Внутрицитоплазматические включения


Содержание:
Описание
Гликоген. В цитоплазме многих клеток обнаруживаются неправильной формы частицы диаметром 20—30 нм. Различают два типа гликогена. Первый тип характеризуется скоплением изолированных частиц, равномерно или неравномерно распределенных в цитоплазме (бета-частицы). Помимо бета-частиц можно обнаружить и альфа-частицы. Отличия сводятся к тому, что в альфа-частицах отдельные гранулы собираются в розетки. Наиболее часто гликоген обнаруживается в клетках печени и мышечной ткани.Липиды. В норме в цитоплазме клеток обнаруживаются капельки липидов. Депозиты липидов имеют различный вид. Часть капелек гомогенного вида, в то время как другие обладают пластинчатой структурой. Отличаются они и плотностью. Большинство свободных липидов исчезает в процессе гистологической обработки материала, поскольку вымываются спиртами и ксилолом.
Пигментные гранулы. В тканях глаза можно обнаружить два типа пигментных гранул. Это меланин и липофусцин.
Меланин широко представлен в тканях глаза, выполняя довольно важные функции в фоторецепции. Меланиновые гранулы обнаруживаются как в нейроэпителиальных производных, таких как пигментный эпителий сетчатки, радужки, ресничного тела, так и в стромальных меланоцитах увеального тракта. Пигментные гранулы перечисленных двух оболочек довольно трудно различать при световой микроскопии. Гранулы, выявляемые в нейроэпителии, несколько больше в размерах и темнее, чем обнаруживаемые в увеальном тракте. Второй тип гранул — липофусцин — может быть спутан с меланином, поскольку он имеет светло-коричневый цвет и по размерам приближается к зернам меланина (рис. 1.1.26).

Рис. 1.1.26. Зерна липофусцина (стрелки) в цитоплазме нейрона (трансмиссионная электронная микроскопия)
Гранулы липофусцина имеют более светлый цвет. Липофусцин чаще обнаруживается в пожилом и старческом возрасте, особенно в клетках миокарда и нейронах. Довольно много появляется его с возрастом в клетках пигментного эпителия сетчатки.
В клетках нередко обнаруживаются и другие пигменты, такие как гемоглобин, билирубин. Продукты деградации гемоглобина, депозиты железа чаще обнаруживаются в патологически измененных тканях глаза (стекловидное тело, увеальный тракт).
↑ Интерфазное ядро
Все клетки содержат ядра, форма и размеры которых могут быть самыми разнообразными. В настоящем разделе мы остановимся на морфологии интерфазного ядра, т. е. ядра, находящегося вне митотического цикла. В тканях подавляющее большинство ядер находится в интерфазе.
В ядре четко выявляются следующие структуры (рис. 1.1.27):
- Ядерная оболочка.
- Хроматин.
- Ядрышко.
- Ядерный сок.
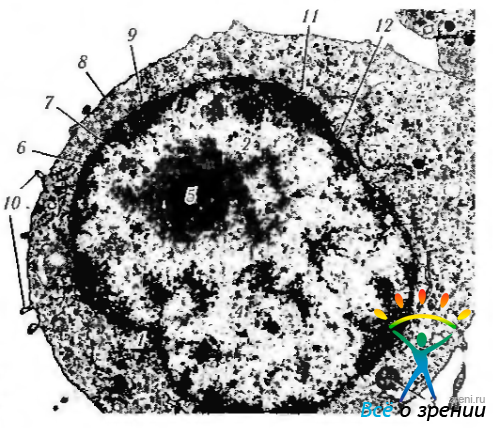
Рис. 1.1.27. Ультраструктурные особенности лимфоцита периферической крови. Четко определяется структура ядра: 1— эухроматин; 2— гетерохроматин; 3 — митохондрии; 4 — ядро; 5 — ядрышко; 6 — ядерная оболочка; 7 — ядерная пора; 8 — плазмолемма; 9 — перинуклеарное пространство; 10— псевдоподии: 11 — полирибосомы; 12 — шероховатый эндоплазматический ретикулум
Ядерная оболочка (кариолемма). Ядерная оболочка окружает ядро и хорошо видна в препаратах. Это связано с тем, что с внутренней стороны к ней прилежит хроматин, интенсивно окрашивающийся гематоксилином. Окрашенный хроматин и контурирует оболочку.
