Ткани │ Часть 2


Содержание:
Описание
↑ Мышечная ткань
Мышечные ткани представляют собой группу тканей различного происхождения и строения, объединенных на основании общего признака — выраженной сократительной способности. Сократимость свойственна в той или иной степени клеткам всех тканей организма вследствие наличия в их цитоплазме сократительных микрофиламентов. Однако мышечные ткани специализированы на этой функции, что обеспечивается особыми свойствами их сократительного аппарата.
Мышечная ткань довольно широко представлена в глазном яблоке и глазнице. Различают два основных типа мышечной ткани — гладкая и поперечнополосатая. Для глазного яблока свойственно наличие и третьего типа мышц, имеющих не мезенхимальное, как предыдущие мышцы, а нейроэктодермальное происхождение. Это сфинктер и дилятатор радужной оболочки. О них речь идет в разделе «Радужная оболочка».
Гладкая мышечная ткань. Гладкая мышечная ткань (рис. 1.4.14)

Рис. 1.4.14. Гладкомышечная ткань: а — гладкомышечные клетки складываются в пучки, между которыми видны прослойки соединительной ткани; б—цитологические особенности гладкомышечных клеток. Ядра палочковидной формы. В цитоплазме видны миофиламенты
является структурным компонентом стенок сосудов, большинства полых органов. В глазнице она формирует мышцу Мюллера. К гладкой мышце относится и ресничная мышца.
Основным структурным элементом гладкой мышцы является мышечная клетка (гладкий миоцит), имеющая, как правило, веретеновидную или звездчатую форму. Длина этих клеток довольно разнообразна (от 20 до 1000 мкм). Гладкие миоциты окружены сарколеммой, которая снаружи покрыта базальной мембраной. В саркоплазме обнаруживаются органеллы и включения. Поскольку сокращение требует затраты большого количества энергии, цитоплазма мышечных клеток насыщена профилями саркоплазматического ретикулума (эндоплазматический ретикулум). В клетке, как правило, одно ядро, которое располагается вдоль клетки. Периферическая часть саркоплазмы занята миофиламентами (рис. 1.4.15).
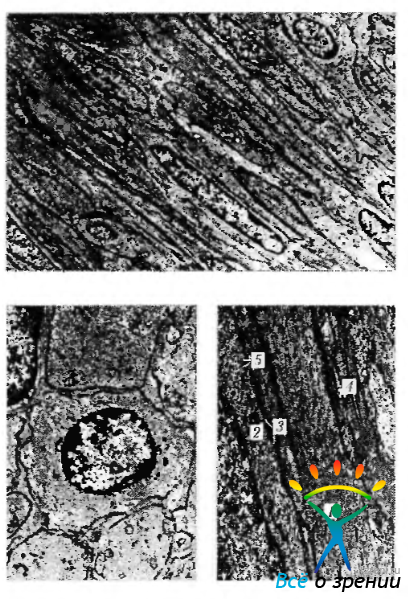
Рис. 1.4.15. Ультраструктурная организация гладкомышечных клеток: а— продольный срез; б -поперечный срез; в — большее увеличение (1 — актиновые фибриллы; 2 — темные зоны; 3 — плотные тельца; 4 — коллагеновые волокна; 5 — пузырьки)
Отдельные мышечные клетки складываются в плотный пучок. В зависимости от типа органа или ткани отдельные клеточные пучки ориентируются в стенке различным образом, но всегда так, чтобы их сокращение поддерживало тонус стенки (сосуда, стенки желудка и т. д.).
Функцию сокращения мышечной клетки и комплекса мышечных клеток обеспечивают тонкие (актиновые) и толстые (миозиновые) миофиламенты. Эти филаменты фибрилл не образуют. Тонкие филаменты преобладают над толстыми по количеству и занимаемому ими объему клетки. Располагаются они пучками, по 10—20 филаментов, лежащих параллельно оси клетки. Концы актиновых филаментов закреплены в особых образованиях, находящихся в саркоплазме — плотных тельцах. Последние также служат местом прикрепления промежуточных филаментов.
Миозиновые (толстые) филаменты отличаются от таковых поперечнополосатой мышцы различной длиной. Сокращение гладких миоцитов обеспечивается взаимодействием актиновых и миозиновых филаментов и развивается в соответствии с моделью скользящих нитей. Возникающая сила передается через внутри -цитоплазматические филаменты плотным тельцам, прикрепленным к сарколемме. Благодаря этому продольная ось волокна укорачивается (рис. 1.4.16, 1.4.17).
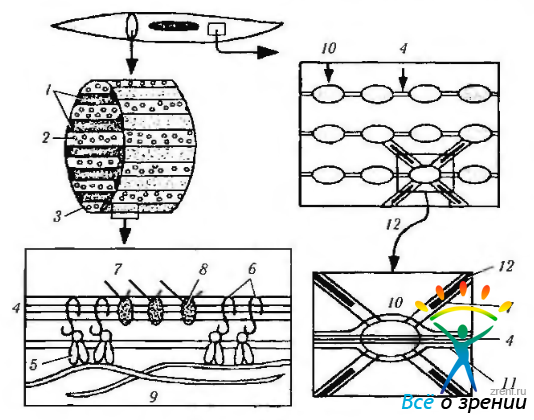
Рис. 1.4.16. Взаимосвязь элементов цитоскелета и сократительного аппарата гладкомышечной клетки (по В. Л. Быкову, 1999): 1 — плотные пластинки; 2— кавеолы; 3— сарколемма; 4— немышечный актин; 5 — интегрины; 6 — комплекс адгезивных белков; 7 — мышечный актин; 8 — связывающие белки; 9 — межклеточное вещество; 10— плотные тельца; 11 — промежуточные филаменты; 12 — миозиновые миофиламенты

Рис. 1.4.17. Механизм сокращения гладкомышечной клетки
Отдельные мышечные клетки очень компактно располагаются и разделены промежутками 40—80 нм. Межклеточные пространства выполнены компонентами базальной мембраны, коллагеновыми, эластическими волокнами, которые совместно с отдельными клетками (фибробластами, тучными клетками) образуют эндомизий. Последний содержит сосуды и нервные волокна и способствует объединению миоцитов в пласты и слои (рис. 1.4.18).
