Ганглиозные клетки сетчатки некоторых млекопитающих


Содержание:
Описание
Нейроны сетчатки млекопитающих изучены Ramon у Cajal [1972] на примере главным образом быка и собаки. Однако им были просмотрены также импрегнированные по Гольджи сетчатки мыши, овцы, лошади, кошки. Ramon у Cajal считал, что морфология каждого типа клеток постоянна для всех видов млекопитающих. В течение последних десятилетий многие ученые исследовали морфологию ганглиозных клеток сетчатки у разных млекопитающих: приматов — Polyak [1941]; Van Buren [1963]; Boycott, Dowling [1969]; Kolb [1970]; кошки — Gallego, Crus [1965]; Leicester, Stone [1967]; С. А. Шибкова [1967, 1983]; С. А. Шибкова, Б. М. Владимирский [1969]; Shkolnik-Yarros [1971]; Е. И. Койдан [1972]; Boycott, Wassle [1974]; Famiglietti, Kolb [1976]; Stone, Clarke [1980]; Kolb, Nelson [1980]; Kolb, Nelson, Mariani [1981]; грызунов — E. Г. Школьник-Яррос [1974]; Bunt [1976]; West [1976] и многие другие.Сравнение фактических данных, приведенных в названных работах, приводит к предварительному выводу о некоторых различиях в расслоенности внутреннего плексиформного слоя и, следовательно, стратификации дендритов ганглиозных клеток у различных млекопитающих.
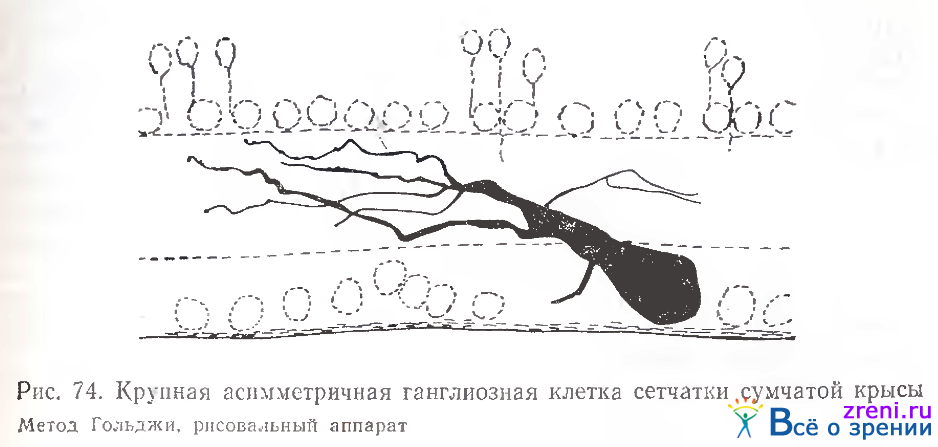
Перед детальным изложением данных по сетчатке белки и кошки приводим две зарисовки ганглиозных клеток сетчатки сумчатой крысы (рис. 74)
и ежа (рис. 75).
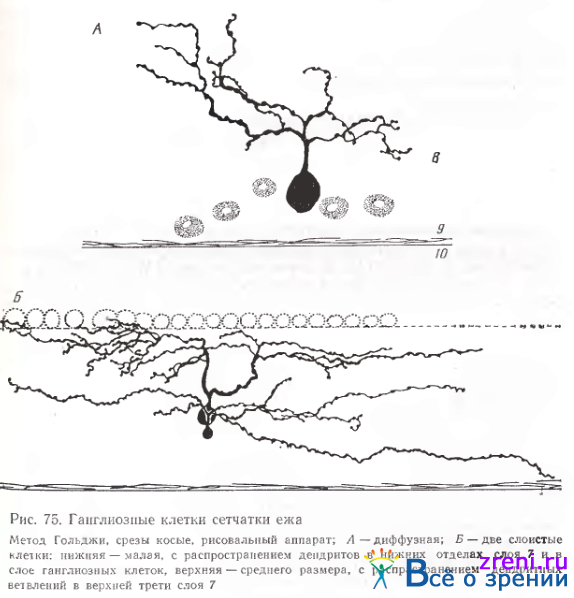
Не имея полных данных по морфологии нейронов у этих видов млекопитающих, мы все же сочли возможным поместить здесь примеры их ганглиозных клеток ввиду отсутствия подобных описаний в доступной нам литературе.
Клетка на рис. 74 характеризуется асимметрией в расположении отростков. В сетчатке ежа найдены примеры как диффузного редковетвистого нейрона (рис. 75, А), так и слоистых клеток (рис. 75, Б).
↑ Сетчатка белки
Нам не удалось найти в литературе данных об общей картине нейронов в сетчатке белки, кроме нашего краткого сообщения [Школышк-Яррос, 1974]. В сетчатке суслика [West, 1976], грызуна, весьма близкого белке по морфологии зрительной системы, обнаружено 15 типов ганглиозных клеток, различающихся по густоте, уровню, форме и размаху дендритных ветвлений.
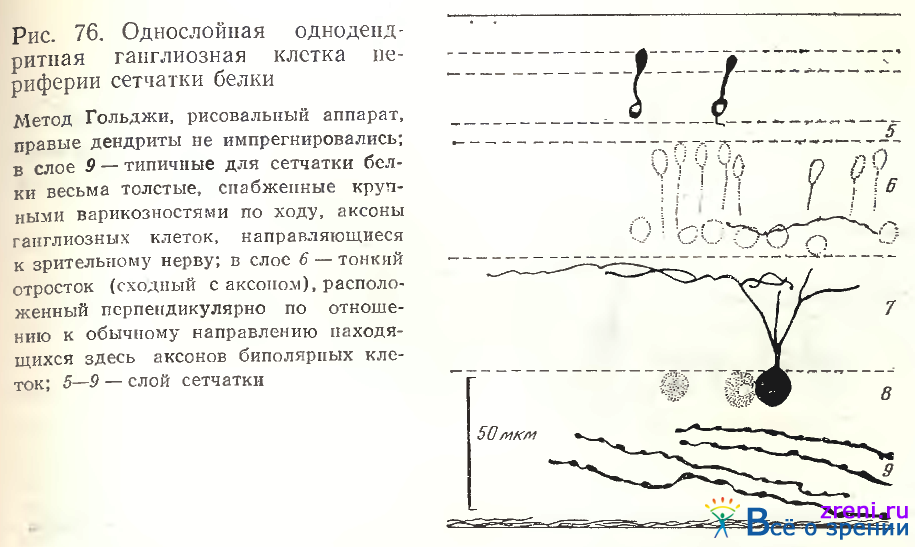
Наблюдались следующие разновидности однослойных ганглиозных клеток сетчатки белки: с ветвлением дендритов в самом наружном отделе внутреннего плексиформного слоя (рис. 76),
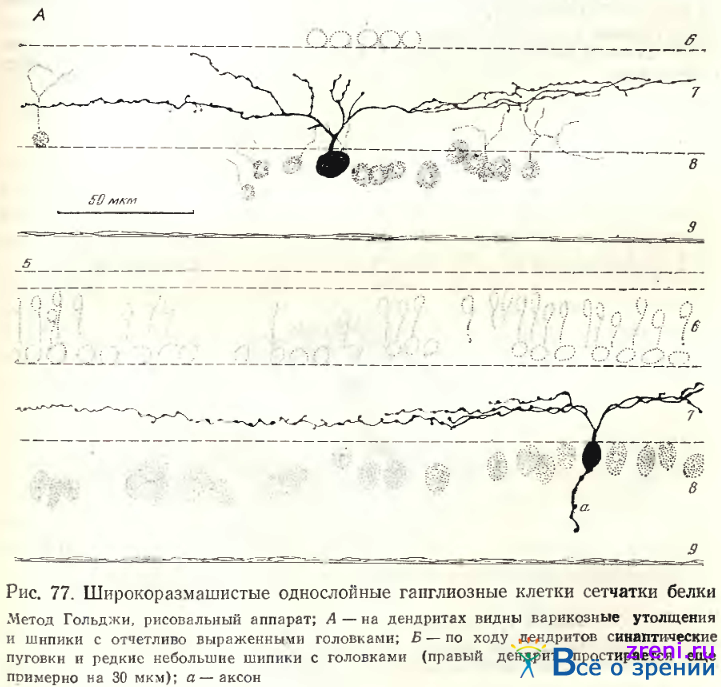
в центре его (рис. 77),
в самом внутреннем отделе слоя или непосредственно над телами ганглиозных клеток [Школьник-Яррос, 1974].