При исследовании в электронном микроскопе оболочка выглядит в виде двух темных мембран, между которыми определяется светлое пространство толщиной 25 нм. Толщина каждой электронноплотной мембраны 8 им.
В ядерной мембране определяются многочисленные «поры» (рис. 1.1.28).
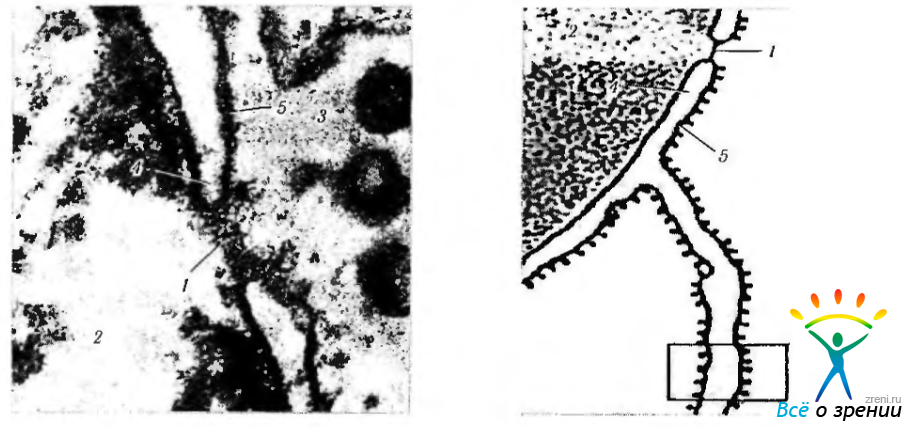
Рис. 1.1.28. Ультраструктурные особенности (а) и схематическое изображение (б) строения поры ядерной оболочки: 1 - пора; 2 — ядро; 3 — цитоплазма клетки; 4 — внутренняя мембрана ядерной оболочки; 5 — наружная мембрана ядерной оболочки; 6 гранулярный эндоплазматический ретикулум
Ядерные поры занимают от 3 до 35% всей поверхности ядра. Именно в этих местах два электронноплотных слоя оболочки как бы сливаются. В области пор обнаруживается скопление хроматина. Ультраструктурные исследования выявили, что в области пор располагаются и довольно сложные структуры, состоящие из канальцев, обращенных как в сторону цитоплазмы, так и внутрь ядра. Этот комплекс структур называют пoровым комплексом (рис. 1.1.29).
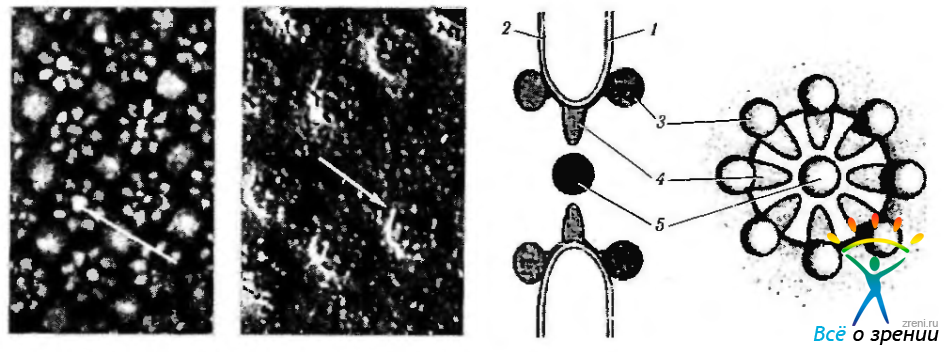
Рис. 1.1.29. Поры ядерной оболочки: а — сканирующая электронная микроскопия (стрелкой указан комплекс пберной поры); б—ядерные поры при применении метода замораживания-скалывания; в — схема организации комплекса ядерной поры (1 — наружная мембрана кариолеммы; 2— внутренняя мембрана кариолеммы; 3— белковые гранулы; 4 — белковые фибриллы; 5 — центральная гранула)
Поровый комплекс содержит два параллельных кольца (по одному с каждой поверхности кариолеммы) диаметром 80 им, которые образованы 8 белковыми гранулами. От этих гранул к центру сходятся фибриллы, формирующие перегородку (диафрагму) толщиной 5 нм. В середине этой мембраны лежит центральная гранула, которая представляет собой субъединицу рибосомы. Поры способствуют обмену метаболитов между ядром и цитоплазмой. Основной функцией ядерных пор является обеспечение регуляции избирательного транспорта веществ между цитоплазмой и ядром, активный перенос в ядро белков, перенос в цитоплазму субъединиц рибосом.