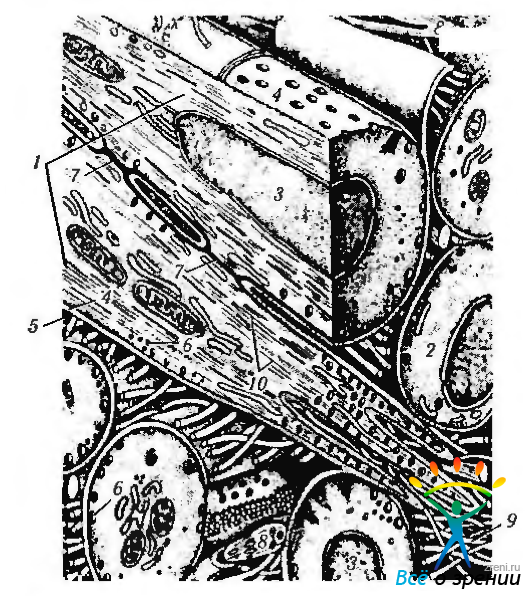
Рис. 1.4.18. Схема строения гладкой (по Р. Кристину): 1 — веретеновидные гладкие миоциты: 2 — цитоплазма миоцита; 3 — ядра миоцитов; 4 — плазмолемма; 5 — базальная мембрана; 6 — поверхностные пиноцитозные пузырьки; 7 — межклеточные соединения; 8— нервное окончание; 9— коллагеновые фибриллы; 10—микрофиламенты
Формированию пласта миоцитами способствует образование ими различных связей (по типу миоцит—миоцит, миоцит—клетка другого типа, миоцит—межклеточное вещество). В местах межклеточных соединений базальная мембрана отсутствует. Межклеточные соединения в пластах обеспечивают механическую и химическую (ионную) связь между ними. К соединениям между гладкими миоцитами относят интердигитации, плотные соединения, щелевые соединения (нексусы).
Благодаря вышеописанным связям сокращение отдельных клеток передается всему клеточному пласту, который обладает свойством обратимой деформации.
Сокращение гладкой мышечной ткани происходит под воздействием нервных импульсов, аморальных влияний, а также вследствие раздражения миоцитов в отсутствие нервных и гуморальных воздействий (миогенная активность).
Иннервация гладкомышечной ткани осуществляется вегетативной нервной системой (симпатическая и парасимпатическая). Нервные окончания обнаруживаются лишь в отдельных клетках и имеют вид варикозно расширенных участков тонких веточек аксонов. На соседние миоциты возбуждение передается при помощи щелевых соединений.
Возможность гормональной регуляции активности миоцитов связана с наличием в клетках соответствующих рецепторов. Благодаря этому на клетки влияют такие вещества, как
- гистамин,
- серотонин,
- брадикинин,
- эндотелии,
- окись азота,
- лейкотриены,
- простагландины,
- нейротензин,
- вещество Р,
- бомбезин,
- холецитокинин,
- вазоактивный интерстициальный пептид,
- опиоиды и др.
Растяжение мышцы является физиологическим раздражителем гладкой мышцы. При этом наступает деполяризация сарколеммы и усиливается приток ионов кальция в саркоплазму. Гладкая мышечная ткань характеризуется спонтанной ритмической активностью вследствие циклически меняющейся активности кальциевых насосов.
Гладкомышечная ткань способна к функциональной гипертрофии. Обладает она в определенной степени и способностью к регенерации (физиологической и репаративной).
Необходимо упомянуть еще о некоторых типах клеток, сходных с гладкомышечными. Это клетки, окружающие секреторные альвеолы экзокринных желез (молочные, потовые, слезные и др.). Их цитоплазма содержит миофиламенты. Поскольку эти клетки не мезенхимного, а эктодермального происхождения, их назвали миоэпигпелиальными клетками (рис. 1.4.19).
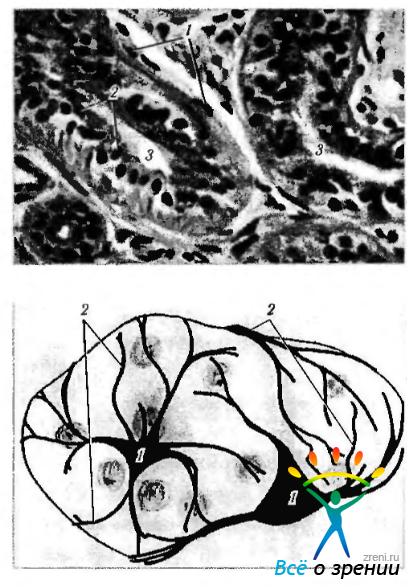
Рис. 1.4.19. Миоэпителиоциты: а — миоэпителиоциты альвеолярно-трубчатой железы (1 — миоэпителиоциты; 2 — эпителий железы; 3 — просвет железы); б — схема расположения тел и отростков миоэпителиоцитов (1— тела клеток; 2— отростки клеток, охватывающие снаружи железу)
С железистыми клетками миоэпителиальные клетки связаны десмосомами. Снаружи они покрыты базальной мембраной. Форма миоэпителиальных клеток в концевых отделах — отростчатая или звездчатая. Эти клетки получили также название корзинчатых, поскольку образуют как бы корзинку, охватывающую железистые клетки.
Помимо миофиламентов эти клетки содержат свойственные эпителиальным клеткам промежуточные филаменты типа цитокератанов. Иммуноцитомическими методами выявляется и свойственный мышечным тканям промежуточный филамент — десмин.
Другой тип клеток обнаруживается в стенках семенных канальцев яичка — миоидные клетки.
Существуют так называемые эндокринные гладкие миоциты, которые обнаруживаются в виде структурного компонента юкстагломерулярного аппарата почек, входя в состав стенки артериол почечного тельца. Эти клетки продуцируют ренин.
Миофибробласты, клетки мезенхимного происхождения. обладающие сократительной функцией, нами описаны выше.
Последний тип сократительных клеток имеет нейроэпителиальное происхождение. Это мионейральные клетки. Поскольку эти клетки обнаруживаются в глазном яблоке, о них подробно будет изложено в соответствующих разделах (см. Радужная оболочка).
Поперечнополосатая мышечная ткань. Поперечнополосатая мышечная ткань (скелетная мышечная ткань) широко распространена в глазнице, в частности из нее сосстоят наружные мышцы глаза, мышцы века и др. Развивается она из мезенхимы. Правда, в области головы и шеи ее происхождение связывают с эктомезенхимой (см. главу 5).
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Основным структурным компонентом поперечнополосатой мышцы является поперечнополосатое мышечное волокно (рис. 1.4.20).

Рис. 1.4.20. Микроскопическое строение поперчнополосатой мышечной ткани: а — светооптическое строение поперечнополосатой мышечной ткани (четко видна поперечная исчерченность мышечных волокон разделенных прослойками соединительнои гкани); б - большое увеличение мышечного волокна.
Длина волокон в зависимости от типа мышцы довольно разнообразна и колеблется от нескольких миллиметров до нескольких десятков сантиметров. Диаметр также различен (12—70 мкм).
Мышечное волокно снаружи покрыто цитоплазматической оболочкой (сарколеммой) и состоит из цитоплазмы (саркоплазмы), в которой видно множество ядер овальной формы, располагающихся по периферии волокна под сарколеммой и ориентированных параллельно ей (рис. 1.4.21).

Рис. 1.4.21. Ультраструктурная организация миофибриллы: а — продольный разрез мышечного волокна; б — продольный срез саркомера (по обеим сторонам Z-линий видны половинки слабоокрашенных I-полос, содержащих только тонкие филаменты. Эти филаменты тянутся от Z-линий и проходят некоторое расстояние между толстыми филаментами, лежащими в более темной A-полосе. Участки A-полосы содержат как тонкие, так и толстые филаменты и поэтому кажутся более темными, чем та часть, где проходят только толстые филаменты — Н-зона. Через середину A-полосы проходит более темная М-линия); в — поперечный срез миофибриллы (видны тонкие и толстые филаменты. Тонкие филаменты образуют шестиугольную фигуру, в центре которой находится толстый филамент)
Саркоплазма содержит многочисленные органоиды—саркоплазматический ретикулум, митохондрии и свободные рибосомы, расположенные вблизи сарколеммы, а также зерна гликогена. Для саркоплазмы характерно наличие специфического растворимого пигментированного белка — миоглобина. близкого по строению к гемоглобину эритроцитов.
