Ориентационные колонки

Описание

Регистрируя реакции нейронов стриарной коры, мы уже в самом начале заметили, что всякий раз, когда одновременно отводится активность двух клеток, эти клетки оказываются сходными не только по глазодоминантности, но и по оптимальной ориентации стимула. Возникает вопрос: однотипны ли соседние клетки и по всем другим свойствам? Ответ будет отрицательным. Как я уже упоминал, положения рецептивных полей в большинстве случаев не вполне совпадают, хотя поля обычно перекрываются; дирекциональная чувствительность часто бывает противоположной, или же у одной клетки она может быть хорошо выражена, а у другой ее может не быть вовсе. В слоях 2 и 3, где встречаются клетки, реагирующие на концы линий, одна клетка может не проявлять совсем этого свойства, а соседняя — обладать им в полной мере. С другой стороны, две соседние клетки очень редко обнаруживают явное различие в оптимальной ориентации стимула или противоположную глазодоминантность.
Предпочитаемая ориентация, так же как и глазодоминантность, остается неизменной при прохождении электрода вертикально сквозь всю толщу коры. Как уже говорилось, в слое 4Сb клетки вообще не обладают избирательностью к ориентации стимула; но как только мы доходим до слоя 5, у клеток выявляется сильно выраженная настройка на определенную ориентацию, причем оптимальная ориентация здесь та же, что была выше слоя 4. Если теперь вынуть электрод и ввести его в каком-нибудь другом месте, то общая картина останется прежней, только ориентация скорее всего будет уже другой. Таким образом, кора разбита на узкие участки с постоянной предпочтительной ориентацией, которые идут от поверхности коры до белого вещества, но прерываются з слое 4, где клетки не обладают ориентационной избирательностью.
Если же, наоборот, вводить электрод параллельно поверхности коры, то наблюдается удивительно закономерное изменение предпочитаемой ориентации — каждый раз, когда электрод перемещается на 0,05 мм (50 мкм), ориентация сдвигается в среднем на 10° по часовой стрелке или против часовой стрелки. Поэтому при продвижении электрода на 1 мм она обычно меняется на противоположную. Величины 50 мкм и 10° близки к пределу доступный ныне точности измерений, так что нельзя сказать определенно, меняется ли ориентация при смещении электрода непрерывно или же сдвигается скачками.