Хроматин при световой микроскопии выглядит в виде глыбок различной степени дисперсности, равномерно или неравномерно распределенных в кариоплазме (рис. 1.1.30).
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
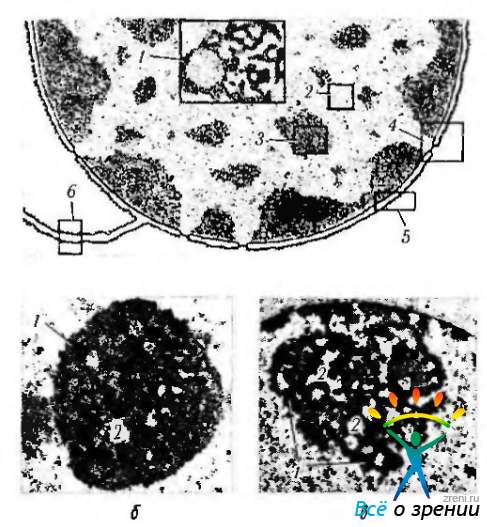
Рис. 1.1.30. Ультраструктурная организация ядра: а — схема структурных компонентов ядра (1 — ядрышко; 2— эухроматин; 3— гетерохроматин; 4 — ядерная пора; 5 — кариолемма; б — цистерна шероховатой эндоплазматической сети); б, в — особенности строения ядрышка (1—темный компонент; 2— светлый компонент)
Отмечается его большая конденсация вблизи ядерной мембраны (периферический хроматин) и вблизи ядрышка (околоядрышковый хроматин). Описанный тип хроматина называется конденсированным, поскольку он виден при световой микроскопии. Но кроме конденсированного гетерохроматина в кариоплазме существует и неконденсированный хроматин, или эухроматнн.
В химическом отношении хроматин представляет собой комплекс ДНК и белка. Этот комплекс соответствует хромосомам, которые в интерфазном ядре представлены длинными, тонкими перекрученными нитями. Они неразличимы как индивидуальные структуры.
Транскрипция информации с молекул ДНК осуществляется только с молекул эухроматина.
Морфологической разновидностью гетерохроматина является тельце Барра, обычно расположенное вблизи ядерной оболочки. Обнаруживаются тельца только у женщин, поскольку представляют собой одну из конденсированных Х-хромосом.
Функция реализации генетической информации в интерфазном ядре осуществляется непрерывно благодаря процессам транскрипции. При транскрипции ДНК образуется одна очень крупная молекула РНК (первичный транскрипт), которая связывается с ядерными белками с образованием рибонуклеопротсидов. В первичном РНК-транскрипте (как и в матричной ДНК) имеются дискретные значащие последовательности нуклеотидов (экзоны), разделенные длинными вставками (нитронами). Процессинг РНК-транскрипта включает отщепление интронов и стыковку экзонов — сплайсинг. При этом очень крупная молекула РНК превращается в достаточно мелкие молекулы информационной РНК, отделяющиеся от связанных с ними белков при переносе в цитоплазму.
Ядрышко (рис. 1.1.30). Ядрышко представляет собой расположенное в ядре плотное образование. Размеры, плотность, форма и локализация ядрышек могут быть самыми разнообразными. Отмечено, что более интенсивна синтетическая деятельность клетки при больших размерах ядрышка. Да это и понятно, поскольку ядрышко обеспечивает синтез РНК.
Ультраструктурные исследования позволили выявить довольно сложную структуру ядрышка. В нем различают
- гранулярный,
- фибриллярный
- и аморфный компоненты.
Гранулярный компонент представлен зернами (диаметр 10—20 нм), состоящими из рибонуклеопротеидных частиц (субъединицы рибосом). Фибриллярная часть состоит из плотных тонких электронноплотных нитей (диаметр 5—8 нм), образующих компактную массу. Эти волокна концентрируются вокруг более светлых сердцевин из менее плотного материала (фибриллярные центры). Считается, что фибриллярный материал представляет собой РНК (рибосомальная РНК), а фибриллярные центры состоят из ДНК и по строению соответствуют зернам хроматина.
Аморфный компонент окрашивается бледно и содержит участки расположения ядрышковых организаторов со специфическими РНК-связывающими белками и крупными петлями ДНК, активно участвующими в транскрипции рибосомальной РНК-Фибриллярный и гранулярный компоненты образуют ядрышковую нить (нуклеонему), толщина которой 60—80 нм.
