в 4В, а 4Сb — в слои 2 и 3. Последние в свою очередь тоже различаются по своим проекциям. Заметное различие в нервных путях, выходящих из двух групп слоев НКТ, — это одно из многих оснований предполагать, что с этими путями связаны две разные системы. У большинства пирамидных клеток в слоях 2, 3, 4В, 5 и 6 аксоны выходят из коры, однако у этих аксонов есть ответвления (коллатерали), которые образуют локальные связи и таким образом помогают распределить информацию по всей толще коры.
Корковые слои различаются не только своими входами и внутренними связями, но также и адресами тех структур, куда они проецируются. Все они, кроме слоев 1, 4А и 4С, имеют выходные волокна, уходящие за пределы коры. Верхние слои 2 и 3, а также слой 4В посылают сигналы главным образом в другие области коры, тогда как нижние слои проецируются на подкорковые структуры: из слоя 5 выходные волокна идут в средний мозг, в верхние бугорки четверохолмия, а из слоя 6 волокна направляются обратно в НКТ. Хотя уже почти столетие известен тот факт, что выходные волокна НКТ идут в основном в корковый слой 4, мы ничего не знали о различиях между выходами разных слоев коры, пора в 1969 году японский исследователь К. Тояма не обнаружил впервые эти различия в физиологических экспериментах. С тех пор эти данные были многократно подтверждены морфологическими методами.
Рамон-и-Кахал был первым, кто понял, насколько коротки внутренние связи в коре. Как уже говорилось, наиболее многочисленные связи идут вверх и вниз, тесно объединяя разные слои. Диагональные и боковые (горизонтальные) связи обычно имеют длину от 1 до 2 мм, хотя некоторые связи прослеживаются на расстояние 4—5 мм.
Такое ограничение «горизонтальной» передачи информации в коре приводит к важным последствиям. Если входы организованы по топографическому принципу (в случае зрительной системы — в соответствии с положением отображаемой точки на сетчатке или в поле зрения), то этот же принцип должен сохраняться и в организации выходов. Какие бы функции ни выполняла кора, проводимый анализ должен быть локальным. Информация о любом небольшом участке поля зрения поступает в какой-то небольшой участок коры, где подвергается преобразованию, анализу, переработке (назовите это как угодно) и передается куда-нибудь для дальнейшей обработки независимо от того, что происходит с информацией о соседнем участке. Таким образом, изображение зрительной сцены анализируется как бы мозаично. Поэтому первичная зрительная кора не может быть тем местом, где распознаются, воспринимаются или подвергаются другой обработке целые объекты, например лодки, шляпы, лица, т. е. не может быть субстратом процессов «восприятия». Разумеется, столь общий вывод вряд ли правомерно делать на основе одних только анатомических данных: ведь информация могла бы передаваться на большие расстояния вдоль коры и по принципу эстафеты, с отдельными этапами по одному миллиметру.
Однако, регистрируя активность клеток при одновременной стимуляции сетчатки, можно показать, что дело обстоит не так: выясняется, что все клетки в данном небольшом участке имеют маленькие рецептивные поля и что соседние клетки всегда имеют рецептивные поля, расположенные почти в одном и том же месте. Нет никаких физиологических данных, которые указывали бы на то, что какая-либо клетка в первичной зрительной коре обезьяны соединена с какой-то другой клеткой на расстоянии больше 2—3 миллиметров.
На «мозаичный» характер обработки информации давно уже указывают и данные клинической неврологии. После небольших инсультов, опухолей или ранений первичной зрительной коры может наступать полная слепота в небольших, четко ограниченных участках поля зрения. При этом во всех остальных участках сохраняется нормальное зрение. Если бы каждая клетка была в какой-то мере связана со всеми другими, то вместо этого можно было бы ожидать некоторого общего, нелокального ухудшения зрения. Отойдя несколько в сторону от нашей темы, следует заметить, что больные с такими поражениями могут и не осознавать наличия у них дефекта поля зрения, особенно в тех случаях, когда не затронута та часть коры, где находится проекция центральной ямки сетчатки, т. е. точки фиксации взора. Во всяком случае, такие больные не видят в поле зрения ни сплошь черных или сплошь серых островков, ни каких-либо других дефектов. Даже если разрушена вся затылочная доля на одной стороне и человек ничего не видит в противоположной половине поля зрения, то у него не возникает впечатления, что с этой стороны зрительный мир от него закрыт. У меня самого иногда бывают приступы мигрени (к счастью, без головной боли) с кратковременной слепотой, нередко в значительной части одного из полуполей зрения. Если спросить, что именно я в этой области вижу, я могу только сказать, что не вижу буквально ничего — ни белого, ни серого, ни черного, просто ничего.
Еще одна любопытная особенность, наблюдаемая у больных с островком локальной слепоты, или скотомой, — это феномен, известный под названием «заполнения». Если человек со скотомой будет смотреть на линию, проходящую через дефектный участок поля зрения, он не заметит никакого разрыва — линия кажется совершенно непрерывной. Можно продемонстрировать аналогичное явление, воспользовавшись тем, что и в нормальном глазу имеется слепое пятно. Это то место, где из глаза выходит зрительный нерв. Слепое пятно представляет собой овальный участок величиной около двух миллиметров, где нет ни палочек, ни колбочек. Отыскать слепое пятно можно с помощью небольшого бумажного диска — метод настолько прост, что доступен каждому. Сначала закройте один глаз, например левый. Держа его закрытым, фиксируйте другим глазом какой-нибудь маленький предмет в комнате. Теперь возьмите в руку бумажный диск и отведите руку на всю длину, поместив ее точно на линии, соединяющей глаз с предметом. Затем медленно сдвигайте руку вправо точно по горизонтали (наличие темного фона облегчает эксперимент). Бумажный диск исчезнет примерно в тот момент, когда смещение его достигнет 18°. А теперь если взять какой-нибудь стержень и поместить его так, чтобы он пересекал слепое пятно, то он будет выглядеть как целостный объект, не имеющий никаких разрывов. Область, занимаемая слепым пятном, аналогична скотоме — вы не заметите ее существования до тех пор, пока не проделаете описанный выше опыт. Вы не видите здесь ни черного, ни серого, ни чего-либо иного, вы просто ничего не видите.