Наибольший размах дендритных ветвлений отличал однослойные нейроны, простирающиеся в центре или нижней трети внутреннего плексиформного слоя (рис. 77). Это наблюдение совпадает с фактами, приведенными West по отношению к сетчатке суслика. На косотангенциальных срезах дендритное поле однослойных нейронов имеет вид веера. Па этих препаратах особенно четко выявляются небольшие овальные утолщения по ходу дендритов и редкие шипики с головками (рис. 78).
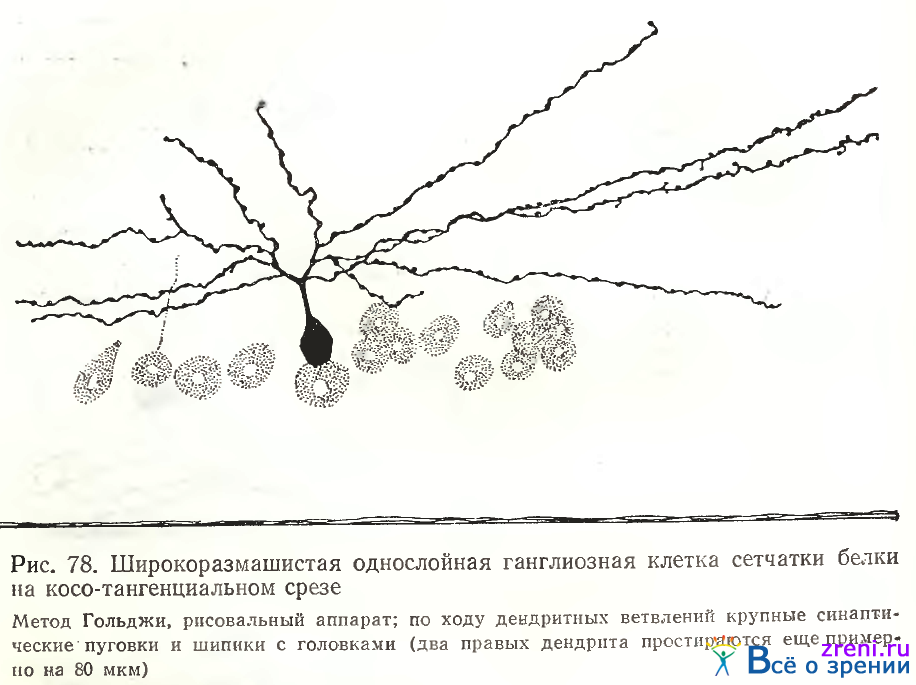
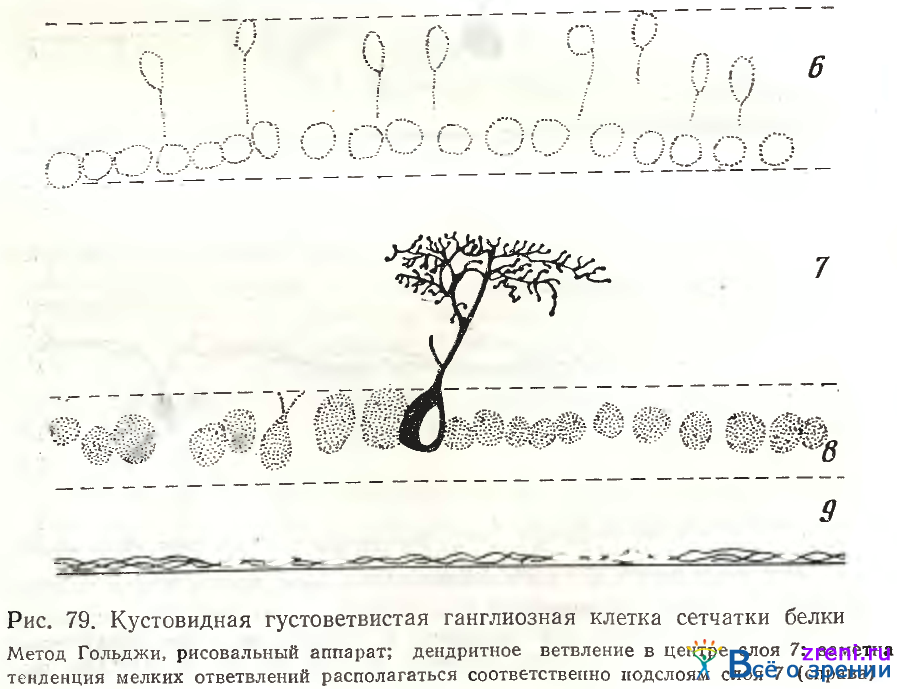
Однослойные нейроны могут быть симметричными и асимметричными. Как и в сетчатке лягушки (см. рис. 63) ветвления асимметричных нейронов чаще наблюдались в нижних отделах внутреннего плексиформного слоя. Кустовидные нейроны, составляющие большую часть ганглиозных клеток в сетчатке лягушки, встречаются в сетчатке белки не так часто (рис. 79).
Многослойные клетки двух-, трехслойные и с большим числом уровней ветвлений наблюдаются чаще. Кроме узкого либо широкого размаха их дендритного ветвления, его густоты, их отличает также наличие либо гладких мало извитых отростков, либо извилистых густо покрытых синаптическими образованиями ветвлений. Представленную на рис. 80, А
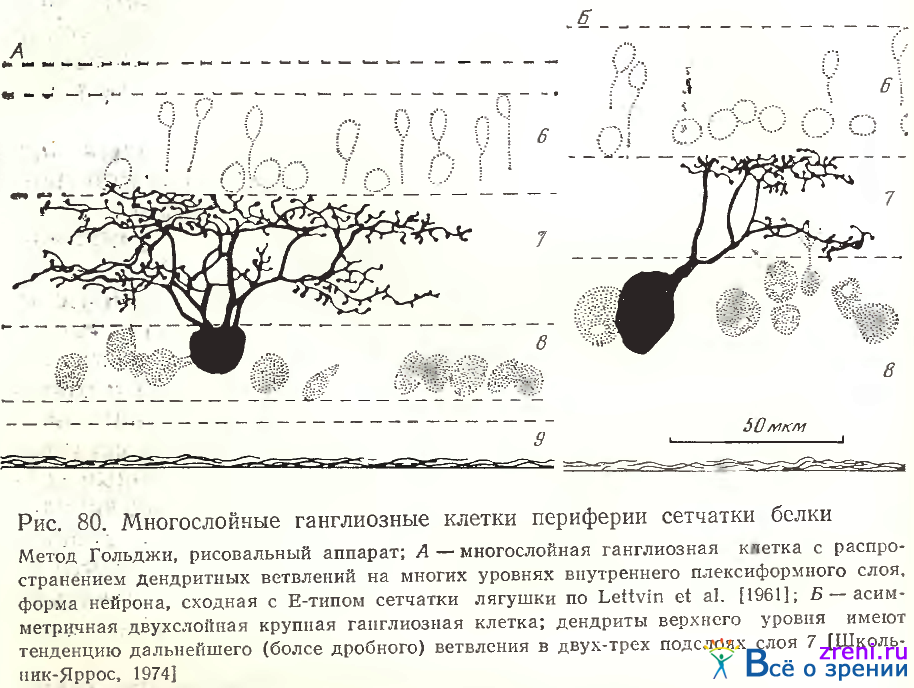
клетку характеризует скопление основных ветвлений в двух подслоях внутреннего плексиформного слоя. Дендритные веточки весьма извилисты, покрыты по ходу множеством пуговчатых образований и отчетливыми шиниками с головками. Величина синаптических образований заметно больше, чем у таких же нейронов в сетчатке лягушки. В сетчатке суслика, по West [1976J, двухслойные нейроны весьма густоветвисты, и различается несколько их вариантов по уровням расположения ветвлений (G2, G3, G4). Нейрон на рис. 80, А несколько отличен от клеток в сетчатке суслика большим размахом отростков; в целом его вид более сходен с Е- типом клетки в сетчатке лягушки по классификации Lettvin et al. [1961].