Ядерный сок (кариоплазма). Ядерный сок представляет коллоидный раствор белка, в котором и располагаются перечисленные структуры. Ядерный сок не окрашивается ядерными красителями.
Основными функциями ядра является хранение генетической информации (в молекулах ДНК, находящихся в хромосомах), реализации генетической информации, контролирующей осуществление различных жизненных функций клетки, воспроизведение и передачу генетической информации. Последняя функция осуществляется благодаря клеточному делению.
↑ Клеточное деление
Задачей этого раздела не является изложение основ генетики. Мы опишем лишь морфологические проявления деления клеток.
Необходимо отметить, что биологический смысл клеточного деления сводится к воспроизведению дочерней клетки, идентичной материнской. В генетическом плане суть деления сводится к тому, что материнская клетка, обладающая определенным набором хромосом (23 пары хромосом у человека), первоначально удваивает генетический материал, и только затем происходит разделение клетки. При этом генетический материал равномерно распределяется между двумя идентичными в генотипическом плане клетками. Это свойство клеток является не только основой возникновения, развития организма, но и поддержания его целостности на протяжении всей жизни. Огромное значение деление клеток имеет и при регенерации поврежденных тканей.
Наиболее распространенной формой самовоспроизведения клетки является митоз, на морфологических проявлениях которого мы и остановимся несколько ниже. Первоначально необходимо охарактеризовать стадию, предшествующую митозу, а именно интерфазу (рис. 1.1.31. 1.1.32).
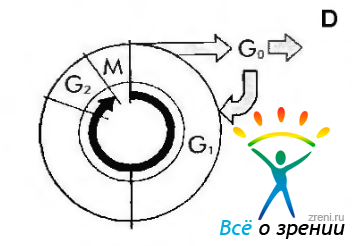
Рис. 1.1.31. Клеточный цикл: G,, G2 и G„— периоды интерфазы; М — митоз; D — гибель клетки
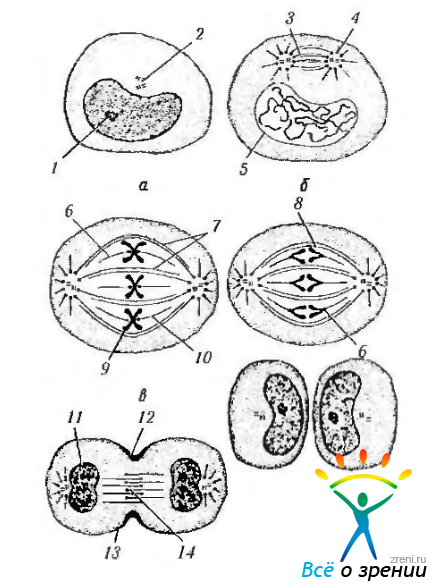
Рис. 1.1.32. Фазы митотического деления (по А. Аэл/, Д.Кормак, 1982): а — интерфаза (G2); б—профаза; в — метафаза; г — анафаза; д — телофаза; е — интерфаза (G,) (1 — ядрышко; 2— центриоли; 3—веретено деления; 4—звезда; 5—ядерная оболочка; 6—кинетохор; 7 непрерывные микротрубочки; 8 s-хромо-сома; 9 — d-хромосома; 10— хромосомные микротрубочки; 11 — формирование ядра; 12— борозда дробления; 13 — пучок актиновых нитей; 14 - остаточное (срединное) тельце)
Интерфаза характеризуется наличием последовательных структурных и биохимических преобразований, подготавливающих клетку к митозу. Весьма важным в интерфазе является матричный синтез ДНК и удвоение хромосом — S-фаза. Промежуток времени между делением и наступлением S-фазы называется фазой G, (постмитотическая или постсинтетичес-кая фаза), а между S-фазой и митозом — фазой G2 (постсинтетическая или предмитотическая фаза). В течение фазы G, клетка диплоидная, в течение фазы S плоидность возрастает до четырех. Таким образом, в фазе G9 клетка уже тетраплоидная.
В интерфазе усиливаются биосинтетические процессы. Удваивается масса клетки, происходит деление центриолей. В течение предмитотической фазы (G2) обе материнские центриоли осуществляют сборку микротрубочек, усиливается формирование лизосом, делятся митохондрии и синтезируются новые белки, необходимые для осуществления митоза. К концу интерфазы хроматин конденсирован, ядрышко хорошо видно, ядерная оболочка не нарушена.