На рис. 80 и 81 представлена часть типичного эксперимента, в котором электрод продвигался в поле 17 в направлении, близком к горизонтальному. В этом опыте точки фиксации двух глаз на экране не вполне совпадали (из-за наркоза и введения вещества, расслабляющего мышцы) — расстояние между ними составляло около 2°.
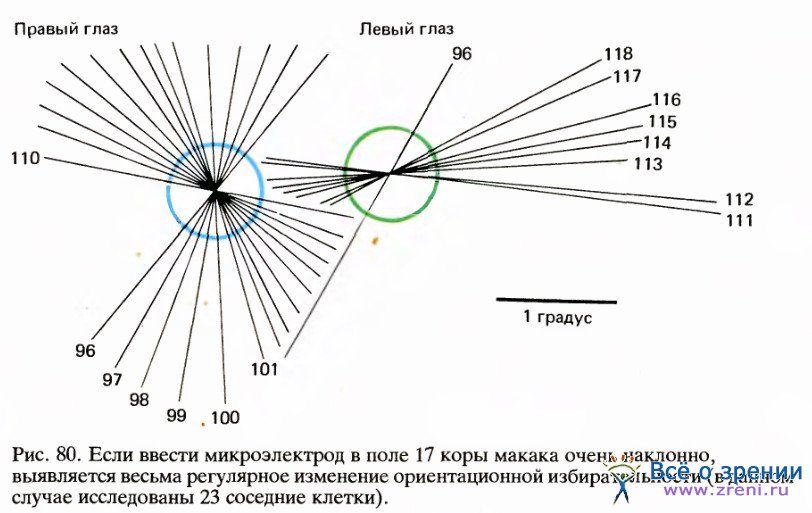
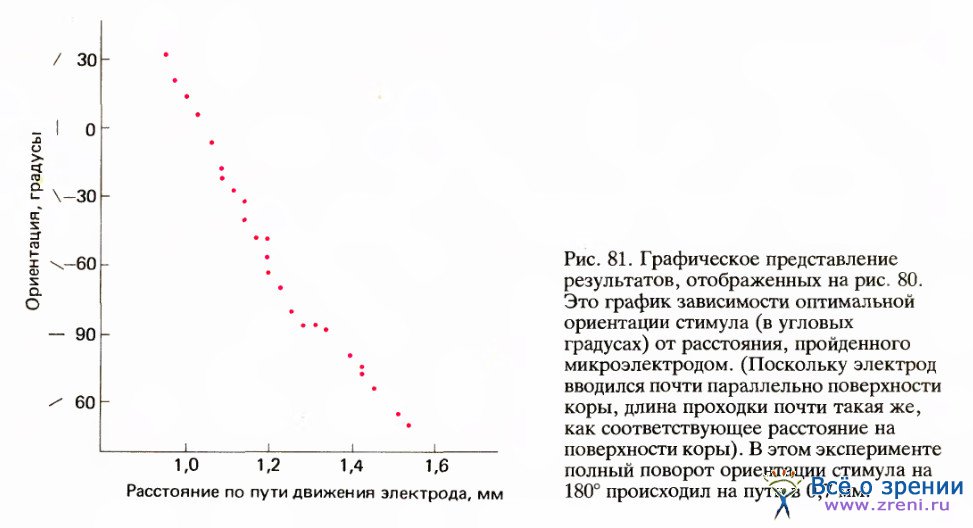
Цветные круги на рис. 86 примерно соответствуют размерам рецептивных полей (около 1° в диаметре), расположенных в 4° ниже и слева от центральных ямок (регистрировались ответы клеток правого полушария). Первая клетка, отмеченная номером 96, была бинокулярной, однако для следующей, 114, явно доминирующим оказался правый глаз. Затем идут клетки под номерами от 111 до 118 с доминированием левого глаза. Легко заметить, как регулярно изменяется ориентация в последовательности клеток, в данном случае в направлении против часовой стрелки. Если построить график зависимости ориентации от продвижения электрода (рис. 87), то все точки лягут на линию, близкую к прямой.

Переход от одного глаза к другому не сопровождался никакими резкими изменениями ни в направлении сдвига ориентации, ни в наклоне линии на графике. Согласно нашей интерпретации, это означает, что между двумя системами группировки — по глазодоминантности и по ориентации стимула — нет тесной связи. Это выглядит так, как если бы кора размечалась двумя совершенно независимыми способами.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
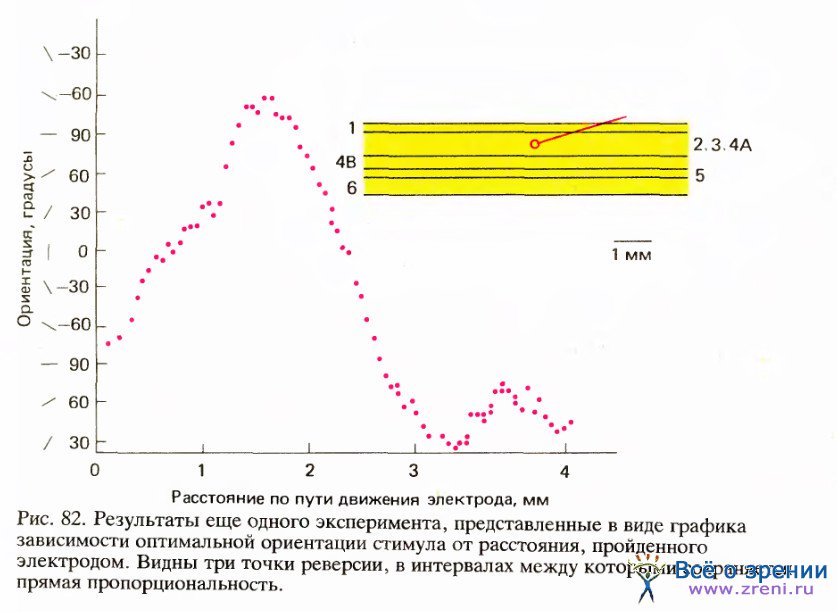
Когда электрод продвигается параллельно поверхности коры, оптимальная ориентация стимула может изменяться либо по часовой стрелке, либо в обратном направлении. В большинстве случаев при достаточно длинном пути электрода направление сдвига ориентации рано или поздно меняется на обратное. Момент такой смены, или реверсии, непредсказуем, но обычно она происходит с интервалами в несколько миллиметров. На рис. 82 приведен пример таких последовательных смен направления.