Сходным образом, если смотреть на большой лист белой бумаги, то в коре будут возбуждаться только те клетки, рецептивные поля которых попадают на край этого листа (поскольку корковые клетки имеют склонность игнорировать равномерно распределенный свет). В этом случае прекращение активности клеток, рецептивные поля которых целиком лежат в пределах листа бумаги, должно приводить к исчезновению различий яркости в этой области поля зрения. Островок слепоты в данном случае не должен быть виден — и он не виден. Мы не можем увидеть свое слепое пятно в виде черного отверстия в листе белой бумаги, на которую смотрим. Феномен заполнения, проявляющийся в опыте с листом бумаги, должен убедить каждого в том, что, исходя из одних только интуитивных представлений, мы не сможем понять, как работает на самом деле мозг.
↑ Архитектура коры
Теперь мы можем вернуться к исходному вопросу: как связаны между собой физиологические свойства корковых клеток и их структурная организация? Можно еще больше заострить этот вопрос: зная, что корковые клетки могут различаться по положению их рецептивных полей, «сложности», предпочитаемой ориентации стимула, глазодоминантности, по оптимальному направлению движения стимула и оптимальной длине стимульной линии, правомерно ли ожидать, что соседние клетки сходны по некоторым из этих параметров или даже по всем? Или же клетки с разными свойствами просто рассыпаны по всей коре случайным образом, без всякой связи с их физиологическими свойствами?
Если изучать анатомию коры невооруженным глазом или под микроскопом, это мало что даст. На поперечных срезах можно увидеть отчетливые различия между отдельными слоями коры. Однако если прослеживать на поперечном срезе тот же слой по длине или же изучать срезы одного слоя, сделанные параллельно границе слоев, то мы увидим один лишь серый однородный материал. Хотя такая однородность может указывать на случайное распределение клеток, мы знаем, что по крайней мере в отношении одной переменной клетки расположены весьма упорядоченно. Речь идет о закономерном соответствии между распределением клеток в стриарной коре и положением их рецептивных полей на сетчатке, т. е. о том, что соседние клетки коры должны иметь рецептивные поля, расположенные близко друг к другу в поле зрения.
Именно такая картина выявляется в экспериментах. У двух клеток, лежащих рядом в коре, рецептивные поля обычно даже перекрываются на большей части своей площади. Тем не менее эти поля не накладываются точно друг на друга. Если сдвигать микроэлектрод вдоль коры от клетки к клетке, то положения соответствующих рецептивных полей сдвигаются в направлении, которое можно предсказать, зная топографию отображения сетчатки в коре. Никто не усомнился бы в таком результате и 50 лет назад, имея данные о связях НКТ с корой и о случаях локальной слепоты после инсультов. Но как обстоит дело с остальными параметрами — глазодоминантностью, «сложностью», ориентацией адекватных стимулов и другими?
Потребовалось несколько лет для того, чтобы научиться достаточно надежно стимулировать корковые клетки и регистрировать их ответы; в результате появилась возможность описывать реакции не только отдельных клеток, но и сравнительно больших групп нейронов. Началось с того, что нам случайно удалось одновременно записать ответы двух или нескольких клеток (пример такой записи был приведен на рис. 59). Записать ответ двух соседних клеток несложно. В тех экспериментах, где мы определяли предпочитаемый клеткой стимул, мы почти всегда использовали внеклеточное отведение, помещая кончик микроэлектрода рядом с клеткой; в этом случае регистрировались не изменения мембранного потенциала, а токи, связанные с импульсами. Часто при этом оказывалось, что мы регистрируем реакции не одной, а нескольких клеток одновременно — скажем, в том случае, когда кончик микроэлектрода останавливался на полпути между двумя телами нейронов. Импульсные разряды одиночных клеток при таком отведении почти идентичны, однако величина и форма импульсов зависят от расстояния и от взаимного расположения клеток, так что разряды, отводимые одновременно от двух клеток, обычно оказываются разными, и поэтому их можно легко различить. Выполняя такого рода отведения от двух клеток, мы смогли отчетливо увидеть, чем различаются соседние клетки и в чем они одинаковы.
В одном из первых таких отведений были обнаружены две корковые клетки, отвечавшие на противоположные движения руки, которой махали перед животным. В этом случае две лежавшие бок о бок клетки давали разные (по существу, противоположные) реакции на движение стимула. Однако в других отношениях эти клетки почти наверное проявляли сходные свойства. Если бы я в 1956 году был уже подготовлен к изучению ориентационной избирательности, то я, весьма вероятно, обнаружил бы, что обе предпочитаемые ориентации близки к вертикальной, так как данные клетки хорошо отвечали на горизонтальные движения стимула. Тот факт, что обе они реагировали на возвратно-поступательное движение руки, означает, что положения рецептивных полей этих клеток приблизительно совпадали. Если бы я исследовал эти нейроны на глазодоминантность, то скорее всего этот параметр тоже оказался бы одинаковым.