Наиболее важным и сложным процессом, происходящим в интерфазе, является удвоение набора хромосом. Суть удвоения состоит в том, что на цепочке ДНК синтезируется точно такая же параллельная цепочка. Этот процесс называется репликацией. Биологическая суть репликации сводится к тому, что при этом происходит передача генетической информации, хранящейся в родительской ДНК, путем точного ее воспроизведения в дочерней клетке. При этом каждая родительская цепь ДНК является матрицей для синтеза дочерней (матричный синтез ДНК). Процесс репликации довольно сложен. Подробное описание этого процесса можно найти в многочисленных руководствах по генетике.
В конце интерфазы клетка практически подготовлена к митотическому делению, которое в последующем и наступает. Морфологически различают 4 фазы митоза:
- профаза,
- метафаза,
- анафаза
- и телофаза

Рис. 1.1.33. Микроскопия фаз митоза на примере клеток бластулы (по Сopen haver, 1971): а — поздняя профаза; б — метафаза; в — ранняя анафаза; г — телофаза
Профаза характеризуется тем, что в этот период происходят интенсивные изменения структуры ядра. На участках эухроматина прекращается транскрипция. Они покрываются белками и становятся неотличимыми от зерен гетерохроматина. Затем наступает спирализация хромосом. При этом хромосома становится видимой в световом микроскопе. Вышеуказанный процесс сопровождается исчезновением ядрышка. Таким образом, в начале профазы в ядре образуется плотный клубок, который к концу фазы разрыхляется, и становятся видимыми хромосомы.
Именно в этой фазе центриоли расходятся к противоположным полюсам клетки, формируется веретено, состоящее из микротрубочек.
Метафаза (следует за профазой). В метафазе основные изменения происходят в цитоплазме клетки. Лизосомальные ферменты растворяют ядерную оболочку, и спирализованные хромосомы оказываются в цитоплазме. Комплекс Гольджи и эндоплазматический ретикулум распадаются на мелкие фрагменты.
На каждом центромере выявляется скопление специальных белков — кинетохор. Сборка микротрубочек на материнских центриолях продолжается. В результате этого процесса формируется биполярное митотическое веретено, состоящее из микротрубочек и ассоциированных с ними белков. Различают несколько видов микротрубочек. Часть микротрубочек расходится от центриоли во все стороны. Часть их образует астральную лучистость. Другая их часть направлена к экватору клетки — полярные микротрубочки. Кроме астральных и полярных микротрубочек от полюсов отходят кинетохорные микротрубочки, т. е. те, которые в области экватора прикрепляются к кинетохорам хромосом. В клетках человека каждый кинетохор связан с 20—40 микротрубочками.
Этап формирования веретена обозначают как прометафазу. В ходе собственно метафазы хромосомы перемещаются и располагаются в одной плоскости перпендикулярно к оси между полюсами. Образуется фигура, называемая материнской звездой. В результате упорядочения положения хромосом система микротрубочек также упорядочивается. Они теперь образуют веретено деления (митотическое веретено).
Важно отметить, что именно в метафазе определяют кариотип. У человека в норме кариотип характеризуется наличием 23 пар хромосом, приведенных на рис. 1.1.34.
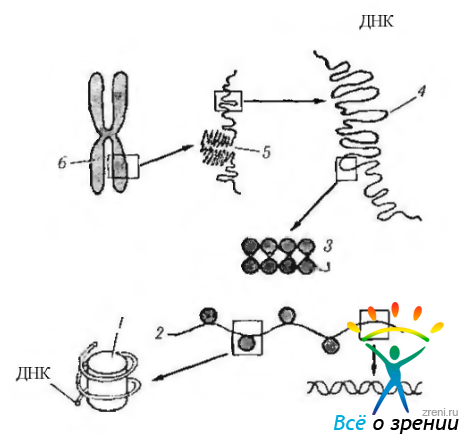
Рис. 1.1.34. Кариотип человека и уровни упаковки хроматина в ядре: а — классификация пар хромосом человека; б — молекулярная организация хромосом (по В. Л. Быкову. 1999) двойная спираль ДНК образует нить диаметром 2 нм, которая намотана на блоки дисковндшш фирмы — нуклеосомы (1), входящие в состав нуклеосомной нити (2) диаметром 11 нм Скрученная нуклеосомнля нить образует хроматиновую фибриллу (3) диаметром 30 нм, которая формирует петельные домены (4) диаметром 300 нм. Более уютно упакованные петельные до.мены образуют конденсированные участки хромосомы (5) диаметром 700 нм, являющими частью метафазной хромосомы (б) размером около 1400 нм
В метафазе изучают кариотип с диагностической целью при различных врожденных и наследуемых заболеваниях.