На наших препаратах сетчатки белки наблюдались также трех- и многослойные ганглиозные клетки. Трехслойные клетки наблюдаются узковетвистые и широковетвистые (рис. 81).
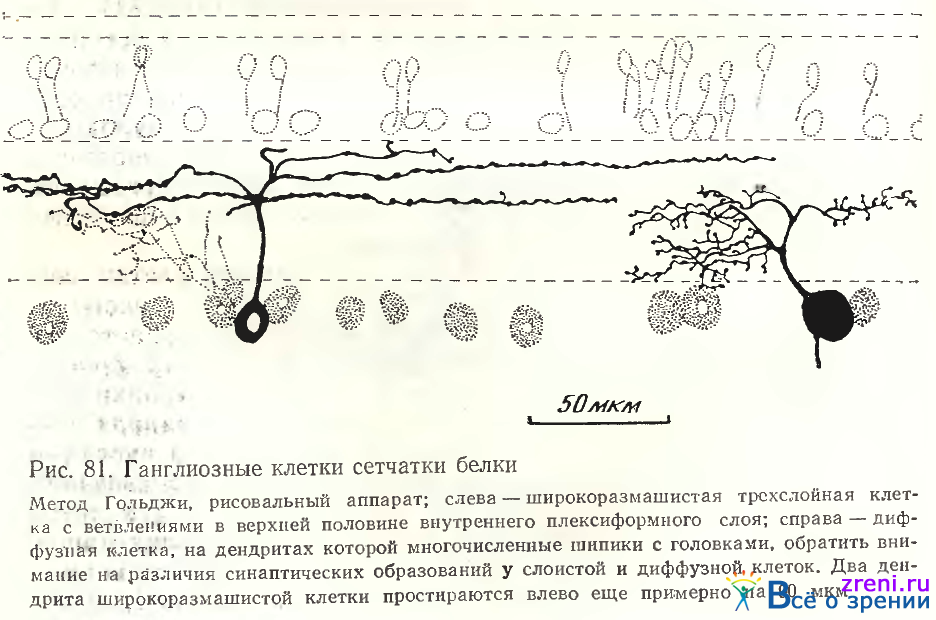
От тела среднего размера прослеживается прямо кверху основной дендрит, отдающий в наружной трети внутреннего плексиформного слоя не менее 5 вторичных ветвлений, покрытых по ходу крупными овальными варикозностями. У другого трехслойного нейрона со сходным типом отростков, покрытых варикозностями, отличалось их расположение — в средней трети внутреннего плексиформного слоя (рис. 82, А).
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
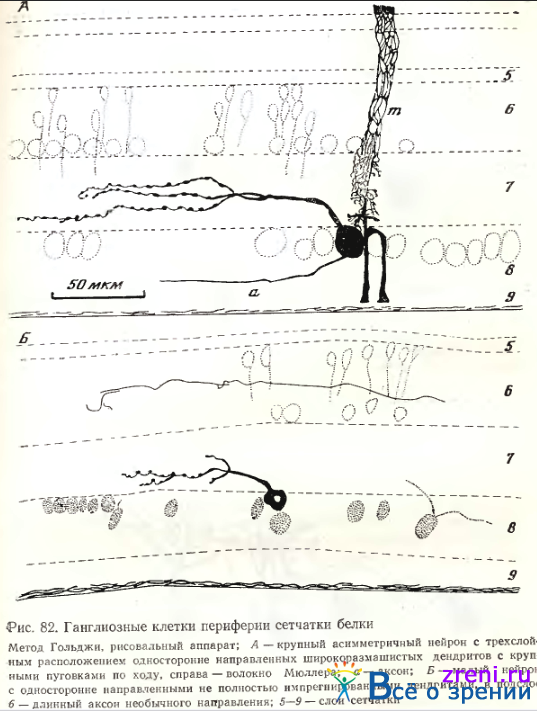
В сетчатке суслика подобных трехслойных нейронов West пе обнаружил.
Весьма разнообразные диффузные ганглиозные клетки сетчатки белки могут быть густоветвистыми и редковетвистыми. У густовствистых основной восходящий дендрит рассыпается на множество многократно делящихся вторичных, весьма извилистых ветвлений, покрытых пуговками и шипиками. Шипики разной формы и величины оканчиваются пуговками; терминали дендритных ветвлений также оканчиваются пуговками, а иногда - в виде двойных пуговок пли более сложных образований (рис. 83).