Уже в первых записях активности корковых нейронов нас поразило то, как часто две клетки, реакции которых можно регистрировать одновременно, одинаковы по глазодоминантности, сложности и, что самое удивительное, по оптимальной ориентации стимулов. Такие совпадения, вряд ли случайные, позволяют предположить, что клетки с одними и теми же свойствами объединены в группы. Возможность такой группировки весьма заинтересовала нас, и как только это предположение подтвердилось, мы стали выяснять, каковы размеры и форма этих групп.
↑ Исследование коры
С помощью микроэлектродов можно исследовать только отдельные точки коры. Для того чтобы получить представление о трехмерной организации мозга, приходится медленно погружать электрод в глубину, время от времени останавливать его для записи активности какой-нибудь клетки (а возможно, — двух или трех клеток), отмечать по специальной шкале показания глубины, а затем повторять все сначала. Рано или поздно кончик микроэлектрода пройдет через весь корковый слой, и тогда электрод можно вынуть и снова ввести его в каком-нибудь другом месте. После окончания эксперимента делают срез, окрашивают его и исследуют под микроскопом с целью определить положение каждой из нервных клеток, активность которых регистрировалась. В одном эксперименте длительностью около 24 часов обычно удается сделать две-три проходки примерно по 4—5 миллиметров каждая. За одну проходку можно наблюдать ответы примерно 200 клеток.
Микроэлектрод настолько тонок, что едва удается найти след от его прохождения под микроскопом, поэтому нет оснований думать, что при проходке микроэлектродом будет повреждено много клеток и это может повлиять реакции близлежащих нейронов. Вначале нам было трудно находить на гистологическом срезе след от микроэлектрода, не говоря уже о том, чтобы определить положение кончика электрода, и это затрудняло оценку положения тех клеток, активность которых была записана. Эту трудность удалось преодолеть, когда было обнаружено, что пропускание через микроэлектрод слабого тока приводит к разрушению клеток в ближайшей окрестности кончика микроэлектрода, и на гистологических срезах эта зона разрушения хорошо видна. К счастью, при пропускании тока сам микроэлектрод не повреждается. Поэтому за одну проходку ток пропускают 3—4 раза, отмечая при этом глубину погружения микроэлектрода, а так как глубину отмечают и при регистрации активности клеток, то можно оценить и положение каждой из них. Разумеется, при воздействии тока погибает несколько клеток около кончика микроэлектрода, однако их не так много, чтобы могла быть нарушена работа более удаленных нейронов. Для того чтобы при этом не исказить ответы клеток, лежащих впереди на пути микроэлектрода, его кончик немного продвигают вперед, регистрируют активность клеток, а затем кончик отводят назад и тогда уже пропускают ток.
↑ Вариации в сложности
Как и следовало ожидать, клетки во входном корковом слое — слое 4 — проявляют более простое поведение, чем клетки на выходе. Мы уже отмечали в этой главе, что у обезьяны клетки в слое 4b3, куда приходят волокна из четырех верхних (мелкоклеточных) слоев НКТ, по-видимому, не обладают избирательностью к ориентации стимула и ведут себя подобно клеткам, имеющим круглые рецептивные поля с центром и периферией. В слое 4Са, имеющем входы от двух вентральных (крупноклеточных) слоев НКТ, некоторые клетки обладают круглыми полями с центром и периферией, а всем остальным, видимо, свойственны простые рецептивные поля и ориентационная чувствительность. Если перейти к следующему уровню — к слоям, лежащим выше и ниже слоя 4С, — то подавляющее большинство клеток окажутся сложными. Клетки, реагирующие на концы линий, в слоях 2 и 3 составляют около 20 процентов, а в других слоях встречаются редко. Таким образом, в целом обнаруживается явная корреляция между сложностью клеток и положением их в зрительном пути, которое можно оценить по числу синаптических переключений до данного места.
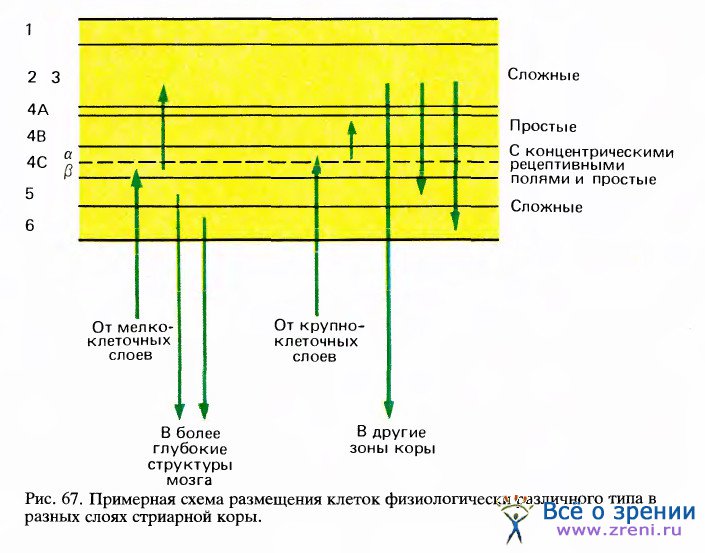
В утверждении, что большинство клеток выше и ниже слоя 4 — сложные, упускаются из виду существенные различия между слоями: сложные клетки здесь далеко не одинаковы. Разумеется, все они имеют одно общее свойство, характерное для сложных клеток, — на движущуюся линию оптимальной ориентации они отвечают по всему рецептивному полю, независимо от конкретного положения стимула. Однако они различаются между собой другими свойствами. Можно выделить четыре подтипа клеток, которые содержатся в основном в разных слоях. Большинство сложных клеток в слоях 2 и 3 отвечает тем лучше, чем длиннее стимульная линия (в этом проявляется свойство суммации по длине). Однако реакция таких клеток становится слабее, когда длина стимула превышает некоторую критическую величину (если данная сложная клетка относится к нейронам, реагирующим на концы линий). Что касается клеток слоя 5, то для них короткие линии, занимающие по длине лишь небольшую долю рецептивного поля, почти так же эффективны, как и длинные; рецептивные поля этих клеток гораздо больше, чем у клеток в слоях 2 и 3. Напротив, клетки слоя 6 реагируют тем лучше, чем длиннее линия с оптимальной ориентацией (их ответ начинает ухудшаться только тогда, когда линия займет всю длину рецептивного поля, которая в несколько раз больше его ширины, т. е. рецептивное поле длинное и узкое). Можно сделать вывод, что аксоны, выходящие из слоев 5 и 6 и из слоев 2 и 3 и идущие к разным участкам мозга (к верхним бугоркам четверохолмия, к НКТ, к другим зрительным полям), должны служить для передачи разных видов зрительной информации.