Анафаза. Анафаза довольно сложный в химическом отношении процесс. В начале анафазы наступает внезапное разделение центромеры d-хромосомы, в результате чего сестринские хроматиды становятся самостоятельными s-xpoмосомами.
Микротрубочки начинают укорачиваться, в результате чего хроматиды подтягиваются к центриолям. Сами центриоли удаляются друг от друга в сторону полюсов клетки, в результате чего образуются две дочерние звезды. В конце анафазы плазматическая мембрана как бы инвагинируется перпендикулярно к продольной оси митотического веретена, образуя борозду. В этой области под плазмолеммой появляется сократительное кольцо, состоящее из актин- и миозинсодержащих нитей Завершает деление телофаза.
Телофаза. Как указано выше, к концу анафазы и началу телофазы в середине клетки образуется цитоплазматическая перетяжка, которая постепенно углубляется и, в конце концов, полностью разделяет клетку на две равные части, содержащие идентичный набор хромосом. После этого вновь появляется ядро, а хромосомы «распадаются» с образованием глыбок хроматина. Примерно в середине телофазы начинается образование нитчатой, а затем гранулярной частей нуклеонеммы. К концу телофазы полностью сформировано ядрышко. Из мембранных пузырьков происходит формирование аппарата Гольджи, эндоплазматической сети. На этом митотический цикл завершается, и клетка входит в интерфазу.
Вышеприведенные данные характеризуют основной тип клеточного деления — митоз. Но существуют и другие типы деления. Это эндомитоз. Морфологически при эндомитозе удвоение числа хромосом происходит внутри ядерной оболочки без ее разрушения и формирования веретена деления. При повторных эндомитозах число хромосом в ядре может значительно увеличиваться и развивается так называемая полиплоидия. Сопровождается этот процесс значительным увеличением объема ядра. Полиплоидия сопровождается значительным увеличением функциональной активности клеток. Полиплоидия характерна как в норме, так и при различных патологических состояниях эндотелия роговой оболочки.
Полиплоидия развивается и при митотическом делении, при котором не происходит цитотомии. При последующем делении такой двуядерной клетки хромосомные наборы ядер объединяются в метафазе, приводя к образованию двух дочерних полиплоидных клеток. Наличие полиплоидных клеток (тетра-, окта- и т.д.) является нормальным состоянием ряда тканей организма человека.
Большое биологическое значение имеет еще один тип деления — мейоз, в результате которого формируются половые клетки. Основной смысл мейоза сводится к делению, при котором достигается уменьшение количества хромосом в клетке в два раза. Обсуждение этого типа деления выходит за рамки данной книги. Более подробные сведения можно получить в большом количестве руководств по цитологии.
В литературе описан еще один тип деления — амитоз. До сих пор обсуждаются вопросы возможности существования подобного типа деления. Считают, что при таком делении исчезает биологический смысл деления, т. е. возможность равного распределения генетического материала в двух вновь образованных клетках. Тем не менее морфологи, особенно патологи, довольно часто наблюдают прямое (амитотическое) деление.
↑ Межклеточные соединения
Межклеточное пространство. Между цитоплазматическими мембранами соседних клеток обнаруживается равномерное светлое пространство шириной 15 нм. Это пространство нередко расширяется или сужается как в норме, так и при патологических состояниях. Несмотря на наличие межклеточного пространства, клетки довольно сильно сцеплены между собой при помощи специализированных органоидов различного типа (рис. 1.1.35).

Рис. 1.1.35. Cxемаическое изображение межклеточных контактов различного типа (по tiogan et al., 1972)
На особенностях строения этих органоидов мы и остановимся ниже.
Десмосома (macula adhearens). Рядом расположенные клетки могут соединяться между собой при помощи локальных уплотнений — десмисим (рис. 1.1.36).
