Движения глаз │ Часть 1


Содержание:
Описание
↑ Движения глаз и их нейронный контроль
Движения глаз являются прямым следствием функционирования наружных мышц глаза. В предыдущих главах нами подробно описано макро- и микроскопическое строение наружных мышц глаза, а также особенности их прикрепления к глазному яблоку. Обращено также внимание на характер взаимоотношения мышц с соединительнотканными образованиями глазницы. Задачей настоящего раздела является изложение современных сведений о движении глаз, контроле этих движений нервной системой, что немаловажно знать офтальмологу для правильной оценки возможных нарушений этой функции.
Необходимо отметить, что литература, посвященная изложению основных закономерностей движения глаз, очень обширная. Она включает в себя как монографии, так и многочисленные статьи в специализированных журналах по нейроофтальмологии и физиологии. Учитывая специфику настоящей книги, мы попытались изложить основные сведения относительно движения глаз, делая упор на морфологические основы нейронного управления этим процессом.
Первоначально необходимо определить некоторые общие понятия движения глаз, имеющие большое значение не только в понимании механики движения, но и являющиеся отправными точками при рассмотрении вопросов нейронного контроля.
Одним из основных понятий является понятие «центр вращения» глаза. Глазное яблоко можно уподобить сфере, вращающейся в глазнице вокруг определенной точки, расположенной приблизительно в центре. Эту точку и называют центром вращения. Необходимо отметить, что центр вращения глаза отличается от анатомического центра. Точно определить его не представляется возможным. Расположение центра вращения зависит от положения глаза в глазнице, анатомических особенностей костных стенок и тканей глазницы, а также степени раскрытия глазной щели. Немаловажно и положение головы. По этой причине выделяют скопление точек, называемое центроидом (рис 4.4.1).
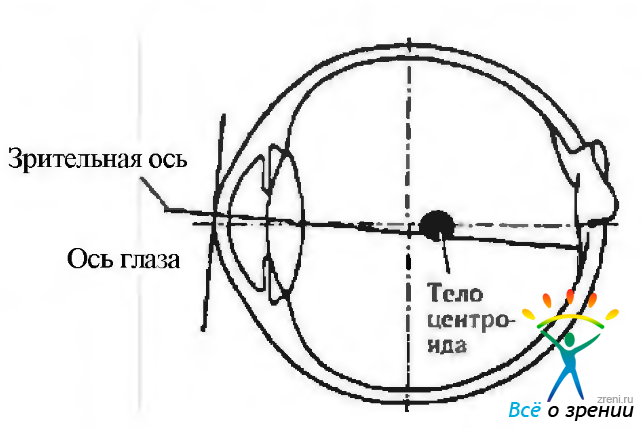
Рис. 4.4.1. Расположение пространственного центроида глазного яблока (по Park, Park, 1933)
Изменение координат при движении глаза оценивают по его смещению относительно точки, расположенной в 13,5 мм от верхушки роговицы и в 1,6 мм в назальную сторону от геометрического центра глаза. При этом используют плоскость Листинга, которая представляет собой фронтальную плоскость, проходящую через центр вращения глаза. В первичном положении глазных яблок плоскость Листинга совпадает с экваториальной плоскостью.
Движение глаза можно описать относительно системы координат с тремя перпендикулярно расположенными осями относительно друг друга, которые пересекаются в центре вращения глаза. Это оси X, Y и Z (рис. 4.4.2).
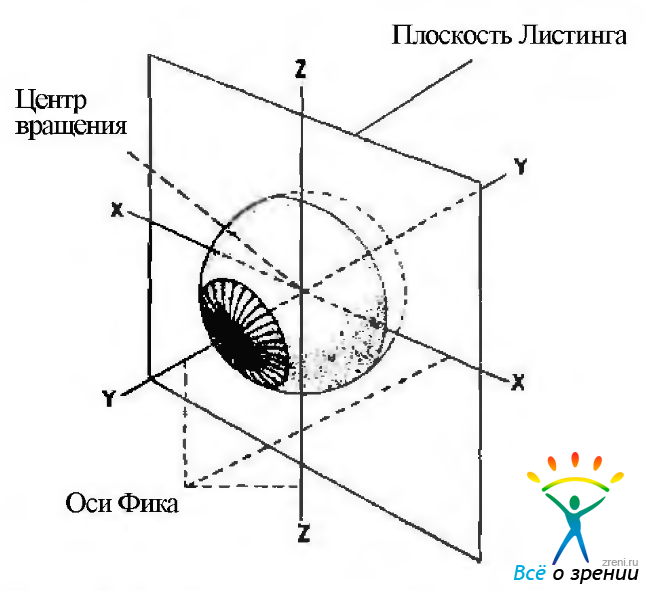
Рис. 4.4.2. Оси и плоскость вращения глазного яблока
Оси X и Z располагаются в плоскости Листинга. Ось Y представляет собой перпендикуляр, восстановленный к плоскости Листинга из центра вращения глаза.