Наконец, в некоторых экспериментах мы обнаружили еще одну особенность, которую назвали разрывом. Как раз тогда, когда нас начинала гипнотизировать монотонная регулярность постепенного изменения ориентации в одном и том же направлении, этот ход событий вдруг — в редких случаях — прерывался, и происходил сдвиг ориентации сразу на 45—90и. После этого опять шла прежняя регулярная последовательность, только направление ее часто менялось на обратное. На рис. 83 приведен пример таких разрывов, находящихся друг от друга на расстоянии в несколько десятых миллиметра.

Как выглядели бы участки постоянной ориентации при взгляде на кору сверху? Ответить на этот вопрос оказалось гораздо труднее, чем на аналогичный вопрос о колонках глазодоминантности. Вплоть до самого последнего времени мы не имели прямой возможности «видеть» ориентационные группировки и могли лишь попытаться логически вывести их форму из результатов микро-электродных исследований вроде описанных выше. Наличие как реверсий, так и разрывов наводит на мысль о сложности интересующей нас конфигурации. С другой стороны, линейная регулярность, которую мы наблюдали часто миллиметр за миллиметром вдоль коры, может означать периодичность, по крайней мере в пределах небольших зон коры; тогда реверсии и разрывы могли бы указывать на то, что эта периодичность прерывается через каждые несколько миллиметров.
В тех областях, где периодичность сохраняется, можно с определенной вероятностью реконструировать искомую конфигурацию. Предположим, эта конфигурация такова, что в пределах данной области при продвижении электрода параллельно поверхности коры мы наблюдаем периодичность без реверсий и разрывов, т. е. везде получаем такой график, как на рис. 81. Если бы мы имели достаточное число таких графиков, то смогли бы реконструировать трехмерную структуру (рис. 84); здесь вертикальная ось (z) отображает ориентацию, а горизонтальные оси (х и у) — расстояния в плоскости коры.

В этом случае ориентации отображались бы на некоторой поверхности; например, если график — прямая линия, то эта поверхность имела бы вид наклонной плоскости, что показано на рис. 84, а в других случаях это была бы криволинейная поверхность. На таких трехмерных реконструкциях искомые поверхности пересекались бы горизонтальными плоскостями (плоскостью х — у или плоскостями, параллельными ей) по некоторым линиям, соответствующим постоянной ориентации (можно назвать эти линии изоориентационными), что аналогично изогипсам на географических картах.
Неровности рельефа — холмы, впадины, горы — на такой трехмерной реконструкции дадут в некоторых участках реверсии на графиках зависимости ориентации от расстояния, а резкие перепады высот в виде отвесных скал приведут к разрывам. Основной вывод из этих рассуждений заключается в том, что наличие зон регулярности означает возможность построить карту с изолиниями, откуда следует, что области постоянной ориентации, если рассматривать их сверху, должны иметь вид полос. Поскольку при вертикальной проходке коры предпочитаемые ориентации остаются неизменными, в объемном представлении эти области должны иметь вид пластин. А поскольку изоориентационные линии могут быть криволинейными, эти пластины не обязательно будут телами правильной формы наподобие ровно нарезанных ломтей хлеба. Многое из сказанного выше было продемонстрировано в прямых экспериментах, в которых производились две или три проходки в параллельных направлениях на расстоянии не более миллиметра друг от друга. Результатом этих экспериментов была реконструкция объемной картины по крайней мере в пределах небольшой исследованной области.