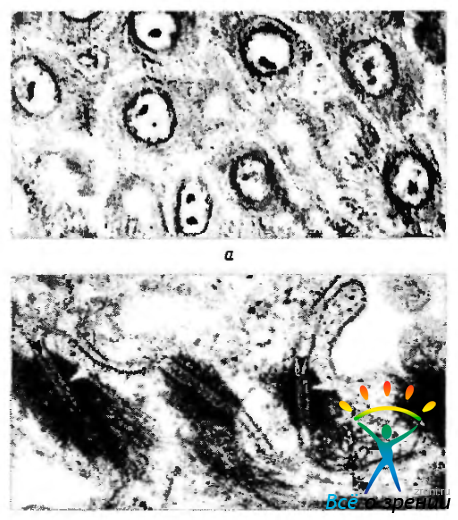
Рис. 1.1.36. Десмосомы: а — светооптическая микроскопия плоскостного препарата однослойного плоского эпителия (между клетками видны десмосомы в виде мостиков); б—ультраструктурные особенности десмосом
Этот тип органоидов относится к адгезивным (контакты типа пятна слипания). При формировании подобного типа контакта цитоплазматические мембраны соседних клеток не сливаются, а как бы «прилипают» благодаря наличию межклеточного вещества. Особенно выражен подобный тип межклеточных контактов в эпителиальных тканях (эпидермис, эпителий роговицы, нейроэпителиальные структуры).
Десмосомы настолько сильно связывают клетки между собой, что их можно разделить только механически. Даже при образовании эпителиальных пузырей на поверхности роговой оболочки (буллезная кератопатия), т.е. отслоения эпителиального пласта от подлежащей стромы, эпителиальный пласт долго сохраняет свою структуру именно благодаря десмосомам.
При ультраструктурном исследовании десмосомы представляют собой противостоящие дисковидные уплотнения мембран клеток (пластинка прикрепления). Диаметр этих пластинок около 0,5 мкм, а толщина 15 нм. Между ними определяется светлое межклеточное пространство (ширина 30 нм), выполненное «цементной» субстанцией умеренной электронноплотности в виде полоски. В межклеточном материале десмосомы определяются трансмембранные Са2+ связывающие адгезивные белки (десмоколлины, десмоглеины), которые, взаимодействуя с белками пластинок прикрепления, связывают их в единую систему. В центре межклеточной щели видно линейное уплотнение (центральная или промежуточная линия). Радиально от участков уплотнения распространяются внутрицитоплазматические промежуточные филаменты, состоящие из особых белков, — десмоплакины, плакоглобин и десмокальмин.
При контакте эпителиальной клетки с межклеточным материалом (базальная мембрана на границе со стромой) обнаруживаются не десмосомы, а полудесмосомы (рис. 1.1.37).

Рис. 1.1.37. Ультраструктурная организация полудесмосомы и базальной мембраны: 1 — полудесмосома; 2 — промежуточные филаменты; 3 плазмолемма; 4 — якорные фибриллы; 5 — базальная мембрана
По строению полудесмосома представляет собой как бы половинку десмосомы. Полудесмосома образована лишь одной пластинкой с входящими в нее тонофиламентами, которые прикрепляют клетку к базальной мембране. Подобные образования в большом количестве можно обнаружить в переднем и заднем эпителии роговой оболочки.
Промежуточное соединение, или опоясывающая десмосома (zonula adherens — поясок сцепления).
Подобного типа соединения обнаруживаются чаще всего на боковой поверхности эпителиальных клеток между областью расположения плотного соединения и десмосом. Это соединение охватывает клетку по периметру в виде пояска. В области промежуточного соединения обращенные к цитоплазме листки плазмолеммы утолщены и образуют пластинки прикрепления, которые содержат актин-связывающие белки альфа-актин, винкулин и плакоглобин. К этим пластинкам прикрепляются актиновые микофиламенты, вплетающиеся также в терминальную сеть. Межклеточная щель расширена до 15—20 нм и заполнена умеренно электронноплотным веществом, состоящим из адгезивного трансмембранного гликопротеина (Е-кадгерин) (рис. 1.1.38).

Рис. 1.1.38. Промежуточное соединение: 1 - цитоплазматические мембраны соседних клеток; 2 — микрофиламенты
Плотное соединение (zonula occludens — поясок замыкания).
Этот тип контактов относится к так называемым плотным контактам. В контактах подобного рода цитоплазматические мембраны соседних клеток как бы сливаются. При этом образуется исключительно плотная стыковка клеток (рис. 1.1.39).