Вращение глаза в горизонтальной плоскости (вокруг оси Z), при котором передний полюс глаза смещается назально (медиально), называется приведением (аддукция), если передний полюс глаза смещается в височную сторону (латерально) — отведением (абдукция).
Вращение в вертикальной плоскости (вращение относительно оси X), сопровождающееся смещением переднего полюса глаза кверху, называется поднятием (супрадукция, элевация; поворот одного глаза кверху), а если книзу — опущением (интрадукция, депрессия) (рис. 4.4.3).
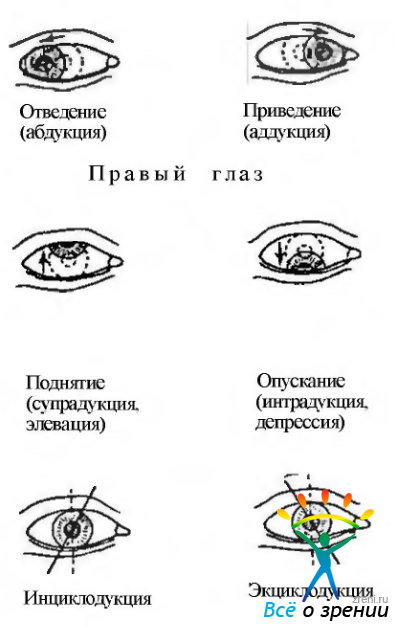
Рис. 4.4.3. Виды движения глазного яблока: а — горизонтальные движения вокруг вертикальной оси (ось Z); б — вертикальные движения вокруг горизонтальной оси (ось X); в — третичное положение глаз, вызываемое одновременным вращением вокруг горизонтальной и вертикальной осей
Мышечная плоскость представляет собой плоскость, проходящую через центр вращения глаза и через длинник мышцы от ее места прикрепления к глазу и глазнице. Она определяет направление действия мышцы.
Ось вращения мышечной плоскости является линией, проведенной перпендикулярно мышечной плоскости через центр вращения. При сокращении мышцы она является и осью вращения глаза.
Первичное положение глаза. Первичное положение — это такое положение глаза, от которого ведется определение типа вращения глаза и измерение движений. Scobee определил первичное положение как «...положение глаза при бинокулярном зрении с вертикально поднятой головой. При этом объект располагается в бесконечности и в пересечении сагиттальной и горизонтальной плоскостей, проходящих через центры вращения двух глазных яблок».
Вторичное положение глаза. Вторичным положением считается вращение глаза исключительно вокруг горизонтальной или вертикальной осей. Вращения глаза вокруг оси Y при этом не наблюдается (рис. 4.4.3, а, б).
Третичное положение глаза. Наклонные положения глаза называются третичными позициями. Третичные позиции возникают при одновременном вращении глаза вокруг горизонтальных и вертикальных осей (кверху вправо, книзу вправо, кверху влево, книзу влево) (рис. 4.4.4).
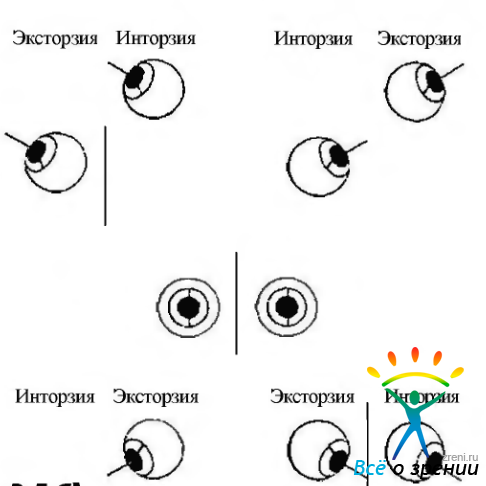
Рис. 4.4.4. Одновременное движение глаз из первичного положения относительно осей X и Z приводит к псевдоторзии (по Adler, 1985)
Это смещение называется ложным вращением, поскольку никакого вращения вокруг оси Y нет.