Если наши рассуждения правильны, то при выполнении множества произвольных проходок некоторые из них случайно будут совпадать с направлением ориентационных изолиний, и тогда ориентация должна оставаться неизменной. Такое действительно случается, но не очень часто. Этого и следовало ожидать, так как тригонометрия подсказывает, что даже небольшое отклонение направления проходки от изолинии должно приводить к довольно большим изменениям в ориентации; поэтому лишь в очень немногих случаях графики зависимости ориентации от глубины введения электрода могут оказаться горизонтальными линиями.
Число градаций ориентации, представленных в одном квадратном миллиметре коры, можно определить по наиболее крутым графикам, получаемым в эксперименте. Эта цифра составляет примерно 400 градусов на 1 мм, откуда следует, что полный набор ориентаций в пределах 180 градусов может содержаться на участке длиной около 0,5 мм. Эту величину нужно будет вспомнить, когда мы вернемся к вопросу о топографии коры и ее удивительной однородности. Однако, забегая вперед, хочу отметить, что толщина пары колонок глазодоминантности составляет примерно 0,4 + 0,4 мм, т. е. близка к 1 мм. Это вдвое больше толщины полного набора ориентационных пластин, однако порядок величины тот же.
Вскоре, однако, мы воспользовались методом картирования с помощью дезоксиглюкозы, который позволил непосредственно изучать геометрию ориентационных колонок. Для этого в качестве стимула мы взяли просто рисунок из параллельных полос с неизменной ориентацией, скажем вертикальной, который предъявляли на протяжении всего эксперимента. Полученная при этом картина (рис. 85) оказалась гораздо более сложной, чем картина колонок глазодоминантности.
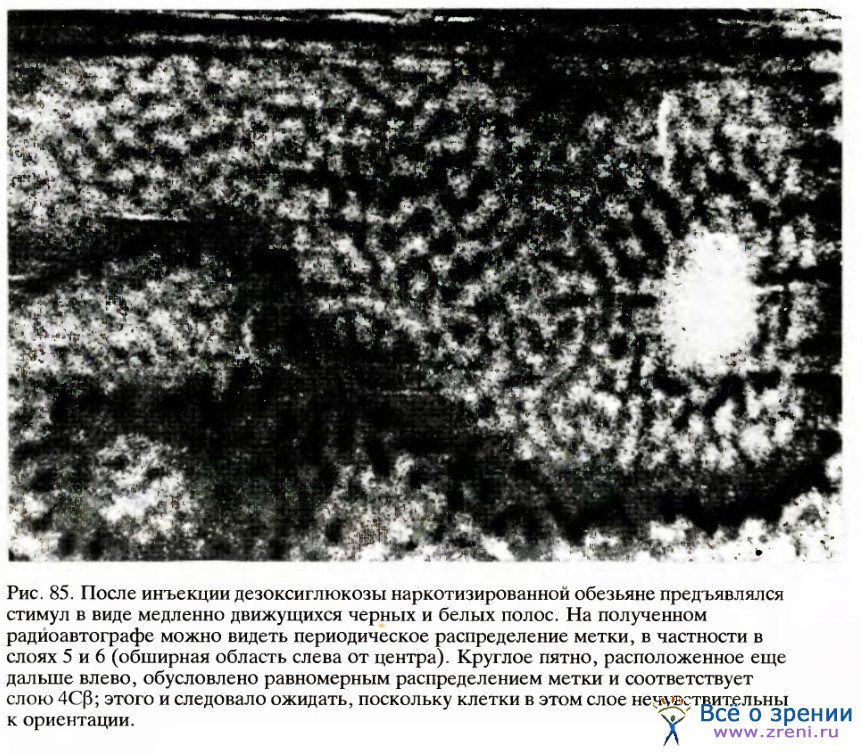
Однако и здесь отчетливо была видна регулярность распределения с периодом 1 мм или чуть меньше (расстояние от середины одной темной полосы до середины следующей). Это согласуется с электрофизиологическими данными (с тем расстоянием, на которое нужно передвинуть электрод от клеток с определенной ориентацией, скажем вертикальной, чтобы через все промежуточные варианты снова дойти до вертикальной ориентации). В некоторых местах мы находили структуру в виде правильных полос на площади в несколько квадратных миллиметров.