Подводя итоги, можно сказать, что при переходе от слоя к слою в поведении клеток выявляются более важные различия, чем, скажем, различия в оптимальной ориентации стимула или же в характере глазодоминантности. Наиболее заметные различия между клетками разных корковых слоев касаются сложности их реакций; это отражает тот простой анатомический факт, что одни слои расположены ближе к входу в кору, чем другие.
↑ Колонки глазодоминантности
Группы клеток стриарной коры с различной глазодоминантностью были обнаружены раньше всего, так как они довольно крупные. Разработано много методов исследования таких группировок, поэтому они сейчас лучше всего изучены. Уже в самом начале исследований на коре обезьян стало очевидно, что всякий раз, когда микроэлектрод входит в кору перпендикулярно ее поверхности, он встречает одну за другой клетки, лучше реагирующие на стимуляцию одного и того же глаза (это показано на рис. 68).
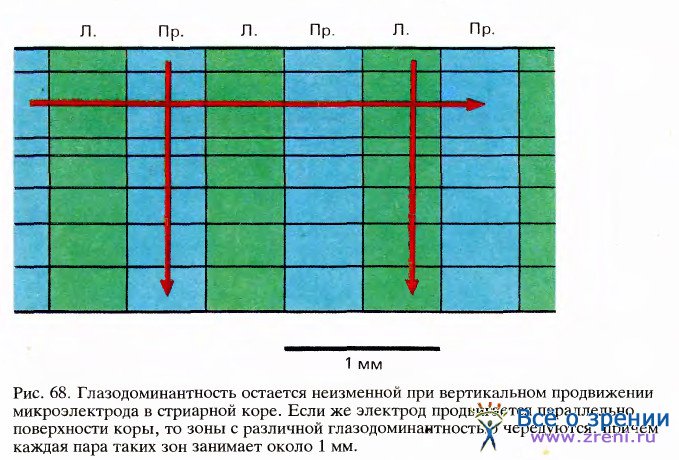
Если микроэлектрод вынуть и ввести его в другом месте в нескольких миллиметрах от предыдущего, то снова для всех встречающихся клеток доминирующим будет один глаз — тот же, что и раньше, или другой. В слое 4С, имеющем входы непосредственно от НКТ, доминирование одного глаза становится уже не относительным, а абсолютным, монопольным. В тех же слоях, которые расположены выше и ниже и, следовательно, находятся дальше в цепи синаптических переключений, более половины всех клеток могут возбуждаться и от недоминирующего глаза. Такие клетки мы называем бинокулярными.
Если вместо того, чтобы перемещать электрод перпендикулярно поверхности коры, мы вводили его с наклоном, как можно ближе к линии, параллельной поверхности, то клетки с разной глазодоминантностью чередовались — доминирующим был то один глаз, то другой. Полный цикл этой смены соответствовал примерно одному миллиметру. Очевидно, если бы можно было видеть структуру коры сверху, она представлялась бы мозаикой, составленной из чередующихся зон с доминированием правого и левого глаза.
Чем обусловлено такое чередование, стало ясно, когда был разработан новый метод окраски, позволяющий прослеживать ветвление отдельных аксонов, приходящих из НКТ, и распределение их окончаний в коре. Ветвление одного аксона таково, что тысячи его концевых синапсов образуют в слое 4С два или три скопления шириной 0,5 мм, разделенных промежутками около 0,5 мм, как показано на рис. 69.

Поскольку клетки НКТ монокулярные, каждый отдельный аксон связан в конечном счете либо с правым, либо с левым глазом. Предположим, что аксон, изображенный на рис. 69 слева, относится к волокнам от левого глаза; в этом случае каждое входящее в тот же участок коры волокно, связанное с левым глазом, будет разветвляться в тех же скоплениях окончаний величиной 0,5 мм каждое. Промежутки между этими скоплениями размером тоже 0,5 мм будут заняты концевыми разветвлениями волокон от правого глаза. Такой особый тип распределения аксонов из НКТ в слое 4С сразу позволяет объяснить строгую монокулярность клеток в этом слое.