Именно они обусловливают поперечную и продольную исчерченность, видимую как в световом, так и электронном микроскопах. Миофибриллы складываются в пучок, расположенный вдоль оси волокна.
Наличие поперечной исчерченности является результатом особой организации миофибрилл и связано с чередованием участков различного химического состава и оптических свойств. Одинаковые участки миофибрилл располагаются на одном уровне, что и приводит к поперечной исчерченности на протяжении всего волокна.
Поперечная исчерченность скелетных мышечных волокон обусловлена чередованием темных A-дисков (анизотропных, обладающих двойным лучепреломлением в поляризованном свете) и светлых I-дисков (изотропных не обладающих двойным луче преломлением). Каждый диск I рассекается надвое тонкой темной Z-линией, называемой также телофрагмой. В середине A-диска определяется светлая зона — полоска Н, через центр которой проходит М-линия— мезофрагма (рис. 1.4.21 —1.4.23).
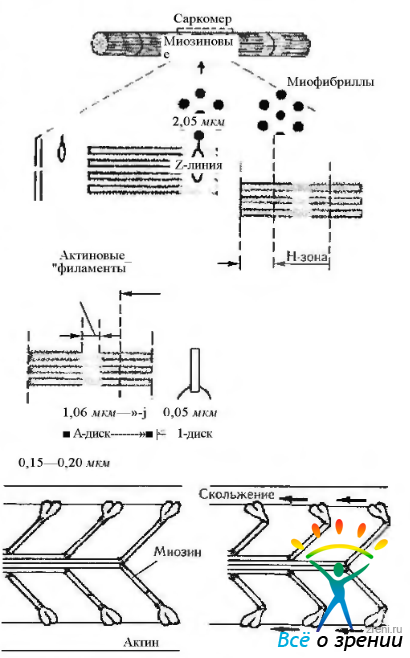
Рис. 1.4.22. Структура саркомсра и механизм сокращения филаментов (объяснение в тексте)
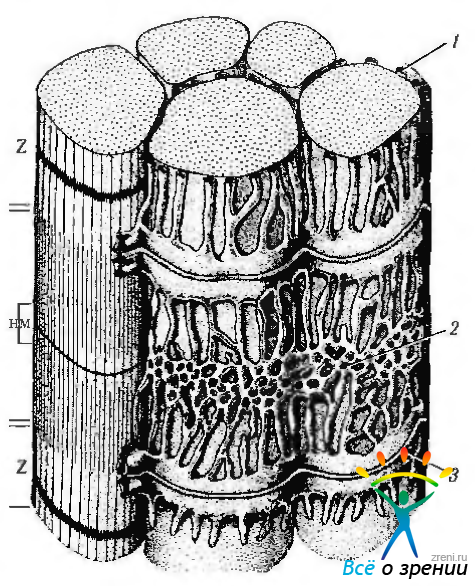
Рис. 1.4.23. Саркотубулярная структура поперечнополосатого мышечного волокна: 1 — сарколемма; 2 — саркоплазматические трубочки; 3 — Т-трубочки
Саркомер (миомер) представляет собой участок миофибриллы, расположенный между двумя телофрагмами (Z-линиями) и включающий A-диск и две половины I-дисков — по одной половине с каждой стороны. В расслабленной мышце длина саркомера составляет около 2—3 мкм, а ширина его участков выражается соотношением Н:А:1= 1:3:2. При сокращении мышцы саркомер укорачивается до 1,5 мкм.
Структура саркомера представлена упорядоченной системой толстых и тонких белковых нитей (миофиламентов). Толстые нити (диаметром около 10—12 им и длиной 1,5—1,6 мкм) связаны с мезофрагмой и сосредоточены в А-диске, а тонкие (диаметром 7—8 нм и длиной 1 мкм) — прикреплены к телофрагмам, образуют I-диски и частично проникают в A-диски между толстыми нитями (более светлый участок А-диска, свободный от тонких волокон, называется полоской Н). В capкомере насчитывается несколько сотен толстых нитей. По сечению саркомера толстые и тонкие нити располагаются высокоорганизованно в углах гексагональной решетки. Каждая толстая нить окружена шестью тонкими, каждая из тонких нитей частично входит в окружение трех соседних толстых.
Толстые нити (миофиламенты) образованы упакованными молекулами фибриллярного белка миозина. Молекула миозина имеет вид нити длиной 150 нм и толщиной 2 нм. На одном из концов эта молекула содержит две округлые головки длиной около 20 нм и шириной около 4 нм. Протеологическими ферментами миозин расщепляется на легкий меромиозин («стержень» молекулы миозина) и тяжелый меромиозин (участки головок и шейки, связывающей их со стержневой частью). Молекула миозина может сгибаться, как на шарнирах, в месте соединения тяжелого меромиозина с легким в области прикрепления головки. Стержневые части молекул миозина собраны в пучки. Такие пучки, соединенные зеркально концами друг с другом в области M-линии, формируют толстые нити с центральной гладкой частью длиной около 0,2 мкм и двумя периферическими участками, в которых от центрального стержня отходят миозиновые головки (около 500). Миозин головок обладает АТФ-азной активностью, однако в отсутствие его взаимодействия с актином скорость гидролиза АТФ ничтожно мала.
Тонкие нити (миофиламенты) содержат сократимый белок актин и два регуляторных белка — тропонин и тропомиозин. Последние формируют единый тропонин-тропомиозиновый комплекс. Актин в мономерной форме представлен полярными глобулярными белками (G-актин), которые имеют активные центры, способные связываться с молекулами миозина. G-актин агрегирует с образованием полимерного фибриллярного актина (F-актина), молекула которого имеет вид двух скрученных нитей толщиной 7 нм и вариабельной длины.
Тропомиозин представлен нитевидными молекулами, которые соединяются своими концами, образуй длинный тонкий тяж, лежащий в борозде, образуемой перевитыми нитями F-актина. Так как таких борозд на молекуле актина две, то и тропомиозиновых нитей тоже две. Всего в состав тонкой нити входит примерно 50 молекул тропомиозина.
Тропонин представляет собой глобулярный белок. Каждая его молекула располагается на тропомиозиновой молекуле вблизи ее конца. Тропонин состоит из трех субъединиц: ТnС — связывающий кальций, ТnТ — прикрепляющийся к тропомиозину и Tnl — ингибирующий связывание миозина с актином.