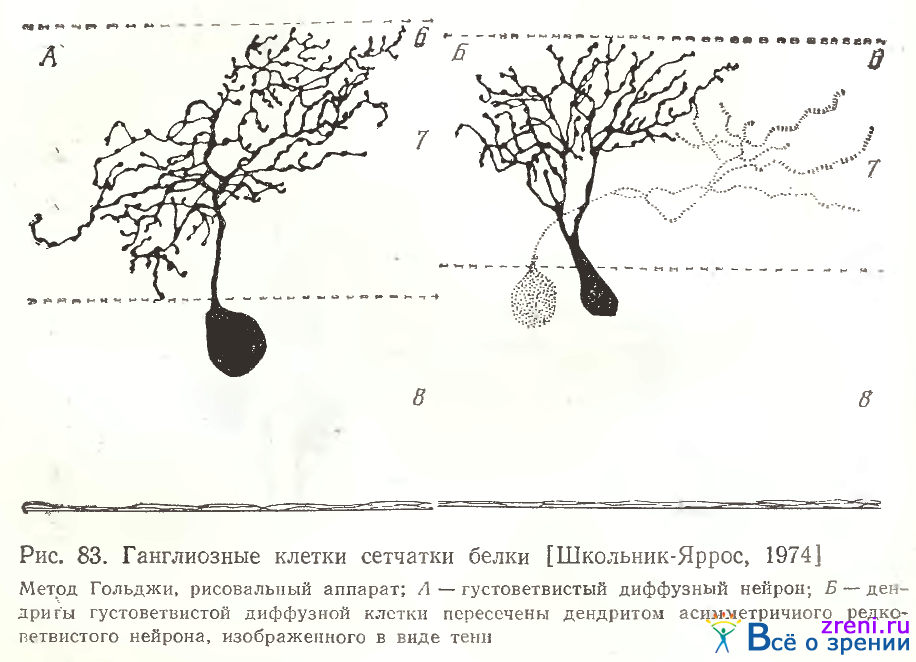
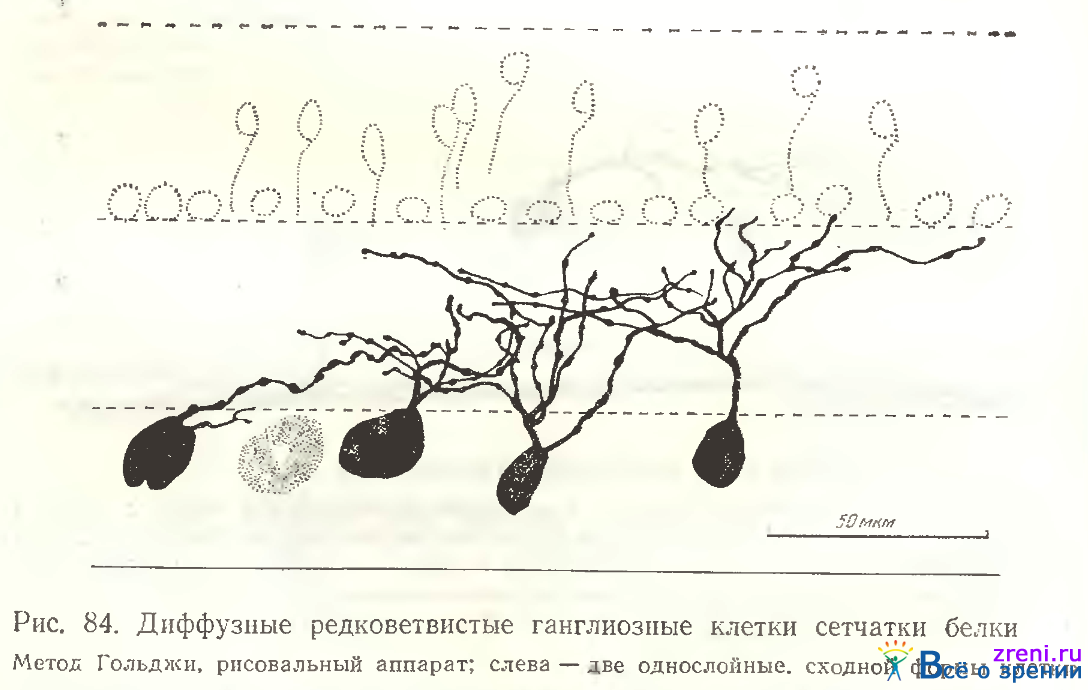
Сходные терминали наблюдались нами у дендритов корковых нейронов и у некоторых их аксонов [Школьник-Яррос, 1965а] Некоторые нейроны характеризуются распространением дендритных отростков по всему плексиформному слою (рис. 83, Л), другие густоветвистые ганглиозные клетки, дендриты которых также извилисты и покрыты синаптическими образованиями, ветвятся лишь в верхней половине внутреннего плексиформного слоя (рис. 83, Б), либо в нижней ее половине (рис. 81, справа). У редковетвистых диффузных ганглиозных клеток дендритные ветвления распространяются по всему плексиформному слою (рис. 84, в центре),
либо в верхних его отделах (рис. 85, Б, в центре).
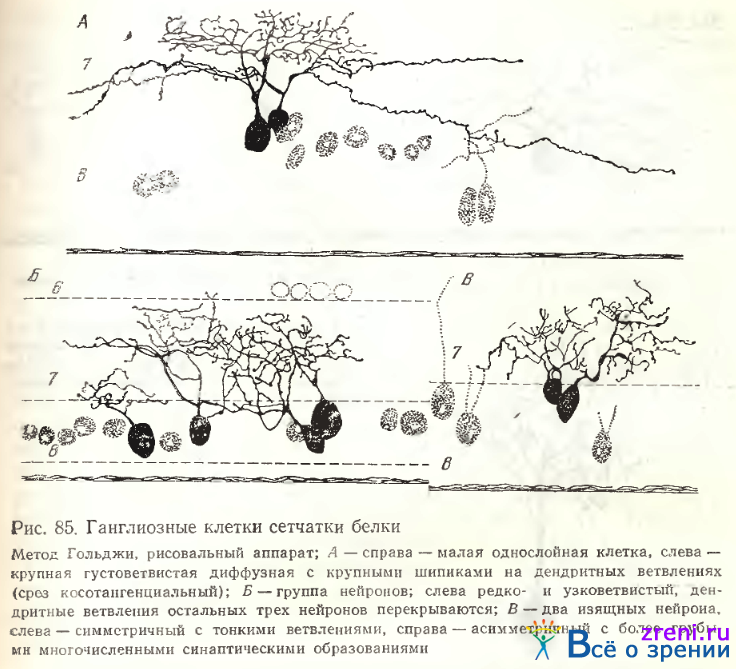
Нежные нейроны с тонкими извилистыми отростками и небольшими пуговками также наблюдались в сетчатке белки (рис. 85, Б; 86, Б).
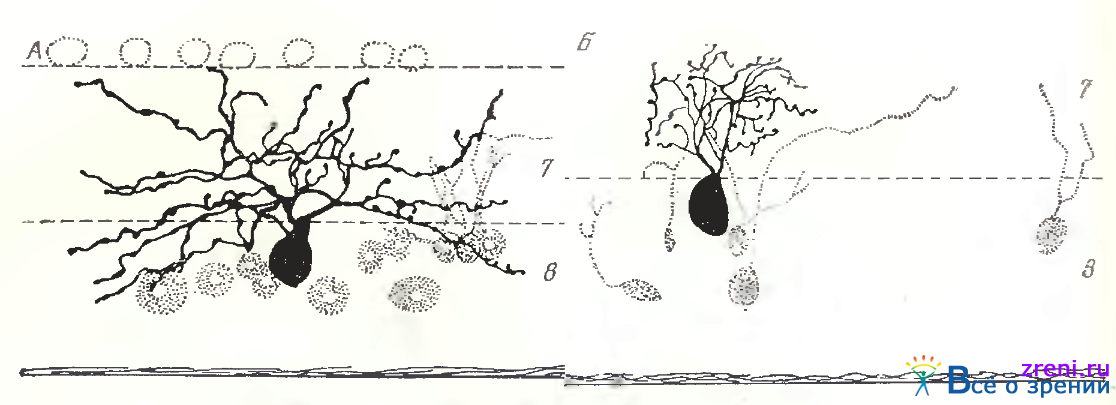
Рис. 86. Две малые диффузные клетки периферии сетчатки белки
Метод Гольджи, рисовальный препарат; А — с распространением извилистых, с синаптическнми пуговками дендритных ветвлений по всему слою 7; Б — узковетвистая с нежными ветвлениями дендритов
Метод Гольджи, рисовальный препарат; А — с распространением извилистых, с синаптическнми пуговками дендритных ветвлений по всему слою 7; Б — узковетвистая с нежными ветвлениями дендритов
Однако подобные нежные клетки импрегнировались значительно реже в сетчатке белки, чем в сетчатке лягушки, где они составляли значительный процент от всех ганглиозных клеток. Все диффузные клетки в сетчатке белки узковетвисты (рис. 87).
