Ассиметричные нейроны сетчатки


Описание
Асимметрия в строении нервных клеток сетчатки привлекает в последнее время особое внимание. Ранее ганглиозные клетки сетчатки обычно представлялись в виде нейронов с радиально распространяющимися дендрлтами, в центре которых расположено тело (на тангенциальных срезах и тотальных препаратах сетчатки). В исследовании Brown, Major [1966], посвященном описанию нейронов сетчатки кошки, радиальность в распределении дендритов специально подчеркивается. Однако в классических трудах А. С. Догеля [Dogiel, 1891] некоторые нейроны, прекрасно окрашенные метиленовой еннью, совершенно асимметричны. На одном из рисунков Ramon у Cajal [1911] можно также видеть асимметрию в расположении дендритов ганглиозной клетки в сетчатке костистой рыбы, но в тексте этот признак не обсуждается.Позже зарисовки асимметричных разветвлений дендритов ганглиозных клеток сетчатки различных позвоночных животных встречаются в ряде работ [Roscmeyer, Stolte, 1930; Leicester, Stone, 197; Шибкова, 1969, 1971a, 6; Stell, Witkowsky,. 1973; и др.].
Авторы указанных работ не обсуждали этой особенности строения ганглиозных клеток.
В наших работах [Школьник-Яррос, 1971а; Shkolnik-Yarros,. 1971; Kalinina, 1974], кроме выделения специальной разновидности ганглиозной клетки с полностью асимметричными ветвлениями дендритов (тип 6 в сетчатке лягушки, соответствующий типу II, по А. В. Калининой), отмечена асимметрия ветвлений и у других типов клеток и сделана попытка трактовки функционального значения таких нейронов.
Асимметрия в строении дендритного дерева ганглиозных клеток сетчатки отмечена у многих представителей позвоночных: у карпа [Murakami, Shimoda, 1977], у лягушки [Kalinina, 1974], у щуки и лягушки [Школьник-Яррос, Подугольникова, Дюбина, 1975], у белки [Школьник-Яррос, 1974], у кошки [Shkolnik- Yarros, 1971], у лошади и коровы [Шибкова, Хусайнова, 1976], у человека [Шибкова, Можайцев, 1975]. Большинство авторов не выделяют асимметричные нейроны как самостоятельный тип клеток, а относят их к вариантам основной классификации [Boycott, Wassle, 1974; Kalinina, 1974; Мантейфель, Школьник-Яррос, 1980]. При этом симметрия в структуре ганглиозных клеток оценивалась качественно либо по степени отклонения дендритов сл плоскости, перпендикулярной к слоям сетчатки, проходящей через клетку (на радиальных срезах сетчатки), либо по распространению проекции дендритов на плоскость сетчатки (на тотальных препаратах). Количественного анализа этой особенности строения нейронов ранее не проводили.
Нами исследованы с точки зрения симметрии разветвления дендритов средних и крупных нейронов ганглиозного слоя сетчатки озерной лягушки [Жуков, Калинина, 1985; Калинина, Жуков, 1985]. Зарисовки нейронов выполнены с помощью рисовального аппарата с препаратов тотальной сетчатки, импрегнированной серебром по Грос-Бильшовскому—Лаврентьеву. В качестве элементов симметрии выбраны ядрышко нейрона и ось, проходящая через ядрышко в направлении слепого пятна, характеризующие центральную и двустороннюю симметрию соответственно (рис. 98).
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
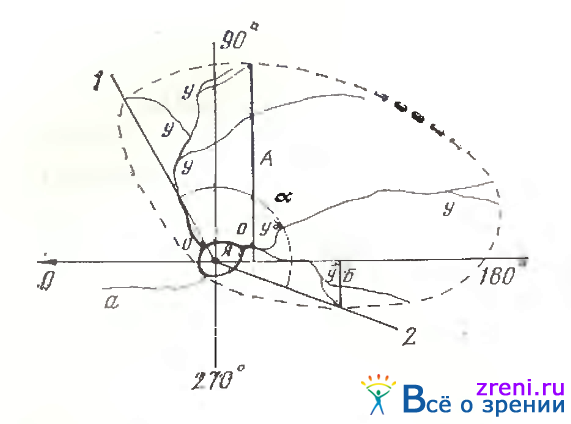
Рис. 98. Параметры структуры крупного нейрона сетчатки лягушки [Жуков, Калинина, 1985]
Тотальный препарат сетчатки, импрегнация серебром по Грос-Бильшовскому—Лаврентьеву, рисовальный аппарат; 0—180° — ось симметрии, проведенная через ядрышко (я) к слепому пятнуу (стрелка); а—аксон; о — основной дендрит; у — узел ветвления дендрита; отрезки А, Б — кратчайшее расстояние максимально удаленных терминалей дендритов до оси симметрии (0—180°). Плавная пунктирная кривая ограничивает дендритное поле нейрона. Угол а образован лучами 1 и 2,проведенными из ядрышка к краевым терминалям дендритов
Тотальный препарат сетчатки, импрегнация серебром по Грос-Бильшовскому—Лаврентьеву, рисовальный аппарат; 0—180° — ось симметрии, проведенная через ядрышко (я) к слепому пятнуу (стрелка); а—аксон; о — основной дендрит; у — узел ветвления дендрита; отрезки А, Б — кратчайшее расстояние максимально удаленных терминалей дендритов до оси симметрии (0—180°). Плавная пунктирная кривая ограничивает дендритное поле нейрона. Угол а образован лучами 1 и 2,проведенными из ядрышка к краевым терминалям дендритов
Анализ расположения дендритов относительно тела клетки (ядрышка) показал, что центральной (радиальной) симметрией не обладает ни один из исследованных нейронов. Отсутствие центральной симметрии удобно характеризовать величиной угла а, образованного лучами 1 и 2, проведенными из ядрышка к краевым терминалям дендритного поля, (рис. 98). Очевидно, что у таких нейронов угол ветвления много меньше 360°, а тело клетки не лежит в центре дендритного поля.