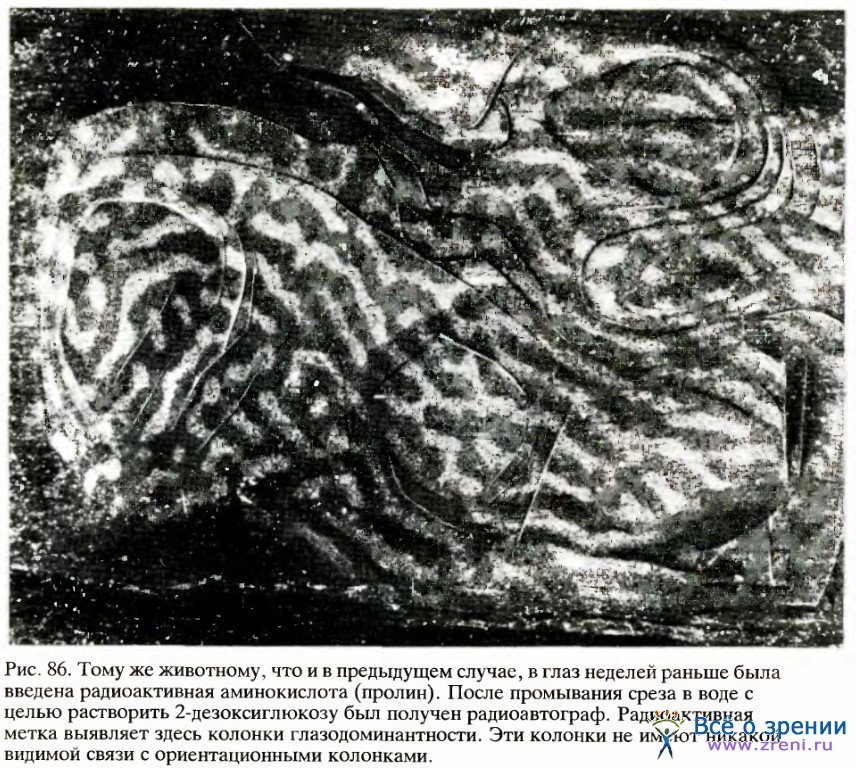
Мы задавались вопросом: нет ли какой-то связи между конфигурациями ориентационных полос и полос глазодоминантности — может быть, например, они идут параллельно или же пересекаются под прямым углом? В том же самом эксперименте нам удалось выявить глазодоминантные колонки, после того как в глаз животного была введена радиоактивная аминокислота, а затем полученные срезы ткани анализировались с помощью двух методик (результат показан на рис. 86). Мы не смогли обнаружить никакой явной корреляции. Сравнивая сложную картину ориентационных колонок с гораздо более простым распределением колонок глазодоминантности, очень трудно было решить, какая могла бы быть связь между ними.
У метода с применением дезоксиглюкозы есть один серьезный недостаток: никогда нет полной уверенности в том, что полученная картина распределения действительно связана с интересующим нас свойством стимула. Например, когда в качестве стимула используется рисунок из черных и белых вертикальных полос, как можно убедиться, что получившееся распределение связано именно с ориентацией этих полос, т. е. что в темных областях находятся клетки, отвечающие на вертикальные линии, а в светлых — клетки, которые на них не реагируют? Ведь вместо этого решающей особенностью данного стимула могло бы быть то, что он черно-белый, а не цветной, или же то, что он состоит из широких, а не узких полос, или, наконец, то, что экран расположен именно на таком, а не ином расстоянии от животного.
Одно из косвенных подтверждений того, что действие дезоксиглюкозы затрагивает механизмы настройки на ориентацию, — это отсутствие каких-либо пятен или полос в слое 4С, где клетки не обладают избирательностью к ориентации. Другим подтверждением служит результат, полученный в исследованиях М. Страйкера из Калифорнийского университета в Сан-Франциско. Страйкер делал протяженные проходки микроэлектродом в стриарной коре кошек параллельно ее поверхности и каждый раз, когда встречались клетки с определенной оптимальной ориентацией, производил локальное разрушение ткани; затем после инъекции радиоактивной дезоксиглюкозы включался стимул с полосками одинаковой ориентации. В этих экспериментах была обнаружена явная корреляция между получаемым распределением метки на гистологическом срезе и ориентацией стимула.