Для того чтобы избирательно окрашивать одно и только одно волокно, требовался новый метод, и он был предложен в конце 1970-х годов. Этот метод основан на использовании аксонного транспорта — процесса, в результате которого различные материалы (белки или даже более крупные частицы) непрерывно переносятся в обоих направлениях внутри аксона. Некоторые из них транспортируются со скоростью нескольких сантиметров в час, другие — несколько миллиметров в сутки. Для того чтобы окрасить одиночный аксон, с помощью микропипетки вводят в него вещество, о котором известно, что оно переносится по аксону и окрашивает его, но не влияет на структуру клетки. В настоящее время чаще всего используют фермент пероксид азу из хрена. Этот фермент распространяется по аксону в обоих направлениях и катализирует химическую реакцию, продукт которой очень эффективно окрашивает клетку. Поскольку фермент служит катализатором, достаточно ничтожных его количеств, чтобы получить интенсивную окраску. Важно и то, что в самой нервной ткани фермента с подобными свойствами нет: это исключает возможность нежелательной окраски фона.
Тот факт, что кора разбита на колонки глазодоминантности, проходящие от поверхности мозга вплоть до белого вещества, подтвердил морфологические данные о том, что группы клеток в слое 4С служат главными местами передачи зрительной информации в клеточные слои, лежащие выше и ниже этого слоя. Наличие некоторого числа горизонтальных и диагональных связей длиной около миллиметра, идущих по всем направлениям, должно приводить к некоторому размыванию четких зон доминирования правого или левого глаза в слоях, расположенных выше и ниже слоя 4С (рис. 70).

Поэтому можно ожидать, что клетка, расположенная прямо над центром зоны доминирования левого глаза в слое 4, будет явно лучше отвечать на стимуляцию этого глаза (а возможно, и полностью контролироваться им), тогда как клетка, расположенная ближе к границе между участками разной глазодоминантности, может оказаться бинокулярнои без какого-либо доминирования одного из глаз. Действительно, при горизонтальном передвижении электрода в верхнем слое коры или же в слое 5 или 6 отмечаются постепенные изменения глазодоминантности: сначала будут встречаться клетки, намного сильнее активируемые одним глазом, затем клетки, у которых эта асимметрия выражена слабее, потом клетки с равной зависимостью от обоих глаз, после чего начнет усиливаться влияние другого глаза. Такие плавные переходы резко контрастируют с той внезапной сменой глазодоминантности, которую мы наблюдаем при движении электрода вдоль слоя 4С (рис. 71).

Если смотреть «сбоку», две совокупности клеток в слое 4С выглядят как чередующиеся участки. Однако нам хотелось узнать, как будет выглядеть распределение этих участков при взгляде на поверхность коры сверху. Допустим, что они отмечены на поверхности коры черным или белым цветом. С точки зрения топологии можно распределить эти участки несколькими разными способами: в шахматном порядке, в виде чередующихся черных и белых полос, в виде черных пятен на белом фоне или использовать любые комбинации этих паттернов. На рис. 72 показаны три возможных варианта такого распределения.

Решать поставленную задачу с помощью одних только микроэлектродов означает использовать одномерные средства для расшифровки неизвестной трехмерной структуры. Это задача безнадежная — все равно что пытаться подстричь траву на лужайке с помощью ножниц для ногтей. Столкнувшись с такими трудностями, многие предпочли бы переключиться на совершенно иной вид деятельности — заняться, скажем, сельским хозяйством или юриспруденцией. (В начале 1960-х годов, когда Торстен Визел и я были более терпеливыми и решительными, чем сейчас, мы действительно попытались выяснить топографию глазодоминантности с помощью микроэлектродов, и нам удалось даже получить кое-какие результаты. Образно выражаясь, я в те дни действительно пытался подстричь травяной газон ножницами, так как мы не могли позволить себе купить газонокосилку. В то время мы были беднее, чем нынешние аспиранты, однако были, вероятно, более настойчивыми.)
К счастью, в последние десять лет были придуманы очень эффективные методы нейроморфологических исследований, и сейчас поставленная нами задача уже решена независимо несколькими способами; два из них описаны ниже.