На рис. 99

Рис. 99. Величина угла ветвления дендритов (а) нейронов разных типов: II (6), III {13—16), IV (2—5) и V (17) ганглиозного слоя сетчатки озерной лягушки [Жуков, Калинина, 1985]
Четыре колонки от 0 до 360° — четыре сектора прямоугольной системы координат с центром в ядрышке нейрона (см. рис. 98). Объяснения в тексте.
Четыре колонки от 0 до 360° — четыре сектора прямоугольной системы координат с центром в ядрышке нейрона (см. рис. 98). Объяснения в тексте.
представлены углы ветвления дендритов нейронов четырех типов. Нумерация типов ганглиозных клеток, по классификации А. В. Калининой [Kalinina, 1974], обозначена римскими цифрами, соответствующие им типы предлагаемой в настоящей работе классификации — арабскими цифрами. Видно, что дендриты ганглиозных клеток типа II (6) ветвятся в секторе, как правило, меньшем 180°. Дендриты крупных ганглиозных клеток (III—IV типов) занимают больший сектор. Величина угла а в целом возрастает от ганглиозных клеток типа II (6) к типу IV (2—5). Лишь у одного нейрона, представленного на рисунке (тип III), угол равен 360°, однако этот случай не может служить примером центральной симметрии, так как дендриты ветвятся неравномерно, а тело нейрона не лежит симметрично по отношению к дендритному полю. Последнее свойство характерно для ганглиозных клеток всех типов, кроме некоторых нейронов IV (2—5) типа (см. ниже).
Ориентация измеренных у 160 крупных нейронов ганглиозного слоя сетчатки лягушки углов а на плоскости сетчатки по отношению к естественным зрительным координатам показан на рис. 100.
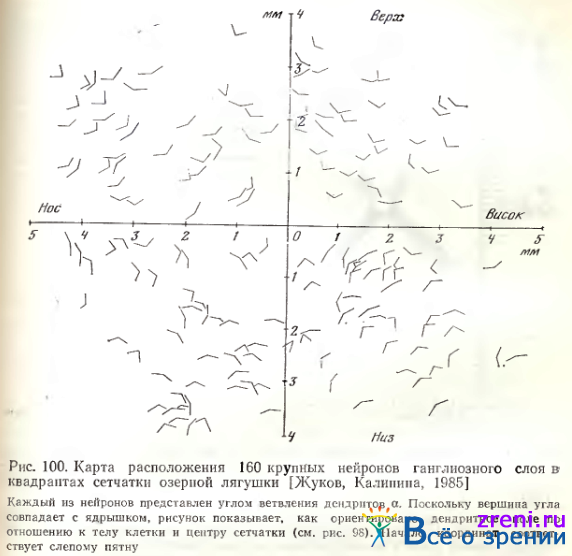
Карта составлена таким образом, что местоположение каждого нейрона, величина и ориентация угла а даны в прямоугольной системе координат с центром в слепом пятне. Очевидно, что у подавляющего большинства ганглиозных клеток во всех квадрантах сетчатки вершина угла ветвления дендритов ориентирована в сторону слепого пятна. Поскольку у каждой ганглиозной клетки в направлении слепого пятна отходит аксон, а вершина угла а совмещена с ядрышком (следовательно, дендриты и аксон находятся на противоположных полюсах нейрона), дендриты ветвятся преимущественно в направлении от центра сетчатки к ее краям. Аналогичная закономерность установлена и для нейронов типа II (6).