Наиболее четко удалось недавно выявить ориентационные колонки с помощью красителей, чувствительных к электрическому напряжению, которые в течение многих лет разрабатывал Л. Коэн в Йеле. Г. Блэсдел из Питтсбургского университета применил их при исследовании коры головного мозга. При использовании этого метода краситель, чувствительный к напряжению и окрашивающий клеточные мембраны, наносят на поверхность коры наркотизированного животного, и нервные клетки поглощают его. Когда в эксперименте животному предъявляют стимул, все реагирующие на него клетки изменяют свой цвет. Если в области, близкой к поверхности коры, таких клеток окажется достаточно много, эти изменения цвета можно уловить с помощью современных методов обработки телевизионных изображений.
Хотя изменения цвета едва заметны, с помощью чувствительных телекамер и машинных методов фильтрации шумов их можно обнаружить. Блэсдел использовал в своих опытах стимуляцию полосами определенной ориентации, после чего фотографировал полученное распределение активности клеток на участке площадью в несколько квадратных сантиметров. Затем та же процедура повторялась при многих других ориентациях стимула. После этого каждой ориентации был поставлен в соответствие свой цвет — красный для вертикальной, оранжевый для положения часовой стрелки в 1 час и т. д.; потом все изображения были наложены друг на друга. Поскольку изоориентационная линия должна постепенно смещаться по мере изменения ориентации, в любой небольшой зоне изображения должен возникнуть радужный узор. Именно это и обнаружил Блэсдел в действительности. Примеров успешного применения этого метода пока еще мало, и нет возможности интерпретировать получаемые изображения в таких понятиях, как разрывы и реверсии, однако метод представляется многообещающим.
Карты коры
Теперь, когда мы узнали кое-что о топографическом представлении в коре параметров ориентации и глазодоминантности, мы можем рассмотреть взаимосвязь между этими отображениями и проекциями зрительных полей. Обычно говорят, что сетчатка отображается на кору взаимно однозначно. Однако, учитывая то, что нам известно о рецептивных полях корковых клеток, можно с уверенностью сказать, что это утверждение, строго говоря, неверно, поскольку каждая клетка получает входные сигналы от тысяч палочек и колбочек, а ее рецептивное поле — далеко не точка. Отображение сетчатки на кору гораздо более сложно, чем любое обычное взаимно однозначное отображение. На рис. 88 я попытался показать распределение в коре тех областей, которые активируются под действием простого стимула (не следует смешивать это с рецептивным полем одиночного нейрона).
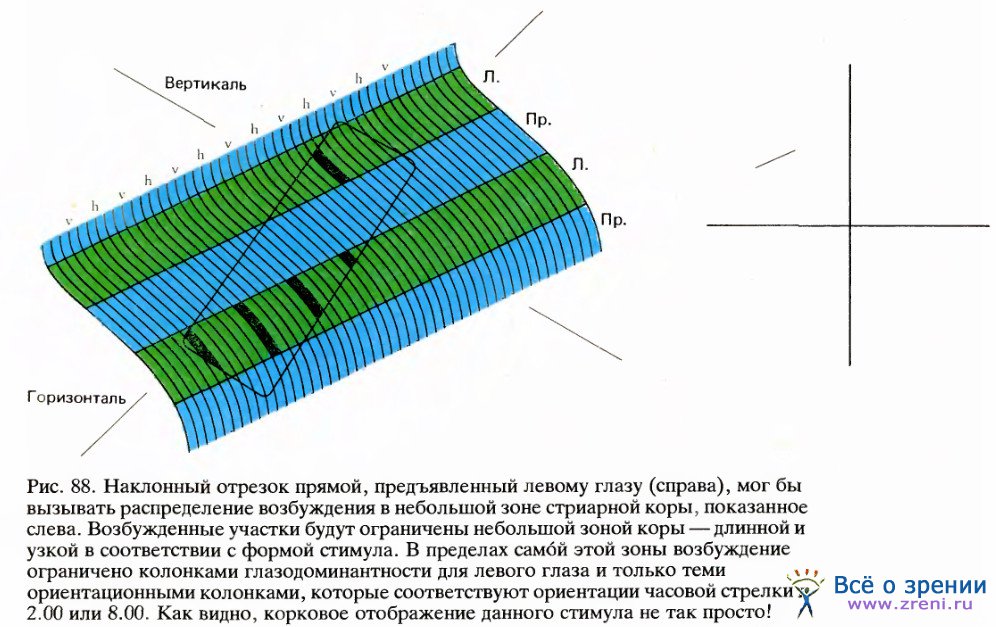