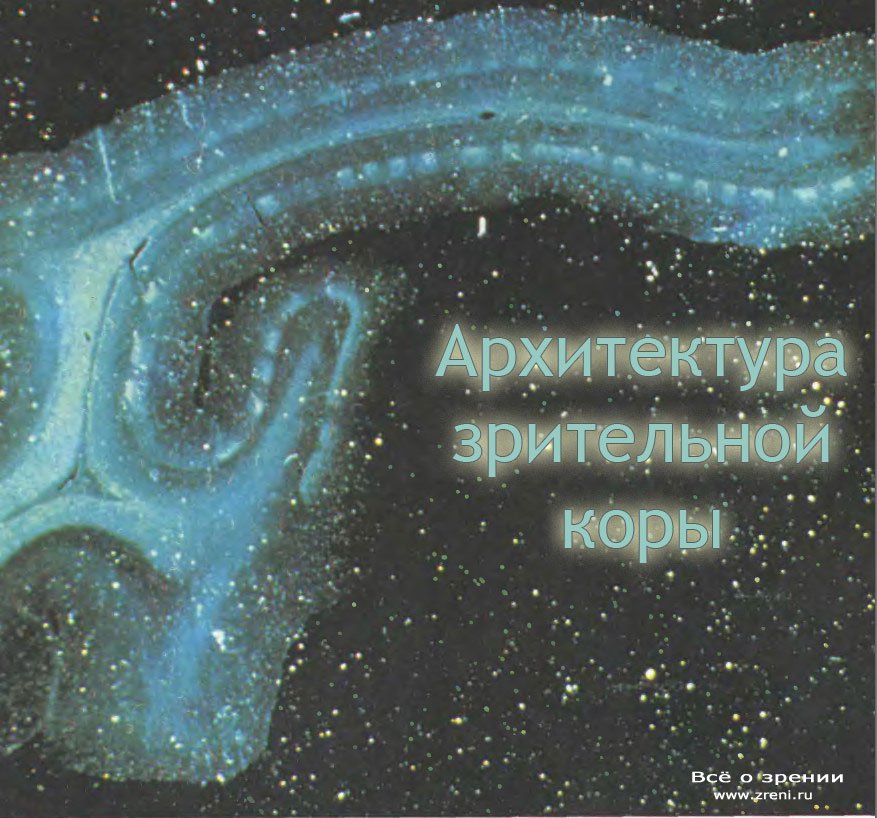
Первый способ опять-таки основан на использовании аксонного транспорта. Берут небольшое количество органического вещества (например, аминокислоты), меченного радиоактивным изотопом (например, 14С), и инъецируют в один глаз обезьяны, скажем в левый. Вещество захватывается клетками глаза, в том числе и ганглиозными клетками сетчатки. Помеченные молекулы, по-видимому уже включенные в состав белков, переносятся по аксонам ганглиозных клеток и их окончаниям в НКТ. Здесь они накапливаются в слоях, связанных с левым глазом. Для переноса метки требуется несколько дней. Затем из ткани приготовляют тонкие срезы, накладывают на них фотоэмульсию и оставляют на несколько дней в темноте. В результате получается радиоавтограф вроде показанного на рис. 73, где можно увидеть в каждом из двух НКТ по три слоя, связанных с левым глазом. Черные области, где образовались зерна, в одном НКТ расположены так, как светлые области в другом.

Для того чтобы наблюдать такую картину организации НКТ, нужно ввести в глаз не так много радиоактивного вещества. Если же при инъекции ввести его в достаточно большом количестве, то концентрация его в слоях НКТ станет настолько высокой, что часть радиоактивной метки перейдет из окончаний волокон зрительного нерва в клетки самих НКТ (в пределах меченых слоев) и будет транспортироваться по их аксонам в стриарную кору. В результате метка начнет накапливаться в окончаниях в слое 4С, образуя регулярный узор, соответствующий тому глазу, в который была сделана инъекция. Когда будет готов радиоавтограф (для этого нужно несколько месяцев, так как концентрация меченого вещества, дошедшего до коры, очень низка), мы действительно увидим на поперечном срезе коры места накопления метки в слое 4С (рис. 74).

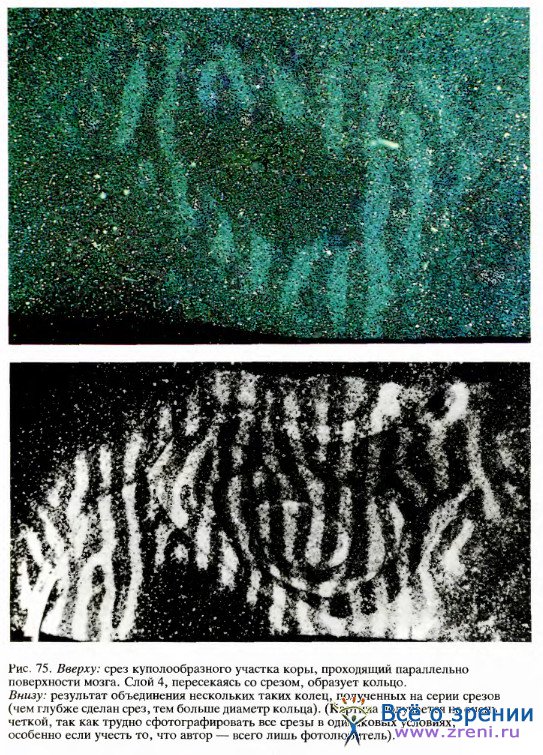
Если же сделать срез коры параллельно ее поверхности (сначала распрямив кору или же разрезав ее на части и скрепив затем вместе последовательные срезы), то мы, наконец, сможем увидеть распределение колонок так, как если бы смотрели на поверхность коры сверху. Четко выраженный набор параллельных полосок можно видеть как на одиночном срезе (рис. 75, вверху), так и на реконструированном изображении (рис.75, внизу).
На всех этих радиоавтографах коры меченые участки, т. е. проекции левого глаза, выглядят светлыми, а немеченые промежутки между ними — проекции правого глаза — темными. Поскольку из слоя 4 пути к верхним и нижним слоям идут главным образом в вертикальном направлении, прямо вверх и вниз, то в трехмерном пространстве зоны глазодоминантности образуют систему чередующихся «ломтиков», соответствующих правому и левому глазу (рис. 76).
Используя другой способ, С. Лe-Вэй успешно осуществил реконструкцию вида всей стриарной коры в затылочной доле. Участок коры, лежащий на поверхности мозга, показан на рис. 77.