Эта закономерность дает основание в качестве элемента симметрии выбрать естественную ось, проходящую через ядрышко в направлении слепого пятна (0—180° на рис. 98), и оценить симметрию дендритного поля по отношению к этой оси. Для этой цели мы использовали некоторые структурные признаки.Один из них — кратчайшее расстояние до оси наиболее удаленных терминалей дендритов (А и Б на рис. 98).
На рис. 101
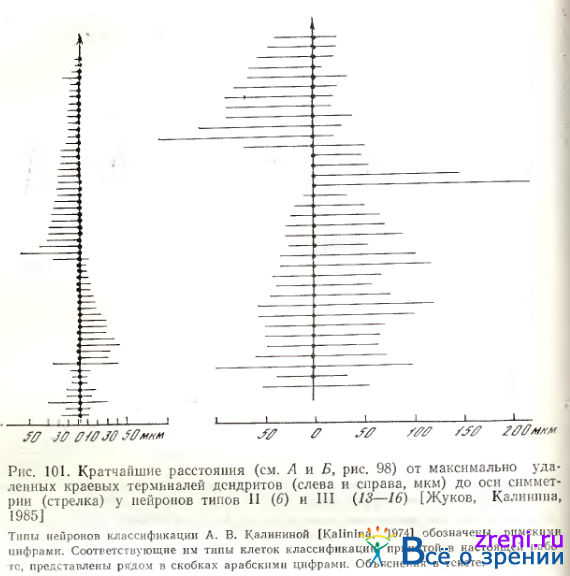
представлены результаты измерений Ли Б для ганглиозных клеток типов II (6) и III (13—16). Видно, что у нескольких клеток (внизу) расстояния справа и слева от оси одинаковы, однако у большинства ганглиозных клеток они существенно отличаются и, следовательно, дендритные паля асимметричны по этому признаку. Среди ганглиозных клеток типа II (6) преобладают нейроны, у которых дендриты располагаются лишь с одной стороны от оси симметрии (слева или справа), при этом удаленность от оси терминалей дендритов у них сравнительно невелика: 10—40 мкм (у крупных нейронов до 500 мкм). У нейронов типов IV (2—5) или V (17) ассимметрия выражена в меньшей степени, чем у нейронов типов II (6) и III (13—16). Например, из 13 ганглиозных клеток типа IV (2—5) у семи терминали дендритов слева и справа от оси удалены одинаково. Вытянутые дендритные поля этих нейронов своей длинной осью ориентированы перпендикулярно оси симметрии. Тело таких клеток лежит симметрично в дендритном поле.
Другой признак — число узлов ветвления, характеризующий степень разветвленное, сложности дендритного дерева. Подсчет числа узлов ветвления слева и справа от оси симметрии показал, что большинство крупных ганглиозных клеток асимметричны, двусторонней симметрией обладают менее четверти исследованных клеток. Общее число узлов ветвления у крупных диффузных нейронов не превышает десяти (среднее 4). Напомним, что у ганглиозных клеток типа II (6) максимальное число узлов ветвления два. Они также не имеют двусторонней симметрии поданному признаку. Третий признак — число основных (отходящих от тела клетки) дендритов — может характеризовать не только «мощность» дендритного дерева, но и направленность основных дендритов, поскольку подсчет проводили в
4 секторах, образованных двумя взаимноперпендикулярными осями. Суммарные диаграммы, представленные на рис. 102
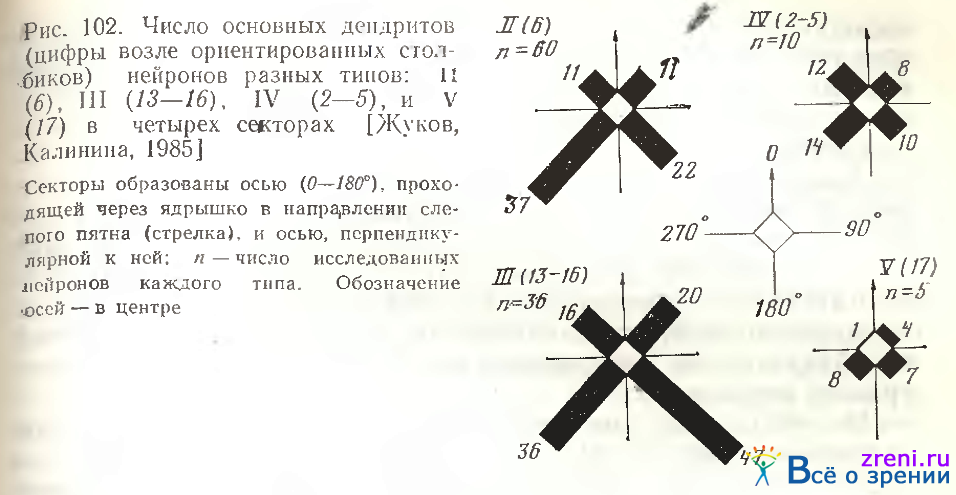
показывают, что у ганглиозных клеток типов II (6), III (13—16),(17) дендриты отходят в основном в смежных секторах 90— 180° и 180—270°, т. е. в противоположном направлении от слепого пятна, что отражает данные, представленные на рис. 100 и наши ранние наблюдения [Kalinina, 1974]. У нейронов этих типов асимметрия относительно оси 0—180° выражена в меньшей степени, чем по отношению к оси, перпендикулярной к ней. Нейроны типа IV (2—5) по этому признаку симметричны, так как числа основных дендритов во всех секторах мало отличаются друг от друга.