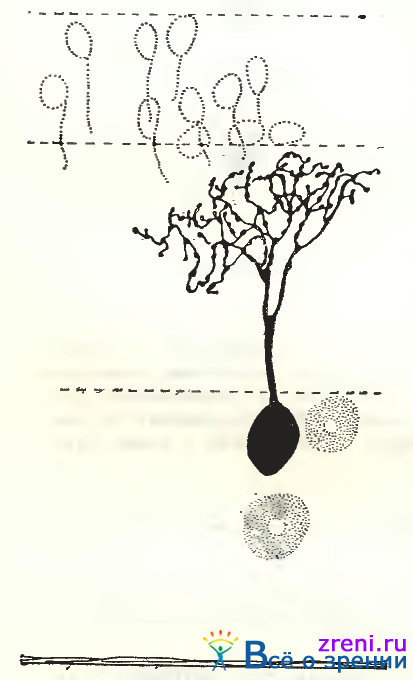
Рис. 87. Диффузная узковетвистая ганглиозная клетка сетчатки белки
Метод Гольджи, рисовальный аппарат; дендриты в верхнем отделе слоя 7
Метод Гольджи, рисовальный аппарат; дендриты в верхнем отделе слоя 7
Нам не удалось импрегнировать широковетвящиеся крупные диффузные клетки, подобные типу 17 в сетчатке лягушки. Та же закономерность отмечается в сетчатке суслика West [1976], где диффузные клетки (G5, G10) узковетвисты. Следует подчеркнуть сходство диффузного нейрона (рис. 86, А) но конфигурации отростков с клеткой [West, 1976, рис. 24] из сетчатки суслика.
Таким образом, при сравнении морфологии ганглиозных клеток сетчатки белки и суслика выявляется их значительное сходство. Так, клетки на наших рисунках 79; 80, А, Б; 77, А, Б, 78; 83, Б, 86, А соответствуют клеткам типов G1, G2, G3, G4, G13, G15, G12, G5, G10 из работы West [1976]. При общем очень большом сходстве можно отметить и некоторые различия. На рисунках West отсутствуют редковетвистые диффузные клетки, представленные на нашем рис. 84. Также нет в сводке West тонко дифференцированных ио подслоям узковетвистых и широковетвистых клеток, подобных изображенным на рис. 81. Однако и в нашем материале отсутствуют клетки, сходные с типами G9, G14, и мало клеток, соответствующих G3 и G11. Отмеченные различия вполне могут объясняться известной капризностью методики Гольджи и при дальнейших исследованиях отсутствующие нейроны могут быть обнаружены.
↑ Сетчатка кошки
За последние годы ганглиозные клетки сетчатки кошки интенсивно изучаются как методом Гольджи, так и другими методами, выявляющими отростки (пероксидазой хрена, внутриклеточными красителями). Несмотря на многие варианты нейронов, обнаруженных Kolb, Nelson, Mariani [1981], основными типами ганглиозных клеток остаются три группы, имеющие четкую функциональную характеристику [Boycott, Wassle, 1974]. Поэтому мы считаем необходимым представить наши данные, полученные ранее методом Гольджи в свете этих основных морфофизиологических фактов. Boycott, Wassle различают три основных типа нейронов: а, b и y. а-клетки характеризуются дендритными полями диаметром от 180 до 1000 мкм; р — от 25 до 300 мкм; у — от 180 до 800 мкм. Размеры дендритных полей клеток а- и b-типов увеличиваются к периферии сетчатки; у y-клеток размеры дендритных полей не меняются, т. е. не зависят от локализации. Предполагается соответствие между морфологическими классами a, b и y физиологическими Y, X и W клетками, так что Х-тип соответствует b, У—а и W—y ганглиозным клеткам. Приводимые далее примеры нейронов [Shkolnik-Yarros,. 1971] будут сопоставлены с классификацией этих авторов (см. также гл. 4). Мы придерживаемся общего деления ганглиозных клеток по Ramon у Cajal на одно-, многослойные и диффузные.
Однослойные клетки чаще всего широкоразмашистые и крупные (рис. 88; 89; 90, А).
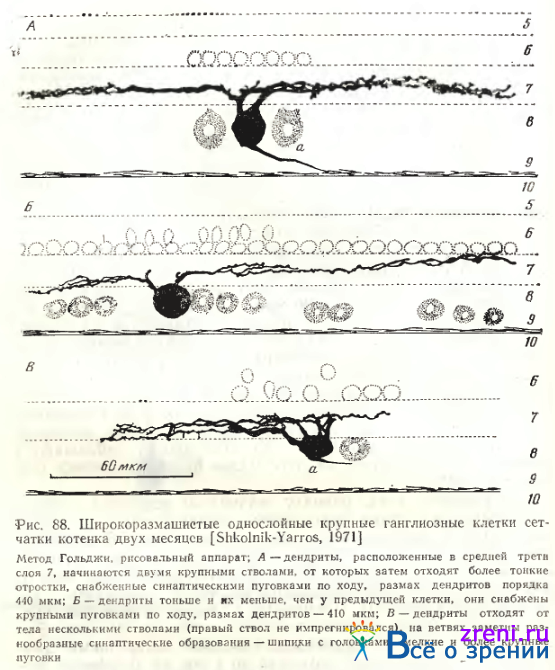
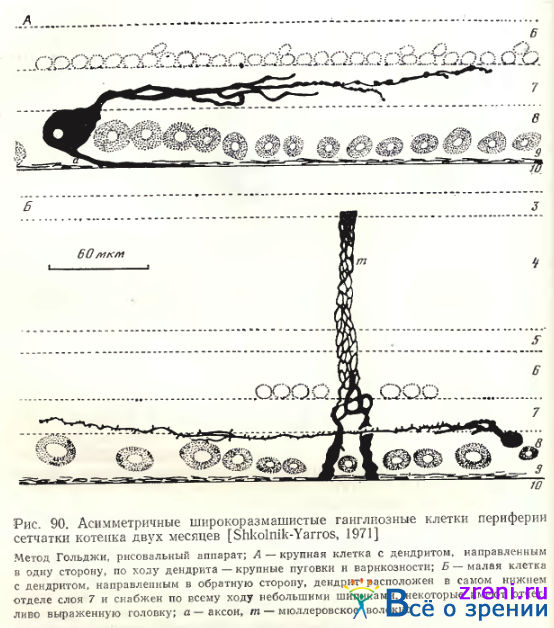
Общее число дендритных ветвлений — от единичных до 20—25. Форма отхождения дендритов также весьма разнообразна: то в виде двух основных крупных стволов (рис. 88, А), то четырех (рис. 88, В); могут наблюдаться и другие варианты (см. рис. 89; 90, А). Размах дендритных ветвлений— примерно от 250 до 600 мкм. Часто можно заметить, что эти даже достаточно полно импрегнированные дендриты оборваны, следовательно, максимальное их распространение может превышать 600 мкм. Расположены дендриты крупных широко-размашистых нейронов преимущественно в среднем отделе внутреннего плексиформного слоя (см. рис. 88; 89; 90, А). Иногда у этой разновидности нейронов дендриты находятся во внутреннем отделе внутреннего плексиформного слоя. По ходу дендритов заметны варикозности (см. рис. 90, А), иногда наблюдаются нежные небольшие отростки, либо малые пуговки и шипики (см. рис. 88, В; 89).