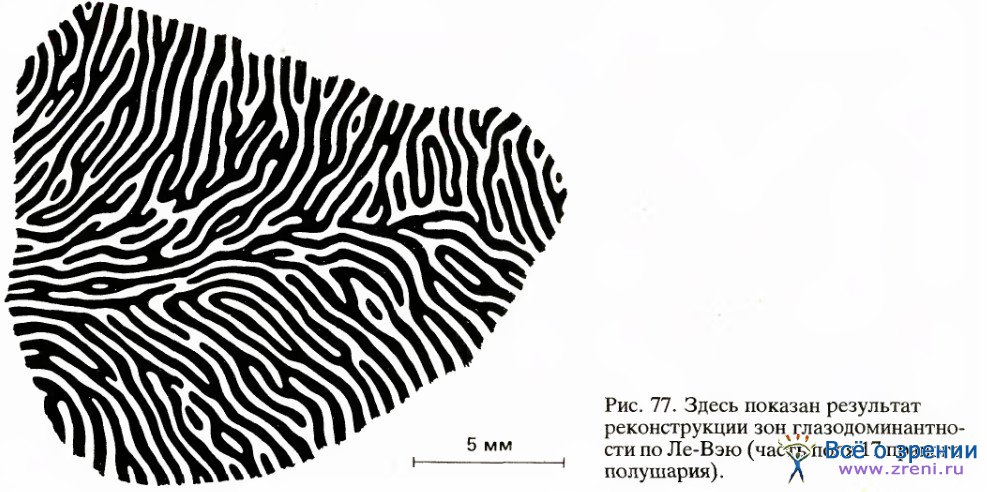
Распределение полос оказалось наиболее регулярным и четким на некотором расстоянии от корковой проекции центральной ямки. Вблизи этой проекции узор по неизвестным причинам наиболее сложен — он отличается регулярностью, но содержит много петель и завитков, поэтому он здесь мало похож на тот правильный узор, напоминающий рисунок обоев, который характерен для областей коры, более удаленных от проекции центральной ямки. Ширина полосок везде постоянна и равна примерно 0,5 мм. В процентном отношении корковые представительства правого и левого глаза примерно равны, пока речь идет о проекции центральной области сетчатки диаметром около 40°. Ле-Вэй и Д. ван Эссен обнаружили, что за пределами этой области из-за уменьшения вклада ипсилатерального глаза соответствующие ему полоски сужаются до 0,25 мм. Если же говорить о проекции зоны, удаленной от центральной ямки на 70—80°, то здесь, разумеется, представлен только ипсилатеральный глаз. Это естественно, так как поле зрения правого глаза простирается вправо дальше, чем влево.
Второй метод изучения колонок глазодоминантности позволяет выявлять их во всей толще коры, а не только в слое 4. Это метод с использованием меченой 2-дезоксиглюкозы, изобретенный в 1976 году Л. Соколоффом в Национальных институтах здоровья в Бетезде. Он тоже основан на способности радиоактивных веществ засвечивать фотопленку. В основе его лежит тот факт, что нейроны, как и большинство других клеток организма, поглощают глюкозу в качестве источника энергии, и чем интенсивнее им приходится работать, тем больше глюкозы они потребляют. Поэтому можно было бы представить себе следующую процедуру. Животному впрыскивают радиоактивную глюкозу, а затем стимулируют один глаз, например правый, в течение нескольких минут предъявляя какую-либо фигуру (время стимуляции должно быть достаточным для того, чтобы возбужденные клетки в мозгу поглотили введенную глюкозу). После этого мозг извлекают, делают срезы и покрывают их фотоэмульсией, а затем после надлежащей экспозиции получают, как и в предыдущем случае, радиоавтограф.
Однако на самом деле такая схема не годится, так как глюкоза, поглощенная клетками, расщепляется с образованием продуктов, которые быстро переходят обратно в кровь. Для того чтобы предотвратить эту утечку, Соколофф предложил остроумный прием — использовать при инъекции дезоксиглюкозу, которая по химической структуре очень близка к обычной глюкозе. «Обманутые» клетки поглощают ее и даже пытаются расщеплять; однако этот процесс прерывается после первого же шага; дезоксиглюкоза превращается в 2-дезоксиглюкозо-6-фосфат, который далее не может быть расщеплен. К счастью, это вещество нерастворимо в липидах клеточной мембраны и поэтому не может выйти из клетки. Оно накапливается в клетке в таком количестве, что уже может быть обнаружено с помощью радиоавтографии. Рассматривая результат этой процедуры — радиоавтограф, можно получить представление о том, какие участки мозга были наиболее активны в период стимуляции и накопили больше всего этой «фальшивой глюкозы».
Если бы в ходе опыта животное делало движения лапой, то на радиоавтографе выявился бы также соответствующий участок моторной зоны коры. То, что мы видим после стимуляции правого глаза, — это та часть вещества коры, которая была в наибольшей степени возбуждена данным стимулом, а именно набор колонок глазодоминантности для правого глаза. Примеры получаемых результатов представлены на рис. 78.