Использование тотальных препаратов сетчатки дает возможность при оценке симметрии в строении нейронов строго определить элементы симметрии. Мы выбрали два элемента и несколько структурных признаков, оказавшихся достаточными для характеристики симметрии в общей картине ветвлений дендритов. Установлено, что ганглиозные клетки сетчатки лягушки, относящиеся к средним и крупным, радиальной симметрией не обладают. В противоположность этому у некоторых млекопитающих, как уже указывалось выше, наблюдается преимущественно радиально симметричное ветвление дендритов ганглиозных клеток.
Анализ структурных признаков выявил отсутствие и двусторонней симметрии у большинства исследованных клеток. Ранее было показано, что асимметрия свойственна также мелким ганглиозным клеткам [Школьник-Яррос, 1971]. Таким образом, асимметричное строение ганглиозных клеток у лягушки является скорее нормой, чем исключением. Асимметричными отростками обладают, как уже указывалось, и нейроны предшествующих уровней сетчатки.
Известно, что рецептивные поля сетчатки проявляют асимметричные свойства. У лягушки функциональная асимметрия: рецептивных полей ганглиозных клеток выражается в дирекциональной избирательности [Morrison, 1975; Backstrom, Hcmila,. Reuter, 1978], а также в ряде других характеристик, например в дирекционально зависимом изменении времени возникновения пиков ответов [Жуков, 1980; Hodos, Dowcs, Keating, 1982}. В последнее время предпринимаются попытки рассматривать асимметричное ветвление дендритов нейронов сетчатки как морфологическую основу детектирования некоторых параметров зрительного раздражителя, в том числе и направления движения [Школьник-Яррос, 1971; Маитейфель, Школьник-Яррос, 1980; Naka, 1980; Mariani, 1982b, 1983]. Однако эти попытки пока не выходят из рамок гипотезы. Согласно модельным представлениям, нейроны с односторонне направленным дендритом (такие нейроны у лягушки обычны среди ганглиозных клеток типа 6 (II)) способны детектировать направление движения стимула [Беркинблит и др., 1971].
Если принять упомянутую гипотезу, то найти в сетчатке лягушки асимметричные нейроны электрофизиологически не представляло бы труда. Между тем в крыше среднего МОЗГУ, например, куда проецируется большинство ретинальных волокон, дирекционально избирательные ганглиозные клетки регистрируются чрезвычайно редко [Grusser, Grusser-Cornehls, 1976]. Получены и экспериментальные данные, противоречащие этой гипотезе, которые, правда, следует считать предварительными. Так, в работе, выполненной на сетчатке кролика [Amthor, Oyster, Takahashi, 1985], были внутриклеточно окрашены с помощью пероксидазы хрена 14 дирекционально избирательных ганглиозных клеток и изучена морфология их дендритов. Авторы не нашли очевидной взаимосвязи между предпочитаемым клеткой направлением движения стимула и асимметрией в общем рисунке дендритных разветвлений. В то же время они обнаружили такие детали морфологии дендритов, которые характерны, по их мнению, исключительно для ганглиозных клеток с дирекциональными свойствами: двухслойное строение, наличие шипиков, дендриты малого диаметра, образующие замкнутые петли. Исследованные нами на тотальных препаратах сетчатки диффузные крупные клетки не обладают набором таких признаков, он характерен для дендритов мелких ганглиозных клеток сетчатки лягушки [Мантейфель, Школьник-Яррос, 1980]. Интересно в связи с этим, что у лягушки дирекцпональная избирательность ганглиозных клеток, имеющих большие рецептивные поля, обнаруживается значительно реже, чем у клеток с полями небольшого размера [Morrison, 1975; Backstrom, Hcmila, Reuter, 1978]
В задаче поиска корреляции между структурой дендритов и функциональными свойствами нейронов сетчатки необходимо корректное описание геометрии дендритного поля. При этом расположение отростков и топография нейрона в целом должны рассматриваться в единой для всей выборки клеток системе координат. Предложенный в настоящем разделе работы метод, по нашему мнению, отвечает этим требованиям.
----
Статья из книги: Нейроны сетчатки | Е.Г. Школьник-Яррос, А.В. Калинина

Комментариев 0