Стимул здесь — это короткий отрезок линии с наклоном в 60°относительно вертикали, предъявленный только левому глазу. Мы предполагаем, что этот элемент поля зрения проецируется на область коры, выделенную прямоугольником с закругленными углами. В пределах этой области будут возбуждены только те полосы, которые связаны с левым глазом, а в этих полосах — только те участки, которые соответствуют ориентации в 60°. Все они на рисунке зачернены. Таким образом, линия, появившаяся в поле зрения, вызывает причудливую картину распределения возбужденных клеток коры в виде ряда узких полосок.
Теперь становится понятно, насколько наивно представление о зеленом гомункулюсе, который сидит у нас в голове и рассматривает передаваемую в кору картину. На самом деле картина, которая создается в коре, имеет примерно такое же отношение к изображению внешнего мира, как электрические процессы в телекамеры — к передаваемой внешней сцене. Если бы распределение корковой активности точно воспроизводило распределение яркости в поле зрения, то это означало бы, что на пути от сетчатки в кору не произошло ничего существенного. В этом случае действительно потребовался бы маленький зеленый человечек.
Вряд ли можно представить себе, что в процессе эволюции было затрачено столько усилий на столь изящную группировку клеток в двух взаимонезависимых системах колонок, если бы это не обеспечивало какой-то пользы для животного. Пока мы не докопаемся до точной схемы связей, определяющей отображение сетчатки в коре, мы вряд ли сможем до конца понять принципы группировки клеток в коре. Сейчас мы можем только строить логичные догадки. Если мы предположим, что схема, приведенная в предыдущей статье, соответствует реальности, то придется признать: то, что нужно для перехода от простых клеток к сложным или для создания чувствительности к концам линий или к направлению движения, — это определенного рода конвергенция множества клеток на одну клетку, причем у рецептивных полей промежуточных клеток должна быть одна и та же ориентация и примерно одно и то же положение.
Пока у нас не было особых оснований предполагать, что клетка с определенной ориентацией рецептивного поля получает входы от клеток с рецептивными полями иной ориентации. (Я здесь несколько преувеличиваю: высказывались предположения, что клетки с разной оптимальной ориентацией стимулов могут быть соединены тормозными связями; в пользу этого имеются лишь косвенные данные, на мой взгляд, не очень убедительные, хотя их нельзя игнорировать.) Если это так, то почему бы не группировать вместе клетки, которые должны быть взаимно связаны? Альтернативный вариант едва ли целесообразен — легко представить себе, как трудно было бы обеспечить надлежащие связи между клетками, если эти клетки разбросаны в коре без учета их сходных свойств! Особенно много связей должно быть между клетками с одинаковой оптимальной ориентацией, и если бы такие клетки распределялись случайным образом, то сеть аксонов, необходимых для их соединения, заполнила бы все пространство между нейронами. Как бы то ни было, фактически они группируются вместе. Изложенные соображения относятся и к клеткам, сходным по глазодоминантности.