Механизм мышечного сокращения описывается теорией скользящих нитей, согласно которой укорочение каждого саркомера (а следовательно, миофибрилл и всего мышечного волокна) при сокращении происходит благодаря тому, что тонкие нити вдвигаются в промежутки между толстыми нитями без изменения их длины. Скольжение нити в саркомере и усилие, развиваемое мышцей, обеспечиваются благодаря циклической активности миозиновых мостиков, которые при сокращении повторно прикрепляются к актину, обеспечивают усилие тяги, а затем открепляются от него. В этом механизме АТФ играет двойную роль, обеспечивая энергию, необходимую как для сокращения, так и открепления мостиков.
Строгая пространственная упорядоченность взаимодействия множества толстых и тонких нитей в саркомере определяется наличием сложно организованного поддерживающего аппарата. Его элементы на всех этапах мышечного сокращения и расслабления, динамично перестраиваясь, фиксируют и удерживают миофиламенты в правильном положении, которое оптимальным образом обеспечивает их взаимный контакт, взаимодействие и взаимное скольжение.
Опорный аппарат мышечного волокна включает особые элементы цитоскелета и связанные с ними сарколемму и базальную мембрану, соединяющие мышечное волокно с сухожилием, на которое передается усилие, развиваемое волокном при сокращении. К опорным элементам мышечного волокна относится телофрагма (Z-линия) (область прикрепления тонких миофиламентов двух соседних саркомеров), мезофрагма (М-линия, область закрепления толстых филаментов в саркомере), тинин (коннектин. белок с эластическими свойствами, нити которого присоединены к толстым филаментам по всей длине и, продолжаясь на I-диски, прикрепляют концы толстых филаментов к Z-линиям), небулин (отвечает за поддержание длины тонких филаментов), промежуточные филаменты (десминсодержащие филаменты, связывающие соседние телофрагмы одной миофибриллы, а также прикрепляющие телофрагмы к сарколемме и элементам Т-трубочек), дистрофии (белок, связывающий сарколемму с компонентами базальной мембраны), костамеры (кольца из белка винкулина, связывающие сарколемму с I-дисками миофибрилл).
Иннервация мышцы. Каждое мышечное волокно иннервируется концевой веточкой двигательного нейрона. Один мотонейрон, его аксон вместе с иннервируемым мышечным волокном образует нервно-мышечное соединение (рис. 1.4.25, 1.4.26).

Рис. 1.4.25. Моторные бляшки на поперчнополосатых мышечных волокнах
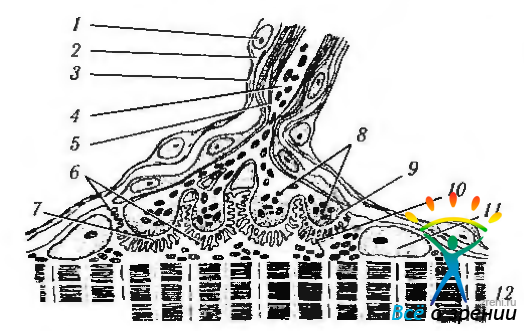
Рис. 1.4.26. Схема строения нервно-мышечного окончания: 1 — ядро нейролеммоцита; 2— цитоплазма нейролеммоцита; 3— плазмолемма нейролеммоцита; 4 — осевой цилиндр нервного волокна; 5 — аксолемма; 6 — постсинаптическая мембрана; 7 — синаптическая щель; 8— пресинаптичские пузырьки; 9 — пресинаптическая мембрана (аксолемма); 10— сарколемма; 11 — ядро мышечного волокна; 12— миофибриллы
В месте контакта аксон и его оболочка образуют на поверхности мышечного волокна двигательную концевую пластинку. В этой области между аксоном и сарколеммой образуется синаптическая щель. Синаптическая щель содержит ацетилхолинэстеразу, необходимую для инактивации нейромедиатора ацетилхолина, высвобождаемого в концевой двигательной пластинке.
В окончаниях аксонов обнаруживается множество синаптических пузырьков, содержащих ацетилхолин. Волна деполяризации приводит к высвобождению ацетилхолина путем эндоцитоза в синаптическую щель. В результате происходит снижение потенциала покоя сарколеммы и возникает волна деполяризации, распространяющаяся от концевой пластинки по всей саркоплазме. Волна деполяризации достигает саркоплазматического ретикулума, который, в свою очередь, управляет мышечным сокращением.
Скелетные мышцы снабжены не только эфферентными (двигательными), но и афферентными (чувствительными) нервными волокнами, с помощью которых они передают мозгу информацию о степени своего сокращения.
Мышца как анатомическое образование. Снаружи мышца покрыта плотной волокнистой соединительной тканью — эпимизием (рис. 1.4.27).
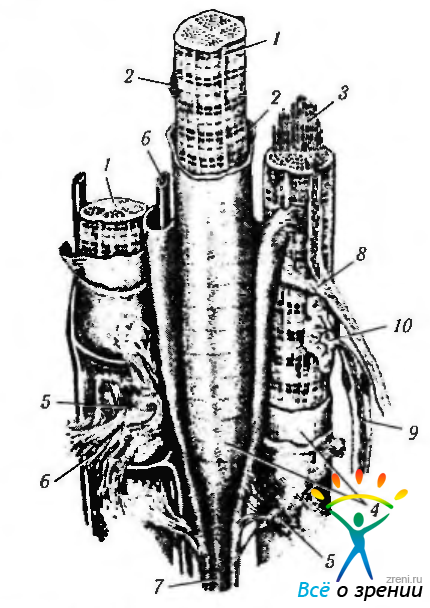
Рис. 1.4.27. Схематическое изображение мышцы как органа (по В. Г. Елисееву и соавт., 1972): 1 — мышечные волокна; 2 — ядра; 3— миофибриллы; 4— сарколемма; 5 — эндомизий; 6- кровеносные капилляры; 7 — сухожильная нить; 8— вегетативное нервное волокно; 9 — двигательное нервное волокно; 10 — аксоно-мышечный синапс
От эпимизия в глубь мышцы отходят соединительнотканные перегородки, содержащие большое количество капиллярных кровеносных сосудов — перимизий. В перемизии располагаются также лимфатические сосуды и нервные волокна. От перемизия отходят тонкие прослойки соединительной ткани, содержащей фибробласты, немного межклеточного вещества и редкие коллагеновые волокна. Эта ткань образует сеть вокруг отдельных мышечных волокон и называется эндомизием. На обоих концах мышцы соединительнотканные элементы продолжаются и смешиваются с плотной соединительной тканью, прикрепляющей мышцу к той структуре, к которой должно быть приложено тянущее усилие. Это может быть как костная, так и соединительная ткань (апоневроз, шов, надкостница, плотная соединительная ткань кожи и др.). При присоединении к кости формируется сухожилие.
Типы мышечных волокон. В различных участках организма мышечные волокна могут довольно существенно отличаться строением и функцией. Условно выделяют три типа мышечных волокон:
- тип I (красные),
- тип ПВ (белые)
- и тип ПА (промежуточные).
Мышцы типа I характеризуются малым диаметром, относительно тонкими миофибриллами, высокой активностью окислительных ферментов, низкой активностью гидролитических ферментов и миозиновой АТФ-азы, преобладанием аэробных процессов, высоким содержанием миоглобина, крупных митохондрий, интенсивным кровоснабжением. Основным отличием этих волокон в функциональном отношении является их способность к длительным тоническим сокращениям с небольшой силой сокращения.