По величине тела, размаху дендритных ветвлений, общей морфологии отростков однослойные крупные широкоразмашистые ганглиозные клетки сетчатки кошки полностью соответствуют типу а по Boycott, Wassle. Приведем основные количественные данные, полученные этими авторами для нейрона типа а. Размеры тел при окраске крезил-виолеттом равнялись 23— 28 мкм в area centralis. Применение Гольджи в быстром варианте давало более крупные величины тела до 50 мкм. Эти данные Boycott, Wassle совпадают с нашими измерениями. Диаметры дендритных полей доходили до 1 мм на периферии сетчатки; основная часть нейронов имела размах дендритов до 250—750 мкм. По подсчетам Wassle, Levick, Cleland [1975] общее количество ганглиозных а-клеток (на препаратах окрашенных крезил-виолеттом) в сетчатке кошки — 6112; пропорция а- клеток составляет 3,3% от всех ганглиозных клеток. Дендритные ветви соседних а-клеток незначительно перекрываются; функционально все а-клетки делятся на on- и оff-клетки, образующие самостоятельные сети. Каждая точка сетчатки покрыта дендритным полем одной on- и одной оff-клетки [Wassle, Peichl, Boycott, 1983].
Однослойными часто являются и широкоразмашистые малые ганглиозные клетки. По размеру тела они относятся к миниатюрным, либо малым (7—13 и 16—20 мкм). Общее количество дендритных ветвлений весьма вариабельно: от 5 до 17 (см. рис. 90, Б; 91).
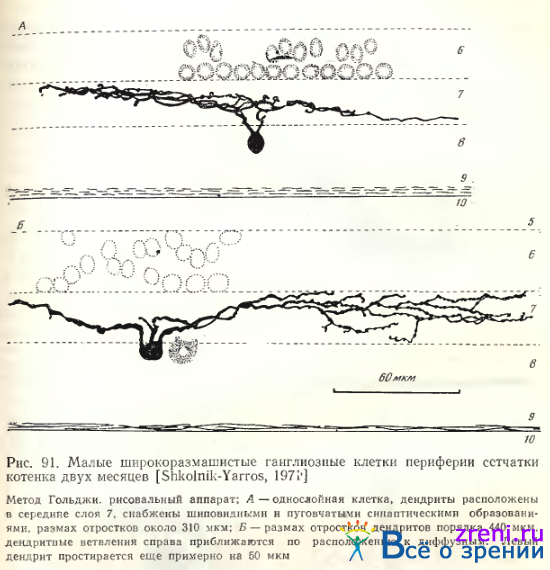
Размах дендритных ветвлений может достигать 400 мкм и более (см. рис. 91), вероятно, эти значения еще больше у взрослого животного. Главная особенность этих нейронов — значительный размах дендритов при малом размере тела. Топография ветвлений значительно отличает эти клетки от предыдущей группы: лишь изредка удается видеть дендриты, расположенные в нижней половине слоя 7 (см. рис. 90, Б), чаще же они находятся в середине или верхней части слоя (см. рис. 91). Синаптические образования на дендритных ветвлениях описываемых клеток отличаются значительным разнообразием. Иногда они имеют вид тонких шипиков с головками (см. рис. 90, Б), наблюдаются и пуговки разных размеров по ходу дендритов (см., рис. 91, Б), некоторые усыпаны мелкими пуговками, другие — крупными, иногда можно наблюдать небольшие варикозности.
По морфологическим признакам: малому размеру тела и большому размаху тонких дендритных ветвлений малые широкоразмашистые ганглиозные клетки сетчатки кошки соответствуют типу y-клеток по Boycott, Wassle. Приводим количественные данные этих авторов: размер дендритных полей — от 180 ДО' 800 мкм, размер тела — от 8 до 18 мкм; аксоны тонкие. Однослойная клетка на рис. 91, А имеет округлое тело, тогда как форма тел на рис. 90, Б и 91, Б — овальная. Возможно, что клетка этого типа (см. рис. 91, А) соответствует d-клеткам [Boycott,. Wassle, 1974], характеризующимся небольшим округлым телом.
Новый вариант ганглиозных клеток в сетчатке кошки обнаружен Stone, Clarke [1980]. Тело у них среднего размера — 16— 25 мкм, а дендритные поля — от 90 до 220 мкм. Авторы считают,, что эти нейроны по характеру ветвления дендритов, а также некоторым физиологическим показателям относятся к типу у.
Однослойные узко- и густоветвистые кустовидные клетки. Величина тела этих нейронов колеблется от малых до средних размеров (см. рис. 89, слева; 92, А, Б).
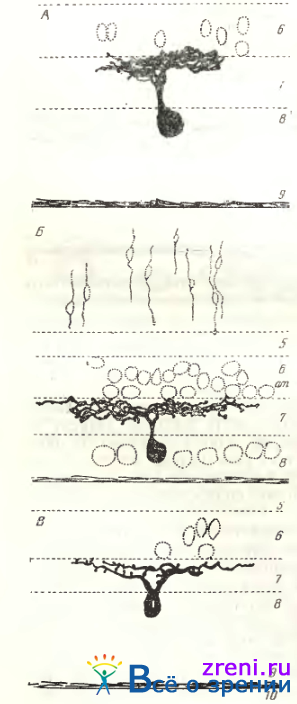
Рис. 92. Кустовидные ганглиозные клетки сетчатки котенка двух месяцев (Б, В — Shkolnik-Yarros, [1971])
Метод Гольджи, рисовальный аппарат; Л.
Метод Гольджи, рисовальный аппарат; Л.
Общее количество дендритных ветвлений — от 22 до 35; размах их также различен — от 90" до 190 мкм. Особенно существенно то, что внутри этой группы нейронов как глубина расположения отростков, так и синаптические образования чрезвычайно вариабельны [Shkolnik-Yarross. 1971]. Дендриты некоторых клеток находятся в нижних отделах слоя 7 (см. рис. 89, слева), у других нейронов они расположены в верхней половине этого слоя (см. рис. 92, А), иногда наблюдаются клетки с деидритами в средней трети слоя. Синаптические образования также весьма разнообразны: то крупные пуговки по ходу дендритов, то много мелких пуговок, число которых доходит до нескольких сот (что соответствует тысячам синапсов при электронно-микроскопическом изучении), то очень тонкие короткие отросточки, заканчивающиеся синаптическими пуговками.
Тип ганглиозной клетки, обозначенный Boycott, Wassle, имеет, по их данным, следующие параметры: диаметр дендритного поля — от 20 до 300 мкм, диаметр тела — от 11 до 24 мкм. По приведенным цифрам, а также по общей морфологии отростков (см. рис. 89, слева; 92, А, Б), сходство густоветвистых, узковетвистых (кустовидных) клеток с типом р несомненно. Больший максимальный диаметр дендритного поля b-клеток зависит от локализации нейрона на периферии сетчатки, далеко от центрального поля. Примерно 55% всех ганглиозных клеток сетчатки кошки — (b-клетки (on- и оff)-клетки расположены попарно), дендритные поля соседних b-клеток полностью перекрываются [Wassle, Peichl, Boycott, 1983].
В группе кустовидных густоветвистых b-клeток обнаруживаются нейроны, приспособленные для различных синаптических связей с амакриновыми и биполярными клетками. Это разнообразие зависит, по-видимому, от структуры синаптических обра зований, от размаха дендритных ветвлений, а также от уровня окончания терминалей во внутреннем плексиформном слое. Так, по Nelson et al. [1981], если дендритные ветвления b-клеток сетчатки кошки находятся в подслое «а» внутреннего плексиформного слоя, то 70% их синапсов происходит от терминалей аксонов колбочковых плоских биполярных клеток. Остальные синапсы у такой клетки — от амакриновых клегок. По ходу дендритного ствола синаптических связен с другими биполярными клетками нет. Если же дендритные ветвления b-клетки оканчиваются в подслое «Ь» внутреннего плексиформного слоя, 70% их синапсов образованы инвагинирующими колбочковыми биполярными клетками. Таким образом, плоские и инвагинирующне колбочковые биполярные клетки сетчатки кошки раздельно связаны с ганглиозными клетками и через них со зрительными центрами.