Вращение (торзия). Истинное вращение происходит тогда, когда глаз поворачивается относительно оси, расположенной в сагиттальной плоскости глаза, и относительно осей Фика (рис. 4.4.2). Если верхний конец вертикального сечения наклонен в назальную сторону, состояние называется инциклодукцией, а если к виску — эксциклодукцией (рис. 4.4.3, в; 4.4.5).
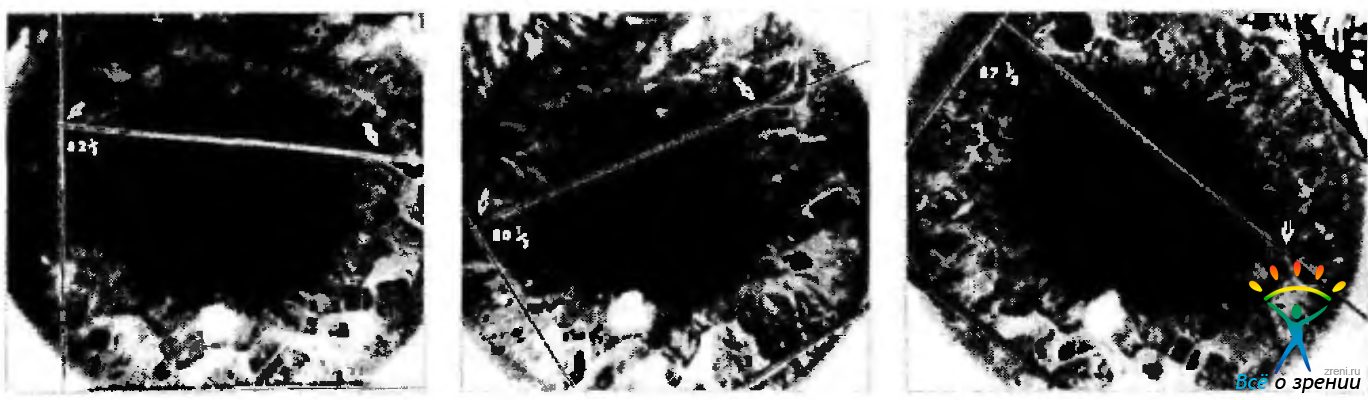
Рис. 4.4.5. Истинные торзионные движения глаз (по Linwong, Herman, 1971): Правый глаз. Нижняя линия проходит поперек глаза. Вторая линия проходит перпендикулярно первой. Верхняя соединяет две маркерные точки (стрелки) на радужке и формирует угол с перпендикулярной линией. Путем измерения угла между двумя линиями выявляется, что наклон головы вправо (второй рис.) приводит к появлению инциклодукции, равной 2 градусам. При повороте головы влево (третий рис.) развивается эксциклодукция. равная 5 градусам
Торзионные движения происходят в небольшом объеме. Так, при наклоне головы на 30° происходит инциклодукция ипсилатерального глаза на 7,0±3,1° и эксциклодукция на 8,36 + 2,5° контрлатерального глаза.
↑ Модели функции наружных мышц глаза
Независимое действие любой наружной мышцы глаза можно рассматривать только теоретически, поскольку в реальности для любого движения глаза необходимо одновременное сокращение всех шести мышц.
В связи со сложностью подробного описания движений глаза были разработаны многочисленные упрощенные модели. Чем более детальная модель, тем более она реально описывает процесс.
Ниже представлено три модели. Модель «синергистов» («агонистов») самая простая и получила наибольшее распространение в клинике. Вторая, более сложная, модель описана Boeder. Это так называемая модель «пар антагонистов», учитывающая вклад в движения глаз вторичного действия мышц.
Robinson использовал экспериментальные данные и обработал их при помощи компьютера. На этой основе он разработал «количественную модель», которая позволяет вычислить относительные вклады всех мышц при любом положении глаза.
Модель «агонистов». Традиционно действие каждой в отдельности мышцы рассматривают с позиций Duane. Он предположил, что вертикальные противолежащие мышцы являются синергистами в определенном положении глаз. При этом их функции изменяются в процессе изменения положения глаз (например, при нарастании абдукции косые мышцы вызывают вращение глаза и еще большую абдукцию). Эта модель применяется в клинике с начала 30-х годов прошлого века до настоящего времени. Первичное и вторичное действие отдельных мышц глаза суммировано в табл. 4.4.1.