Мышечные волокна типа ПВ характеризуются большим диаметром, крупными и сильными миофибриллами, высокой активностью гидролитических ферментов, низкой активностью окислительных ферментов, преобладанием анаэробных процессов, низким содержанием митохондрий, липидов и миоглобина. Подобные волокна выполняют быстрые сокращения большой мощности. Они быстро утомляются.
Мышечные волокна ПА типа напоминают волокна I типа. В функциональном отношении они занимают промежуточное положение между вышеописанными волокнами.
В мышцах определяется различное соотношение волокон разного типа. Свойственно это и наружным мышцам глаза.
Регенерация мышечной ткани. Поперечнополосатая мышечная ткань регенерирует на протяжении всей жизни (физиологическая регенерация). При этом происходит самообновление органоидов и других структурных компонентов.
Репаративная регенерация мышечных волокон направлена на восстановление их целостности после повреждения. При любых видах травмы процесс регенерации включает ряд последовательных процессов.
На первом этапе происходит инфильтрация поврежденного участка фагоцитами (нейтрофильные гранулоциты и макрофаги). Миграция происходит в область повреждения под хемотаксическим действием продуктов, выделяемых травмированными волокнами. Фагоциты поглощают тканевой детрит. Параллельно происходит восстановление целостности сосудов (реваскуляризация).
Следующий этап сводится к пролиферации миогенных клеток-предшественников (миосателлициты), которые сливаются с формированием мышечных трубочек. В последующем происходит дифференцировка трубочек с образованием зрелых мышечных волокон. Завершает процесс восстановление иннервации мышцы.
Полноценная регенерация поперечнополосатой мышечной ткани возможна лишь при незначительных дефектах. Необходимым условием регенерации является сохранение базальной мембраны, служащей барьером для проникновения клеток фибробластического ряда в поврежденное волокно. Базальная мембрана также играет роль направляющей, поддерживающей и ориентирующей структуры для мигрирующих миосателлицитов и для формирующихся мышечных трубочек. Неполноценная регенерация наступает при обширных повреждениях. Полноценной регенерации в этих случаях обычно препятствует разрастание соединительной ткани эндо- и перимизия. Поврежденная мышца замещается соединительнотканным рубцом.
В последние годы разработаны методы использования миосателлицитов для стимуляции регенерации мышечной ткани путем введения взвеси клеток в регенерирующую мышцу.
↑ Нервная ткань
Основной задачей данного раздела является изложение сведений о клеточной и тканевой организации нервной системы.
В центральной нервной системе различают две группы клеточных элементов — нейроны и нейроглии.
Нейроны (рис. 1.4.29)
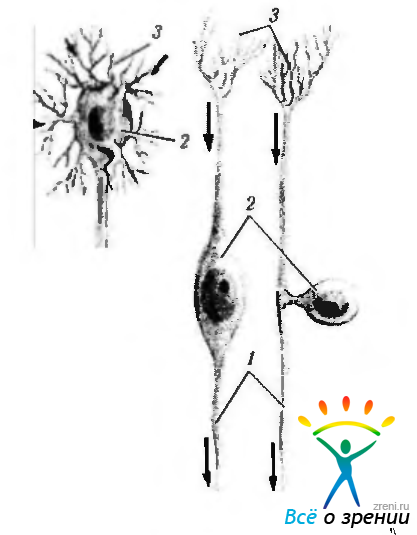
Рис. 1.4.29. Основные морфологические типы нейронов: а — мультиполярный нейрон; 6 — биполярные нейроны (1 — аксон: 2 — тело клетки; 3 — дендриты)
состоят из тела клеток и отростков. В зависимости от количества отростков различают псевдоуниполярные (биполярные нейроны, у которых два отростка вблизи тела клетки приближаются друг к другу, в результате чего складывается впечатление о су ществовании только одного отростка), биполярные (нейроны, имеющие два отростка — аксон и дендрит) и мулътиполярные нейроны (нейроны, от тела которых отходит более двух цитоплазматических отростков — аксон и многочисленные дендриты).
Независимо от количества отростков нейроны всегда содержат один отросток, по которому информация передается от нейрона к исполняющему органу. Этот отросток имеет различную длину и может достигать нескольких десятков сантиметров. Называется он аксоном (нейрит).
Остальные отростки, а их число может быть самым различным, отличаются тем, что по ним информация подходит к нейрону. Эти отростки дихотомически ветвятся, в связи с чем получили название дендриты.
Нейроны довольно существенно отличаются по строению от клеток других тканей. Это, в первую очередь, относится к телу нейрона. Тела нейронов обычно крупные и могут иметь самую разнообразную форму. Ядро, как правило, располагается в центре клетки, содержит незначительное количество зерен гетерохроматина и обладает 2—3 крупными четкими крутлыми ядрышками. Эти особенности отражают высокую активность процессов транскрипции в ядре нейрона. Около ядрышка в нейронах у женщин часто выявляется тельце Барра — крупная глыбка хроматина, содержащая конденсированную Х-хромосому.
Цитоплазма нейронов содержит большинство известных органоидов и окружена плазмолеммой, способной к проведению нервного импульса вследствие локального тока Na в цитоплазму и К+ из нее через потенциал-зависимые мембранные ионные каналы.
Для нейронов характерны и специфические черты организации цитоплазмы. К таковым можно отнести наличие вещества Ниссля (тигроидное вещество, тигроид) (рис. 1.4.29). Вещество Ниссля обнаруживается в крупных нейронах (ганглиозные клетки сетчатки) и представляет собой крупные базофильные внутрицитоплазматические глыбки. Ультраструктурно показано, что вещество Ниссля есть не что иное, как насыщение цитоплазмы цистернами гранулярного эндоплазматического ретикулума, содержащими обилие свободных и связанных рибосом и полирибосом. Это указывает на исключительно высокий белковый метаболизм в нейронах. Важно подчеркнуть, что при развитии патологических состояний, сопровождающихся повреждением отростков нейрона или тела, вещество Ниссля исчезает (хроматолиз). Нередко хроматолиз является первым признаком развивающегося заболевания.
Второй особенностью организации цитоплазмы нейрона является наличие нейротрубочек, нейрофиламентов (промежуточные филаменты) (рис. 1.4.30, 1.4.31).
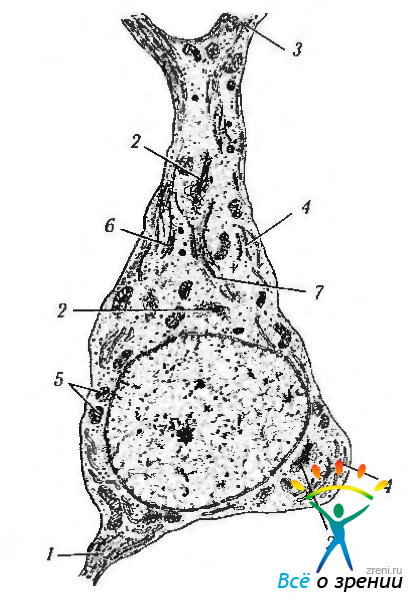
Рис. 1.4.30. Схема ультраструктурной организации нейрона (по А. Хэм, Д. Кормак, 1983): 1—аксонный холмик; 2—комплекс Гольджи; 3—дендриты; 4 — гранулярный эндоплазматический ретикулум; 5 — митохондрии; 6—микротрубочки; 7 — филаменты

Рис. 1.4.31. Нейротрубочки и нейрофиламенты нейронов: а — продольный срез слоя нервных волокон сетчатки (по Hogan et al., 19~1); 6 — поперечный срез слоя нервных волокон сетчатки (1 — нейротрубочки; 2— нейрофиламенты)
Нейрофиламенты диаметром 10 нм складываются в пучки, выполняющие цитоплазму клетки.