Однослойные узко- и редковетвистые клетки. Величина их тела преимущественно малая, общее количество дендритных ветвлений — от 6 до 20 (см. рис. 92, В, 93, справа).
Размах дендритных ветвлений различен — от 60 до 180 мкм. Как и у клеток предыдущей группы, различна также глубина расположения дендритов в слое 7 и характер синаптических образований. По ходу дендритов видны разных размеров пуговки, у некоторых клеток дендриты снабжены крупными шипиками с пуговками на концах. Нейроны этой разновидности не представлены в работе Boycott, Wassle .[1974]; сходные по размерам тела и размаху дендритов клетки предыдущей группы (см. рис. 89, слева; 92, А, Б) отличаются выраженной густоветвистостыо [Shkolnik-Yarros, 1971], что характеризует и соответствующие им клетки типа р. Вероятно, редковетвистые клетки соответствуют группе свободно, редковетвящихся клеток, выделенной Leicester, Stone [1967], и противопоставленных группе густоветвистых клеток.
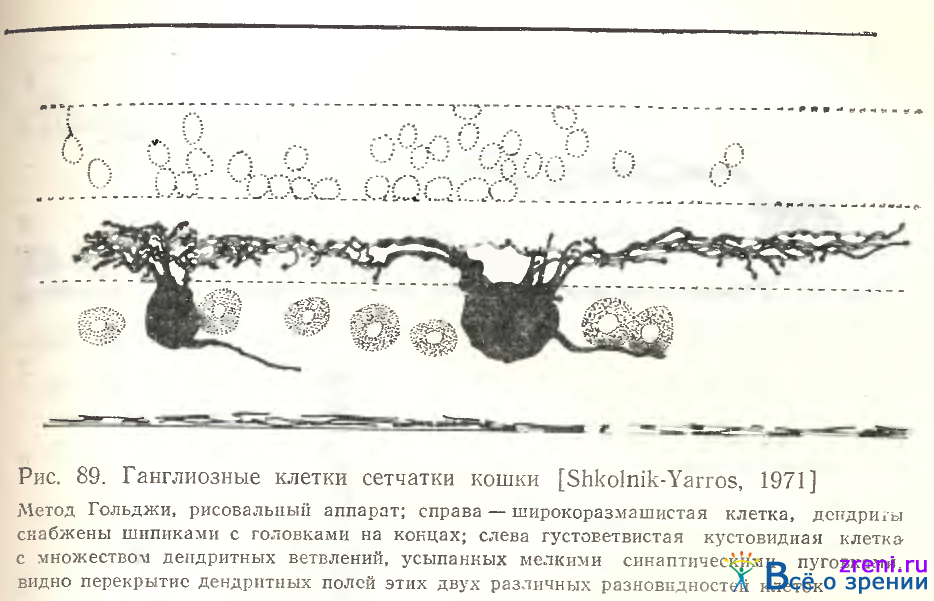
Кроме перечисленных четырех основных типов ганглиозных клеток, в сетчатке кошки наблюдаются реже импрегнирующиеся диффузные клетки (рис. 94).

У представленного на рис. 94 нейрона дендриты отходят от тела тремя тонкими стволиками, тотчас же разветвляющимися на еще более тонкие, усыпанные пуговчатыми образованиями отросточки и шипики. Ветвления распространяются диффузно по внутреннему плексиформному слою, оставляя свободной лишь верхнюю его треть; размах их 95 мкм. По величине тела, очень крупного, и чрезвычайно малому размаху дендритов, не коррелирующему с размерами тела, описываемый нейрон не подходит ни к одному из типов ганглиозных клеток по Leicester, Stone [1967] и Boycott, Wassle [1974].
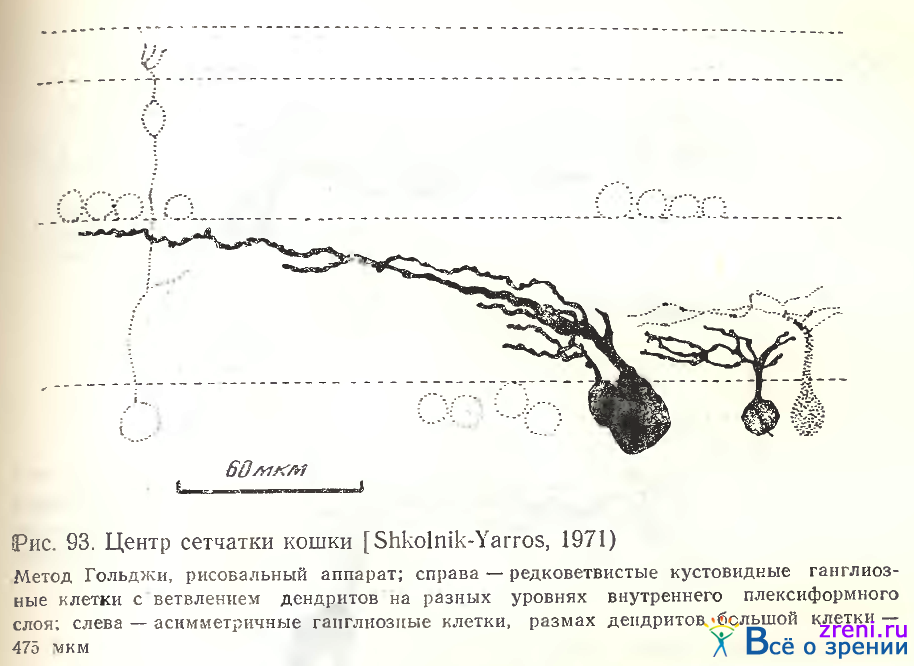
В area centralis наблюдается много плотно упакованных нейронов с небольшим диаметром тела [Stone, 1965] и малым размахом дендритов [Leicester, Stone, 1967]. Однако нейронный состав area centralis не ограничивается совсем малыми и миниатюрными клетками с небольшим размахом дендритов (рис. 95). Рядом находятся клегки со значительно большим размахом ветвлений, дендриты их распространяются на разной высоте слоя 7 (см. рис. 93, справа). Наряду с кустовидными редковетвистыми клетками обнаруживаются и односторонние широкоразмашистые (см. рис. 93, слева). На рис. 93 видно дублирование сходных по форме ганглиозных клеток. Слева дублированы две односторонние клетки, направление их отростков совпадает; справа дублированы две кустовидные редковетвистые клетки с расположением дендритных ветвлений на разных уровнях внутреннего плексиформного слоя. Boycott, Wassle [1974] указывают па наличие в area centralis разных типов ганглиозных клеток; это полностью согласуется с нашими данными [Shkolnik-Yarros, 1971].
Асимметричные ганглиозные клетки у кошки наблюдаются как крупные широкоразмашистые однослойные (см. рис. 90, А), так и малые широкоразмашистые однослойные (см. рис. 90, Б). Соответствие их типам, выделенным Boycott, Wassle, обсуждалось выше. Асимметричные ганглиозные клетки могут быть кустовидными густоветвистыми (рис. 96, Б).