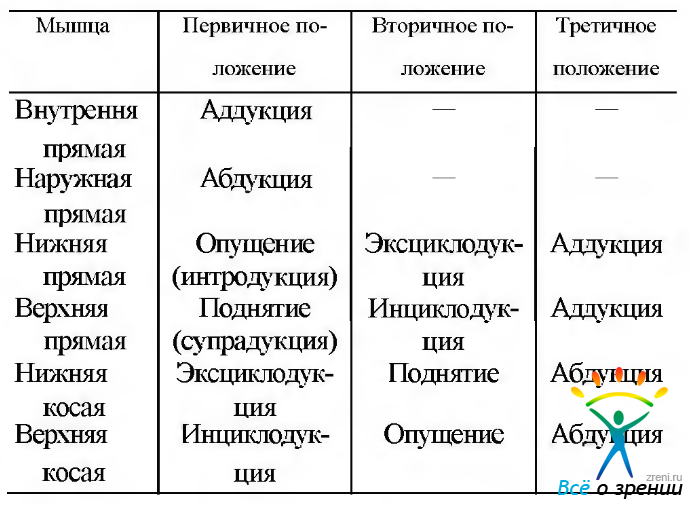
Таблица 4.4.1. Действие наружных мышц глаза
В соответствии с этой моделью действие отдельных наружных мышц приводит к следующим эффектам.
Внутренняя прямая мышца (рис. 4.4.6, а).

Рис. 4.4.6: а—действие внутренней прямой мышцы (аддукция); б — действие наружной прямой мышцы (абдукция). Функционирующая мышца окрашена более интенсивно
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Мышечная плоскость внутренней прямой мышцы располагается точно в горизонтальной плоскости глазного яблока. По этой причине, когда глазное яблоко находится в первичном положении, ее сокращение приводит только к вращению глаза кнутри (приведение, аддукция). Когда зрительная ось направлена выше горизонтальной линии, сокращение мышцы способствует еще большему поднятию глаза. В тех случаях, когда зрительная ось направлена ниже горизонтальной плоскости, внутренняя прямая мышца опускает глазное яблоко.
Наружная прямая мышца (рис. 4.4.6, б). Наружная прямая мышца расположена в той же плоскости, что и внутренняя. По этой причине. когда глаз находится в первичном положении, сокращение ее приводит только к отведению глаза (абдукция). Если же глазное яблоко приподнято или опущено, сокращение мышцы еще более поднимает или опускает его.
Верхняя прямая мышца (рис. 4.4.7).
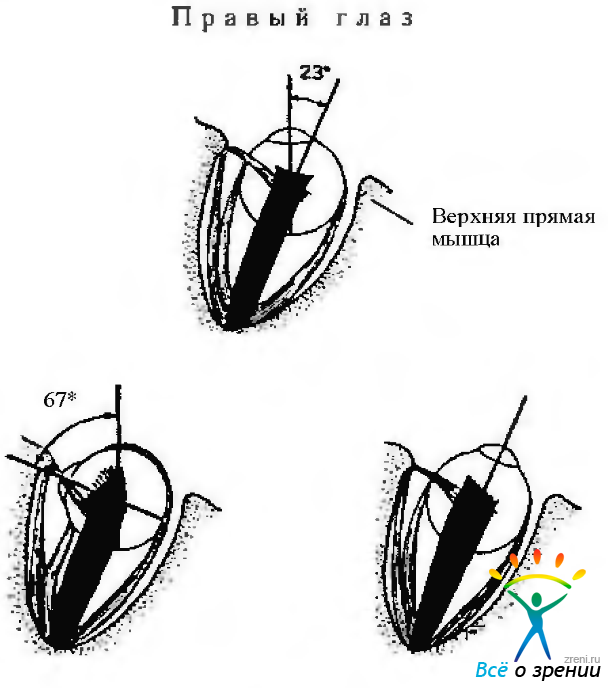
Рис. 4.4.7. Верхняя прямая мышца: а — когда глаз находится в первичном положении, плоскость мышцы расположена под углом 23 градуса относительно зрительной оси. В этом положении мышца поднимает глаз. Вторичным ее действием является инциклодукция и аддукция; б — если глаз отведен, мышца в меньшей степени поднимает глаз. При этом она и в большей степени является аддуктором и инциклодукто-ром. При отведении глаза на 67 градусов мышца становится исключительно инциклодуктором; в — при абдукции глаза на 23 градуса верхняя прямая мышца становится чистым элеватором. При этом плоскость мышцы совпадает со зрительной осью
Верхняя прямая мышца находится относительно зрительной оси в первичном положении под углом 23—25°. Первичным действием этой мышцы является поднятие (элевация, супрадукция) глаза, а вторичным — приведение (аддукция) и инторзия (инциклодукция). По этой причине сокращение мышцы приводит к вращению глаза вокруг нескольких осей.