Нейротрубочки выполняют опорную функцию, особенно в цитоплазматических отростках, и имеют типичное строение. Их диаметр 24 нм.
К другим особенностям строения нейрона необходимо отнести и возможность наличия в их цитоплазме двух типов пигмента — меланина (substantia nigra) и липофусцина.
Особенности строения отростков нервных клеток имеет смысл рассматривать в связи с выполняемыми ими функциями.
Как указывалось выше, аксон (нейрит) может иметь довольно большую длину (от 1 мм до 1,5 м у человека). Аксон отходит от утолщенного участка тела нейрона, не содержащего хромофильной субстанции, — аксонного холмика, в котором генерируются нервные импульсы. Он почти на всем протяжении покрыт глиальной оболочкой. Аксон может по своему ходу давать ответвления (коллатерали), которые обычно отходят от него под прямым углом. В конечном участке аксон нередко распадается на тонкие веточки (телодендрии). Аксон заканчивается специализированными терминалями (нервными окончаниями) на других нейронах или клетках рабочих органов.
Отличается по строению и цитоплазма аксона. Центральная часть цитоплазмы аксона (аксоплазма) содержит большое количество аксоплазматических пузырьков, нейрофиламентов, микротрубочек, ориентированных продольно. Гранулярный эндоплазматический ретикулум. аппарат Гольджи отсутствуют. Встречаются редкие рибосомы.
Таким образом, видно, что белковый метаболизм в аксоне определяется обменом, происходящим в теле клетки. Поскольку объем аксона может быть во много десятков раз больше объема тела нейрона, метаболизм в аксоне поддерживается специальным механизмом — аксонным транспортом.
Различают два типа аксонного транспорта —
- антероградный (из тела нейрона по аксону)
- и ретроградный (направлен от терминалей в сторону тела клетки).
Медленный поток несет пузырьки, лизосомы и ферменты, участвующие в синтезе нейромедиаторов (норадреналина). Быстрый поток несет некоторые ферменты, участвующие в выполнении синаптической функции, гликопротеиды, фосфолипиды, митохондрии и нейросекреторные гранулы.
Ретроградный аксонный транспорт (100— 200 мм сут.) способствует удалению веществ из области терминалей, возвращению пузырьков, митохондрий.
Феномен транспорта используется для изучения межнейронных связей путем введения маркеров в область расположения терминалей или клеточных тел и выявления областей его последующего распространения описанными механизмами.
Дендриты, в отличие от аксона, дихотомически делятся. Крупные дендриты отличаются от аксона тем, что они содержат рибосомы и цистерны гранулярного эндоплазматического ретикулума, а также нейротрубочки, нейрофиламенты и митохондрии. Существует и дендритный транспорт со скоростью, примерно равной быстрому потоку в аксоне.
После описания структуры нейронов логично остановиться на механизмах передачи нервного импульса. Связь нервных клеток между собой осуществляется с помощью синапсов.
Синапсы. Синапсы подразделяются на электрические и химические.
Синапсы могут различным образом располагаться на нейроне. В тех случаях, когда аксон оканчивается на дендрите, синапс называют аксодендритным, а когда на теле нейрона— аксосинаптическим. Аксоны, заканчивающиеся на других аксонах, называются аксо-аксональными (рис. 1.4.32).

Рис. 1.4.32. Схематическое изображение различных типов синаптических соединений между нейронами: 1 — аксосоматический синапс; 2 — аксодендритные синапсы различных типов; 3 — аксоаксонный синапс; 4 — аксодендритные синапсы с дендритным шипиком
Электрические синапсы в центральной нервной системе млекопитающих редки. Довольно большое их количество в сетчатой оболочке. Эти синапсы имеют строение щелевых соединений, в которых мембраны синаптически связанных клеток (пре- и постсинаптическая) разделены промежутком шириной 2 нм, пронизанным коннексонами. Последние представляют собой трубочки, образованные белковыми молекулами и служащие водными каналами, через которые мелкие молекулы и ионы могут транспортироваться из одной клетки в другую. Когда потенциал действия, распространяющийся по мембране одной клетки, достигает области щелевого соединения, электрический ток пассивно протекает через щель от одной клетки к другой. Импульс способен передаваться в обоих направлениях и практически без задержки.
Химический синапс при ультраструктурном исследовании характеризуется следующим (рис. 1.4.33).
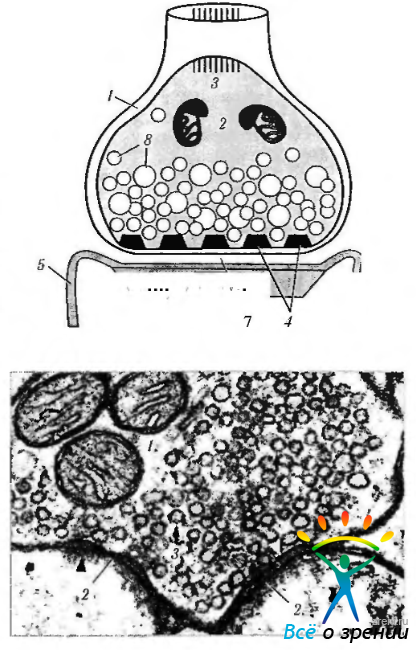
Рис. 1.4.33. Схема строения синапса (а) и его ультраструктурные особенности (б): а - схема строения синапса (1 - пресинаптическая часть; 2 -митохондрии; 3 — нейротрубочки и нейрофиламенты; 4 — пресинаптическая мембрана с пресинаптическими уплотнениями; 5 — постсинаптическая мембрана; 6 — постсинаптическое уплотнение; 7 — синаптическая щель; 8 — синаптические пузырьки); 6 — ультраструктурные особенности синапса (1 — митохондрии; 2 - постсинаптическое уплотнение; 3 — синаптические пузырьки)
Между параллельно расположенными пре- и постсинаптическими мембранами двух нейронов имеется пространство шириной 20—30 нм. называемое синаптической щелью. Пресинаптическая часть образуется аксоном по его ходу (приходящий синапс) или представляет собой расширенную конечную часть аксона (концевой бутон). В ней содержатся митохондрии, гладкая эндоплазматическая сеть, нейрофиламенты, нейротрубочки и синаптические пузырьки диаметром 20—65 нм, в которых находится нейромедиатор. Нейромедиаторы вырабатываются в теле клетки и механизмом быстрого транспорта переносятся в окончание аксона. На внутренней стороне плазмолеммы, обращенной к синаптической щели (пресинаптической мембраны), имеется пресинаптическое уплотнение, образованное фибриллярной гексагональной белковой сетью, ячейки которой способствуют равномерному распределению синаптических пузырьков по поверхности мембраны.