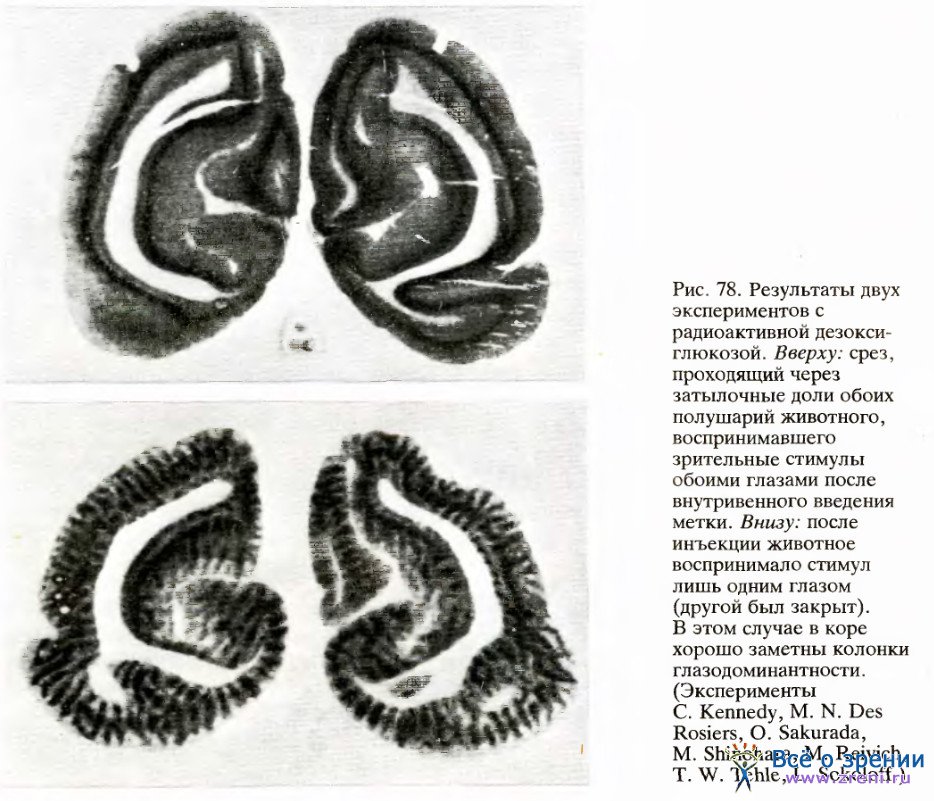
Идею того же метода исследования удачно развил Р. Тутелл из лаборатории Рассела де Валуа в Беркли, взяв в качестве стимула для животного, смотревшего на экран одним глазом, большую фигуру в виде концентрических окружностей и радиальных линий (рис. 79, вверху).

Получившаяся в результате картина корковой проекции тоже содержала круги и радиусы, но только в искаженном виде, что связано с неодинаковым увеличением разных участков сетчатки в их проекции на стриарную кору (с этим же связано и различие в остроте зрения на периферии сетчатки и в центральной ямке). Кроме того, каждая из окружностей и каждый из радиусов разбиты на множество мелких участков, соответствующих колонкам глазодоминантности. При одновременной стимуляции обоих глаз полосы были бы непрерывными. Редко так бывает, что в одном эксперименте удается столь четко продемонстрировать сразу несколько важных фактов.
Колонки глазодоминантности есть у кошек, некоторых низших обезьян, шимпанзе и человека. У грызунов и тупайи их нет. У саймири (род обезьян Нового Света) некоторые указания на возможное существование таких колонок получены в физиологических экспериментах, но современные методы морфологического анализа не выявляют их. В настоящее время нам неизвестна роль этого сложного распределения сигналов, приходящих от разных глаз; возможно, что оно имеет какое-то отношение к стереоскопическому зрению.
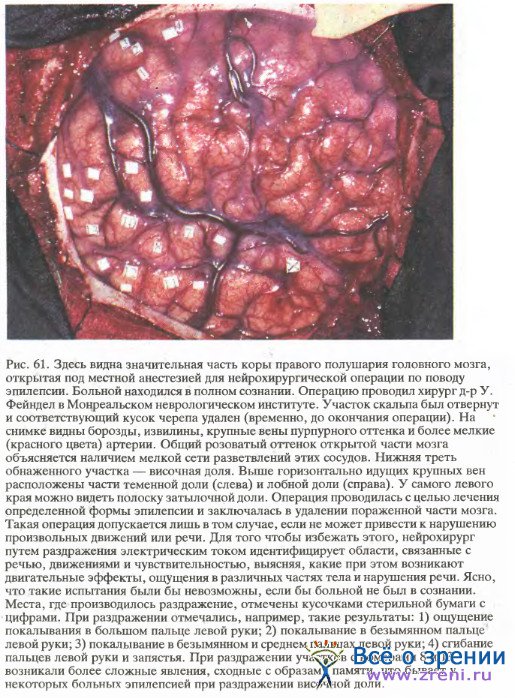
Подразделение на участки с функционально специализированными клетками было обнаружено не только в стриарной коре. Впервые такие участки описал в середине 1960-х годов В. Маунткасл в соматосенсорной коре. Это явилось наиболее важным открытием с того времени, когда были получены первые сведения о локализации функций мозга. Соматосенсорная область коры имеет такое же отношение к осязанию и проприоцепции, как стриарная кора — к зрению. Маунткасл показал, что эта область подразделяется на вертикально ориентированные зоны, внутри которых клетки чувствительны к прикосновению, и зоны, в которых клетки отвечают на сгибание в суставах или на приложение значительного давления к конечности. Так же как и в случае колонок глазодоминантности, ширина этих зон составляет примерно 0,5 мм. Однако еще не ясно, образуют ли эти зоны полосы, шахматный рисунок или же отдельные островки на общем фоне. Маунткасл назвал их колонками, и можно думать, что он мысленно представлял себе некую сотовидную структуру.
Теперь мы уже знаем, что для зрительной коры более подходящим был бы термин пластина или слэб (slab). Однако введенную терминологию очень трудно изменить, поэтому лучше всего, наверное, сохранить прежний термин, несмотря на его недостатки. Сегодня мы говорим о колончатой организации, когда некоторое свойство клеток остается постоянным во всей толще коры от ее поверхности до белого вещества, но изменяется в направлениях, параллельных поверхности коры. По причинам, которые будут ясны из следующей главы, мы обычно не применяем этот термин, когда речь идет о топографическом отображении, т, е. о проекции расположения рецептивных полей на сетчатке или на поверхности тела.
---
Статья из книги: Глаз, мозг, зрение | Хьюбел Д.

Комментариев 2