В тех случаях, когда зрительная ось направлена в горизонтальной плоскости наружу под углом 23° от первичного положения, сокращение верхней прямой мышцы приводит только к поднятию глаза. Если глаз отклонен на 67° в назальную сторону от первичного положения так, что зрительная ось располагается под прямым углом к плоскости мышцы, сокращение мышцы приводит только к приведению (аддукция) и инторзии (инциклодукция). При параличе верхней прямой мышцы отведенный глаз не может быть поднят.
Нижняя прямая мышца (рис. 4.4.8).
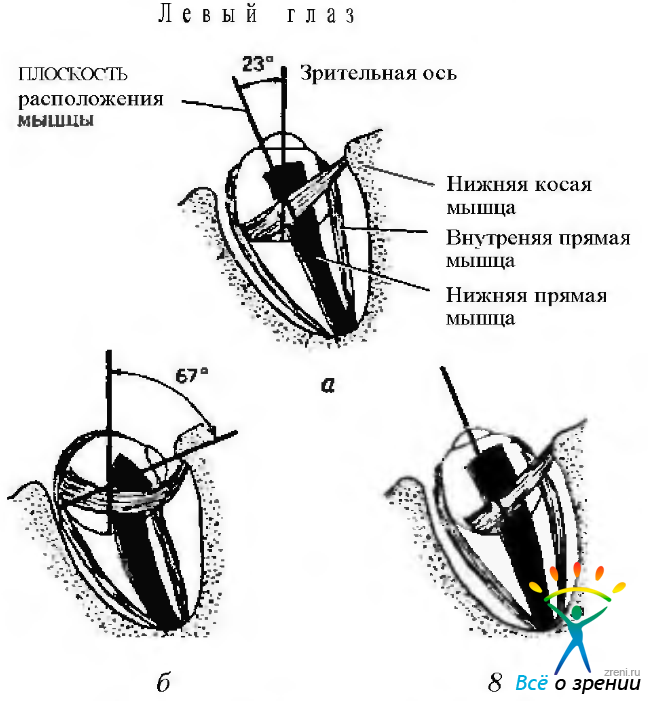
Рис. 4.4.8. Нижняя прямая мышца: а — в первичном положении нижняя прямая мышца формирует угол со зрительной осью, равный 23 градусам. Этот угол аналогичен углу, образованному между зрительной осью и верхней прямой мышцей. При расположении глаза в первичном положении мышца опускает глазное яблоко (депрессия). Вторичным ее действием являются экспиклодукция и аддукция; б — если глаз отведен, нижняя прямая мышца в меньшей степени опускает глаз, но в большей степени отводит его. Развивается и эксциклодукция; в — если глаз приведен на 23 градуса, мышца опускает глазное яблоко (депрессия)
Нижняя прямая мышца располагается в той же плоскости, что и верхняя прямая. По этой причине, когда глаз находится в первичном положении, действие мышцы при сокращении аналогично таковому при сокращении верхней прямой мышцы. В результате сокращения происходит вращение глаза, состоящее из его опущения (депрессия), а также вторичного приведения (аддукция) и эксциклоторзии.
Верхняя косая мышца (рис. 4.4.9).