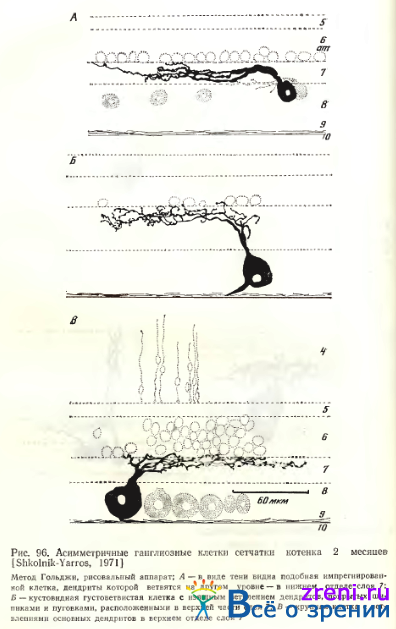
Часто наблюдаются полностью асимметричные нейроны (см. рис. 90, Б; 93; 96, Л).
В сетчатке кошки наблюдались также две небольшие двухслойные ганглиозные клетки [Kolb, Nelson, Mariani, 1981]. Особенности ветвления дендритов ганглиозных клеток сетчатки, их извитость, густота хорошо выявлены исследователями на тангенциальных и косых срезах (рис. 97).
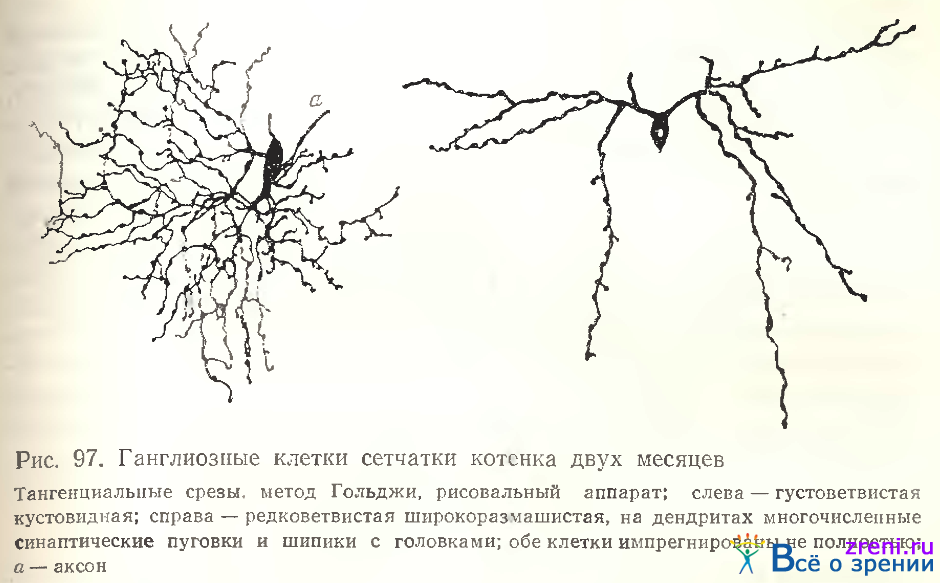
Предложенная ими классификация ганглиозных клеток сетчатки кошки основана также на учете только уровня ветвления дендритов во внутреннем плексиформном слое [Nelson, Famiglietti, Kolb, 1978] (см. гл. 4).
↑ Сетчатка приматов
Ramon у Cajal не изучал сетчатку приматов. Детальные исследования ганглиозных клеток сетчатки приматов принадлежат Dоgiel [1891], Polyak [1941], Van Buren [1963], Boycott, Dowling [1969], С. А. Шибковой [1972], С. А. Шибковой и В. М. Можайцеву [1975], Bunt et al. [1975].
При описании нейронов сетчатки низших, высших обезьян н человека Polyak [1941] ввел оригинальную номенклатуру, обозначив их описательными терминами. Polyak [1941] впервые досконально изучил карликовые ганглиозные клетки, связанные с карликовыми биполярами (см. рис. 22) н через них с колбочками. Boycott, Dowling [1969] отказались от описательных терминов Polyak и вернулись к более обобщенной классификации Ramon у Cajal. Авторы обнаружили следующие типы ганглиозных клеток сетчатки приматов: однослойные, двухслойные, диффузные, диффузно-стратифицированные, карликовые (см. рис. 43).
Новый тип ганглиозной клетки, названной бпплексиформной, в сетчатке макака методом Гольджи обнаружен Mariani [1982а]. Не отличается от обычных тело клетки (диаметр 9 мкм), расположенное в слое 8, и тонкий аксон, прослеживающийся в слое 9. Во внутренней половине внутреннего плексиформного слоя ветвятся 3—4 основных дендрита, дающие вторичные и третичные ветви. В отличие от обычной морфологии от одного из основных дендритов отходит тонкий отросток, идущий к наружному плексиформному слою.
Использованная автором в дополнение, к методу Гольджи Гольджи-электронная микроскопия позволила выявить совершенно новый тип связи: тонкий дендритный отросток оканчивается, образуя центральный элемент ленточного комплекса палочки, подобно синапсам дендрита палочкового бнполяра. Во внутреннем плексиформиом слое связи дендритов тоже отличаются от обычных, они являются постсинаптическнми по отношению к терминалям аксонов биполяров и к неленточным синапсам по ходу аксонов. Mariani видел 19 таких клеток в 8 сетчатках обезьян Macaea mulatta. Внутриклеточные метки пероксидазой хрена подтвердили наличие биплексиформных клеток в сетчатке макак [Zrenner, Nelson, Mariani, 1983].
De Monasterio, Gouras 11975 выделили 6 физиологических типов ганглиозных клеток в сетчатке приматов; из них класс I, типы 1, 2, характеризующиеся тоническими цветооппонентными реакциями, вероятно, соответствуют карликовым ганглиозным клеткам. Они сходны с Х-клетками сетчатки кошки. У клеток класса II, типов 3, 4 — реакции физические, они более характерны для периферической сетчатки и сходны с Y-клетками сетчатки кошки, имеющими наибольший диаметр аксона.
Авторы предполагают, что класс III, типы 5 и 6, сходны с W-клетками сетчатки кошки. О возможной гомологии между Х-клетками сетчатки кошки и ганглиозными клетками сетчатки обезьяны, проецирующимися в мелкоклеточные слон наружного коленчатого тела, а также Y-клетками сетчатки кошки и ганглиозными клетками сетчатки обезьяны, проецирующимися в крупноклеточные слои наружного коленчатого тела, упоминают Rodieck, Brening [1983]. В постедней работе, однако, Rodieck, Binmoeller, Dineen [1985] на основании морфологического исследования методом Гольджи сетчатки человека приходят к выводу, что среди ганглиозных клеток нет аналога а-клеткам сетчатки кошки соответствующих физиологически Y-клеткам.
Современные методы исследования с применением пероксидазы хрена для прослеживания хода аксонов позволили уточнить связи крупных, средних и малых клеток сетчатки обезьян со зрительными подкорковыми центрами [Bunt et al., 1975]. Показана связь всех нейронов определенной ограниченной зоны сетчатки с соответствующей ей зоной мелкоклеточных слоев наружного коленчатого тела. С крупноклеточными слоями связаны только самые крупные клетки периферической сетчатки и примерно 26% клеток parafovea. С передним двухолмием связаны клетки всех размеров, редко разбросанные по сетчатке, но полностью отсутствуют связи с ганглиозными клетками центра сетчатки.
Сравнение морфологии ганглиозных клеток у различных видов позвоночных приводит к предварительному выводу о наличии многослойных нейронов преимущественно в сетчатке низших позвоночных. Однако без учета экологических особенностей того или иного вида можно слишком упростить проблему (см. гл. 5). Так, совершенно ясно, что у ночных либо дневных приматов, у ночных и дневных птиц морфология ганглиозных клеток будет существенно различаться.
----
Статья из книги: Нейроны сетчатки | Е.Г. Школьник-Яррос, А.В. Калинина

Комментариев 0