Синаптическая щель выполнена жидкостью, содержащей гликопротеиды, гликолипиды, а также электронноплотные частицы. Иногда она содержит гликопротеиновые интрасинаптические филаменты толщиной 5 нм, которые являются элементами специализированного гликокаликса, обеспечивающими адгезивные связи пре- и постсинаптической частей, а также направленную диффузию медиатора.
Постсинаптическая часть представлена постсинаптической мембраной, содержащей особые комплексы интегральных белков — синаптические рецепторы, связывающиеся с нейромедиатором. Мембрана утолщена за счет скопления под ней плотного филаментозного белкового материала (постсинаптическое утолщение).
Процесс передачи нервного импульса схематически представляется следующим образом. Поступление нервного импульса в окончание пресинаптического нейрона к синапсу проявляется в деполяризации пресинаптической мембраны, что делает мембрану проницаемой для ионов кальция. При этом ионы кальция начинают поступать в пресинаптическую цитоплазму. Это приводит к слиянию синаптических пузырьков, содержащих нейромедиатор, с пресинаптической мембраной. Слившиеся мембраны открываются в синаптическую щель, и содержимое пузырьков изливается туда. Медиатор, выделившийся в синаптическую щель, и воздействует на постсинаптическую мембрану, делая ее более проницаемой для ионов натрия, проникающих в цитоплазму постсинаптического нейрона. При этом потенциал покоя уменьшается и происходит деполяризация постсинаптической мембраны и генерируется нервный импульс. Подобный тип передачи нервного импульса характерен возбудительным нейронам. В тормозных нейронах высвобождение медиатора в синаптическую щель приводит к открытию каналов для ионов хлора, которые и переходят в цитоплазму постсинаптического окончания. При этом увеличивается ее отрицательный потенциал (гиперполяризация), а возбудимость клетки понижается. Таким образом, суммарное действие обоих видов синапсов на один нейрон приводит к определенному балансу между двумя противоположными видами синаптических эффектов. Медиаторами, опосредующими возбуждение, например, служат ацетилхолин и глютамат, а торможение опосредуется ГАМК и глицином.
После прекращения взаимодействия медиатора с рецепторами постсинаптической мембраны большая часть его эндоцитозом захватывается пресинаптической частью, меньшая рассеивается в пространстве и захватывается глиальными клетками. Некоторые медиаторы расщепляются ферментами на компоненты, которые далее захватываются пресинаптической частью. Затем они повторно используются для образования новых синаптических пузырьков.
В заключение необходимо привести также сведения относительно биохимической классификации нейронов. Основана эта классификация на химическом составе нейромедиатора того или иного нейрона. Выделяют
- холинергические (медиатор — ацетилхолин),
- адренергические (медиатор — норадреналин),
- серотонинергические (медиатор — серотонин),
- дофаминергические (медиатор — дофамин),
- ГАМК-ергические (медиатор — гамма-аминомасляная кислота),
- пуринергические (медиатор — АТФ и его производные),
- пептидергические (медиаторы — субстанция Р. энкефалины, эндорфины, вазоактивный интерстинальный пептид, холецистокенин, нейротензин, бомбезин и друтие нейропептиды) нейроны.
Вторую группу клеток нервной ткани составляют клетки нейроглии.
Нейроглия. Нейроглия — термин, обозначающий обширную группу клеток нервной ткани, обеспечивающих деятельность нейронов и выполняющих опорную, трофическую, разграничительную, барьерную, секреторную и защитную функции.
Морфологически различают три типа глиальных клеток —
- олигодендроциты,
- астроциты
- и микроглиальные клетки
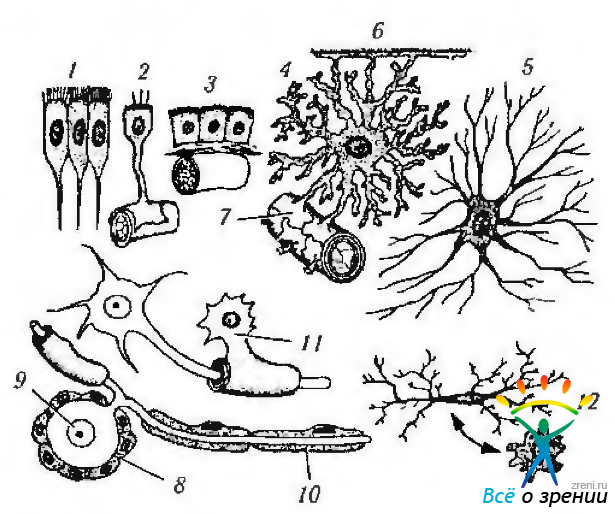
Рис. 1.4.34. Различные типы глиальных клеток нервной ткани (по В. Л. Быкову, 1999): 1 — эпендимоцит; 2 - таницит; 3 - хориоидный эпендимоцит; 4 — протоплазматический астроцит; 5 — волокнистый астроцит; 6 — пограничная глиальная мембрана мозга; 7—периваскулярная пограничная мембрана; 8 — клетки-сателлиты; 9 — нейрон; 10 — леммоцит (шванновская клетка); 11- олигодендроцит; 12 — микроглиальная клетка и ее трансформация в процессе фагоцитоза
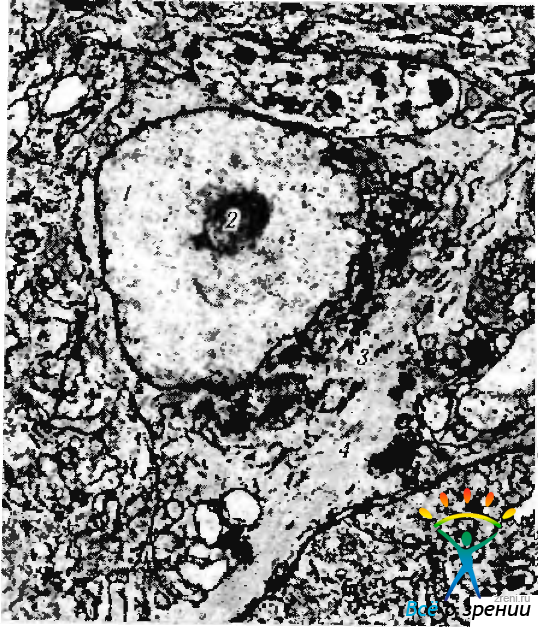
Рис. 1.4.36. Ультраструктурная организация глиальной клетки (олигодендроцит сетчатки): 1- ядро; 2 — ядрышко; 3 — цитоплазматические включения; 4—микрофибриллы
Олигодендроглия состоит из мелких клеток (олигодендроцитов) с короткими немногочисленными отростками. В цитоплазме клеток обнаруживается хорошо развитый синтетическии аппарат. Высокое содержание лизосом, митохондрий и гранул гликогена. В зависимости от особенностей строения олигодендроциты разделяют на крупные светлые, мелкие темные и промежуточные. У взрослых обнаруживаются преимущественно мелкие темные клетки. Длительность существования светлой клетки невелика. и она постепенно вызревает, превращаясь в малый темный олигодендроцит.