Но если клетки со сходными свойствами упаковываются вместе, то почему, продвигая электрод, мы наблюдаем последовательное изменение ориентации небольшими шагами? И чем объясняется цикличность — почему, пройдя все возможные варианты, мы опять возвращаемся к исходной ориентации, а потом весь цикл повторяется снова? Казалось бы, можно было собрать вместе все клетки с ориентацией 30°, все клетки с ориентацией 42° и т. д.; и наконец, объединить все клетки с доминированием левого глаза и все — с доминированием правого глаза. Если бы мы знали, как устроена кора, то смогли бы предложить много ответов на эти вопросы. Сейчас выскажем одно предположение: возможно, что клетки с разной ориентацией действительно оказывают тормозящее воздействие друг на друга.
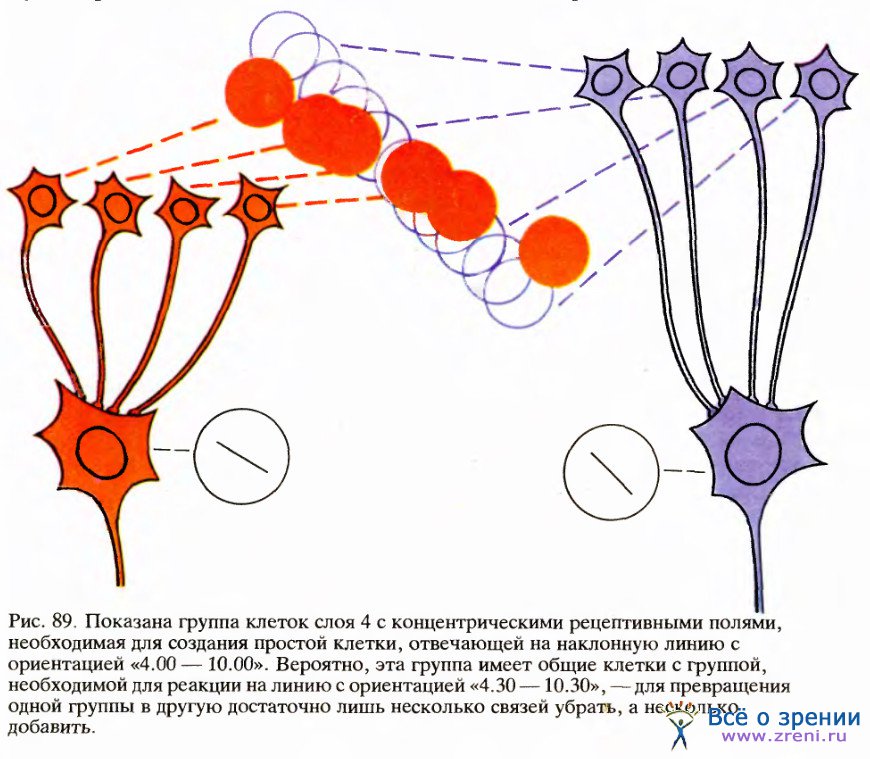
Если нежелательно, чтобы какая-то клетка отвечала на другие ориентации, отличные от ее собственной, то нетрудно представить себе наличие тормозных связей, которые обостряют настройку данной клетки на определенную ориентацию. Тогда наблюдаемая картина — как раз то, что нам нужно: клетки с одинаковой ориентацией расположены ближе всего друг к другу, но не слишком далеко от клеток с иной, но очень сходной ориентацией; тогда тормозные связи будут достаточно короткими. Возможно другое предположение: если рассмотреть, какие входы от клеток слоя 4 (с круглыми полями) нужны для организации простой клетки с определенной оптимальной ориентацией, легко увидеть, что потребуется лишь небольшое изменение набора входов для того, чтобы получить близко расположенную простую клетку с близкой, но несколько иной ориентацией. Для этого достаточно добавить несколько новых входов, а несколько прежних убрать (рис. 89). Такого рода соображения позволили бы объяснить физическую близость расположения клеток со сходной ориентацией.
Предметом следующей главы будет взаимосвязь между ориентацией, глазодоминантностью и проекцией поля зрения на кору. Рассмотрение этого вопроса может помочь нам понять, почему нужно так много колонок. Когда к этому добавится еще вопрос о топографии, соответствующая корковая организация будет представляться еще более сложной и интересной.
---
Статья из книги: Глаз, мозг, зрение | Хьюбел Д.

Комментариев 0