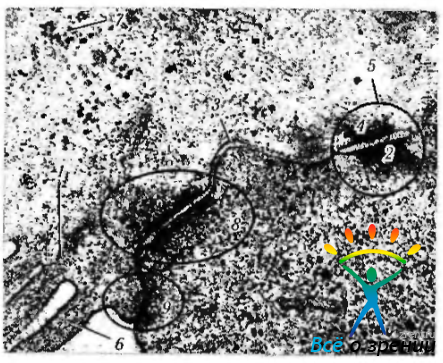
Рис. 1 1.39. Юнкциотальный комплекс эпителиальных клеток: 1 - актиновые филаменты микроворсинок; 2 — цитоплазматическая пластинка десмосомы; 3 — межклеточное пространство; 4 — промежуточные филаменты; 5 — десмосома (macula adherens): 6 — микроворсинка; 7 — полирибосомы; 8 — поясок сцепления (zonula adherens); 9 — плотное соединение (zonula occludens)
Такие контакты наиболее часто встречаются в тканях, в которых необходимо полностью предотвратить проникновение метаболитов между клетками (эпителий кишечника, эндотелий роговой оболочки). Как правило, соединения этого типа располагаются на апикальной поверхности клетки, опоясывая ее. Поясок замыкания представляет собой область частичного слияния наружных листков плазмолемм двух соседних клеток. Плотное соединение имеет вид пояска шириной 0,1 —0,5 мкм. окружающего клетку по периметру и состоящего из анастомозирующих тяжей внутримембранных частиц. Эти частицы образованы белком окклюдином. Каждая из них представляет собой область точечного слияния плазмолемм двух соседних клеток. Проницаемость плотных соединений тем ниже, чем выше число тяжей этих частиц. Эти образования могут динамично перестраиваться и временно размыкаться.
В литературе нередко используется термин «юнкциональный комплекс». При этом авторы используют этот термин, описывая сложные межклеточные соединения, включающие одновременно типичные десмосомы, зону замыкания и ленту замыкания (fascia occludens) (рис. 1.1.39). Последний тип контакта отличается от зоны замыкания лишь тем, что контакт не полностью опоясывает клетку, а местами прерывается.
Щелевидное соединение (nexus). Щелевидное соединение характеризуется тем, что между цитоплазматическими мембранами соседних клеток видна щель шириной в 2 нм. При этом обе плазмолеммы соседних клеток соединены между собой коннексонами — полыми гексагональными белковыми структурами размерами около 9 нм. каждая из которых образована шестью белковыми субъединицами (рис. 1.1.40).
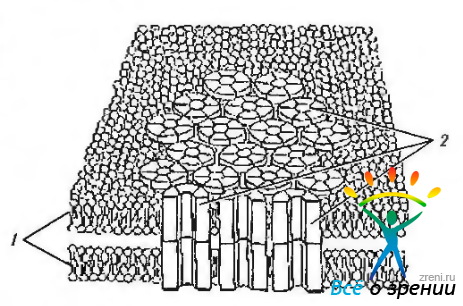
Рис. 1.1.40. Схематическое изображение щелевидного соединения: 1 -цитоплазматические мембраны соседних клеток; 2—коннексоны
Число коннексонов в щелевом соединении обычно исчисляется сотнями. Через эти образования осуществляется перенос метаболитов из одной клетки в другую. При этом молекулярный вес этих веществ не превышает 1500 Да (неорганические ионы, сахара, витамины, аминокислоты, нуклеотиды, АТФ и др.). Эти соединения обеспечивают между клетками ионное и метаболическое сопряжение. Близкое строение имеют синапсы.
Щелевидные контакты играют важную роль в осуществлении функций клеток, особенно обладающих выраженной электрической активностью. По этой причине неудивительно, что большое количество подобных контактов обнаруживается между нейронами сетчатой оболочки (см. Сетчатка). Широко распространены они также в сердечной мышце и ткани центральной нервной системы.
Интердигитации (пальцевые соединения). Интердигитации представляют собой межклеточные соединения, образованные выпячиваниями цитоплазмы одних клеток, вдающимися в цитоплазму других (рис. 1.1.41).

Рис. 1.1.41. Интердигитации соседних эпителиальных клеток: а — трансмиссионная электронная микроскопия (1 — интердигитации; 2 — митохондрии: 3 — десмосомы); б—метод замораживания-скалывуния (сканирующая электронная микроскопия)
Между плазмолеммами соседних клеток всегда остается межклеточная щель 15—20 нм. За счет интердигитации увеличивается прочность межклеточного сцепления и увеличивается поверхность контакта между клетками.
----
Статья из книги: Строение зрительной системы человека | Вит В.В.

Комментариев 0