Одной из наиболее важных функций олигодендроцитов является образование миелиновой оболочки вокруг нервных волокон в центральной нервной системе. Миелин, выполняя функцию изолятора, участвует в процессе передачи по волокну нервного импульса.
Процесс миелинизации сводится к тому, что олигодендроцит обертывает своей цитоплазмой определенный участок нервного волокна последовательными спиральными слоями (рис. 1.4.37—1.4.39).

Рис. 1.4.37. Взаимоотношение олигодендроцита с нервным волокном и строение миелинизированного волокна (по Шаде, Форд, 1976): 1 — олигодендроцит; 2 — аксон; 3 — связь между телом клетки глии и миелиновой оболочкой; 4 — цитоплазма- 5 — цитоплазматическая мембрана; 6 — перехват Ранвье; 7 — петля плазматической мембраны; 8- мезаксон; 9 — гребешок

Рис. 1.4.38. Механизм формирования миелиновой оболочки шванновскими клетками (в периферической нервной системе) или олигодендроцитами (в центральной нервной системе)
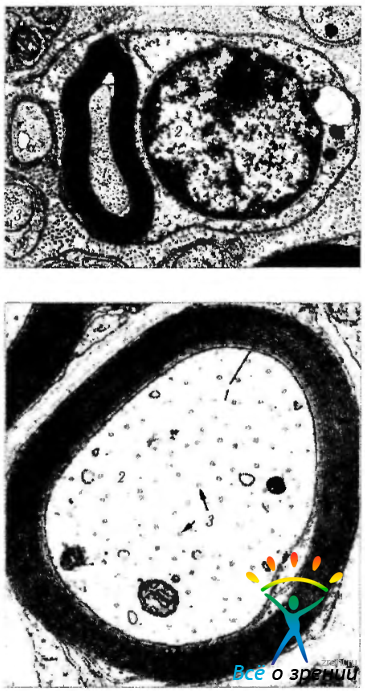
Рис. 1.4.39. Ультраструктурные особенности шванновской клетки (а) и миелиновой оболочки периферического нерва (а, б): а отношение олигодендроцита к аксону (1 -аксон; 2 ядро олигодендроцита; 3—немиелинизированный нервный ствол); б— миелиновая оболочка аксона (1—миелиновая оболочка; 2— аксон; 3 — микротрубочки)
В результате этого нервное волокно оказывается окутанным целым пакетом двухслойных цитоплазматических мембран, содержащих миелин. В результате этого и наступает изоляция нервного волокна. В миелинизации периферических нервов участвует шванновская клетка (леммоцит) (см. ниже).
Астроглия представлена астроцитами (см. рис. 1.4.36). Астроциты обладают многочисленными отростками, расходящимися от тела клетки в разных направлениях, напоминая при этом звезды.
Характеризуется клетка наличием светлого овального ядра. Ее цитоплазма содержит небольшое количество органоидов, но значительное количество зерен гликогена и промежуточных филаментов. Промежуточные филаменты содержат особый глиальный фибриллярный кислый белок (ГФКБ), который служит маркером астроцитов. Астроциты образуют щелевые соединения между собой, а также с клетками олигодендроглии и эпендимной глии.
Разделяют астроциты на
- волокнистые (фибриллярные)
- и прототазматические (плазматические).
Ультраструктурная организация этих клеток приблизительно одинаковая. Пучки филаментов распространяются от одного отростка к другому, проходя через тело клетки, что создает ее особую жесткость.
Протоплазматические астроциты преимущественно встречаются в сером веществе центральной нервной системы, а волокнистые — в белом. Кроме того, волокнистые астроциты содержат большое количество ГФКБ.
Учитывая то, что клетки астроглии плотно контактируют с сосудами и нейронами, предполагают, что эти клетки выполняют опорную, разграничительную, транспортную, барьерную, трофическую, защитную и регуляторную функции.
Опорная функция сводится к формированию астроцитами каркаса, внутри которого располагаются нейроны и волокна. В ходе эмбрионального развития они служат опорными и направляющими элементами, вдоль которых происходит миграция нейронов. Направляющая функция связана также с секрецией ростовых факторов, распознаваемых нейронами.
Разграничительная, транспортная и барьерная функции астроглии сводятся к участию в образовании гемато-энцефалического и лиферативная активность способствует также глиальному рубцеванию поврежденных нервных тканей.
Микроглия — это мелкие клетки, разбросанные в белом и сером веществе мозга. Они составляют всего 5% популяции глиальных элементов. Микроглия обнаруживается и в сетчатой оболочке. Предполагают, что схожие по происхождению и функции клетки лежат в стекловидном теле вблизи сетчатки.
Считается, что микроглиальные клетки происходят из моноцитов или периваскулярных макрофагов мозга (мезенхимное происхождение). Структурной особенностью микроглии является насыщенность цитоплазмы лизосомами. Вероятней всего микроглия определяет защитную функцию в нервной системе, относясь к клеткам системы иммунитета. При патологических состояниях микроглиальные клетки проявляют способность к передвижению, фагоцитозу. Их количество существенно увеличивается при воспалительных и дегенеративных заболеваниях нервной ткани. При этом они утрачивают отростки, округляются и способны фагоцитировать остатки погибших клеток. При повышении активности микроглиальных клеток усиливается секреция ряда цитокинов и токсических радикалов. Именно с этим связывают усиленную гибель нейронов путем апоптоза при некоторых заболеваниях нервной системы.
К глиальным элементам относят и эпендимную глию (эпендима). Клетки эпендимной глии выстилают желудочки мозга и спинномозговой канал. К эпендимной глие ряд авторов относит и плоские клетки, выстилающие мозговые оболочки (менинготелий).
Особенностью эпендимоцитов является наличие на апикальной поверхности ресничек, которые при своем движении перемещают спинномозговую жидкость. Клетки эпендимной глии плотно прикрепляются друг к другу межклеточными соединениями. Часть клеток лежит на базальной мембране. Некоторые клетки от базальной своей поверхности отдают отростки по направлению мозга, входящие в состав поверхностной пограничной глиальной мембраны (краевая глия).
Эпендимная глия выполняет опорную, трофическую, барьерную и секреторную функции. Барьерная функция эпендимной глии сводится к участию в образовании нейро-ликворного и гемато-ликворного барьеров.
Покрывает эпендима и сосудистые сплетения мозга (специализированными клетками — хориоидными эпендимоцитами и таницитами).
Завершая описание строения структурных элементов центральной нервной системы, необходимо упомянуть и о нейропиле, т.е. структурном компоненте центральной нервной системы, представляющем собой при световой микроскопии светло-голубой материал, в который погружены нейроны, их отростки, капиллярные сосуды. Ультраструктурно показано, что нейропиль представляет собой переплетение клеточных тел, отростков нейронов и глиальных элементов.
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0