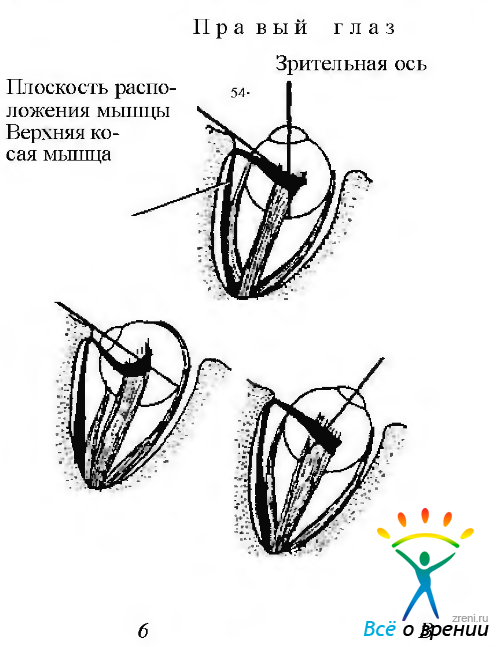
Рис. 4.4.9. Верхняя косая мышца: a — в тех случаях, когда глаз находится в первичном положении, плоскость мышцы формирует угол со зрительной осью, равный 54 градусам. При этом основной функцией мышцы является инциклодукция. Вторичное действие мышцы сводится к аддукции и опущению глазного яблока (депрессия); б — когда глаз отведен на 54 градуса, зрительная ось совпадает с плоскостью мышцы. В этом положении глаза мышца действует как инциклодуктор. При этом ее вертикальное действие становится более выраженным; в — когда глаз отведен, верхняя косая мышца действует как инциклодуктор и абдуктор
При сокращении верхней косой мышцы в тех случаях, когда глаз находится в первичном положении, происходит сложное вращение глаза. Состоит это вращение из трех компонентов: первичное действие — инторзия (инциклодукция), а вторичное — опущение (депрессия) и приведение (аддукция). Когда глаз развернут под углом 51° в назальную сторону относительно первичного положения, зрительная ось параллельна напряжению мышцы и сокращение ее приводит к опущению (депрессии) глаза. Если глаз направлен в височную сторону на 39° относительно своего первичного положения так, что зрительная ось располагается под прямым углом к напряжению мышцы, сокращение ее приводит к инторзии (инциклодукция) и отведению (абдукция).
Нижняя косая мышца (рис. 4.4.10).
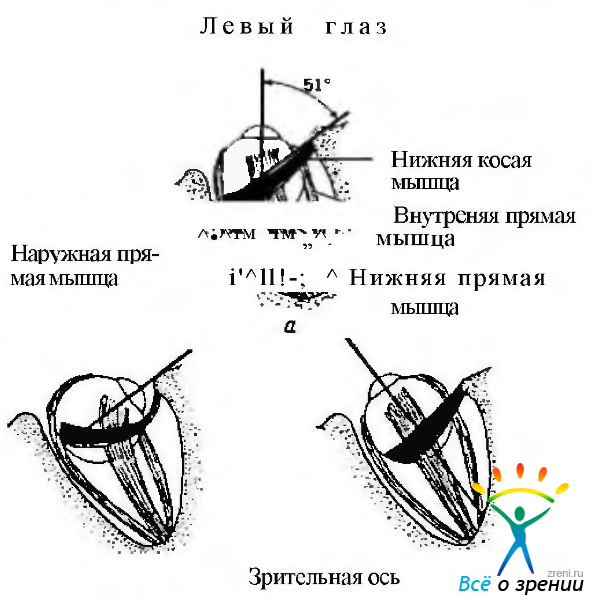
Рис. 4.4.10. Нижняя косая мышца: а — когда глаз находится в первичном положении, нижняя косая мышца формирует со зрительной осью угол, равный 51 градусу. В этом положении основной функцией мышцы является эксциклодукция, а вторичной — абдукция и элевация; б—когда глаз отведен на 51 градус, зрительная ось располагается в плоскости мышцы. В таком состоянии нижняя косая мышца действует только как эксциклодуктор. При этом функция супрадук-ции становится более выраженной; в — при абдукции первичным действием мышцы является эксциклодукция, а вторичным — аддукция
Плоскость мышцы располагается под углом 51° относительно зрительной оси при первичном положении глаза. По этой причине сокращение нижней косой мышцы приводит к сложному движению глаза, состоящему из трех компонентов: первичное действие — поворот кнаружи (абдукция), а вторичное — приведение (аддукция) и поднятие (элевация).
Когда глаз смещен на 39 в височную сторону от своего первичного положения, сокращение нижней косой мышцы приводит к повороту его кнаружи и эксциклодукции. Когда глаз направлен на 51° в назальную сторону, сокращение мышцы приводит к его поднятию (элевации) и эксциклодукции.
Естественно, что мышцы действуют совместно. По этой причине разработаны модели содружественного их действия.
Модель пар «антагонистов». Одновременный анализ действия шести наружных мышц был упрощен Boeder с выделением так называемых пар антагонистов. Пары антагонистов для правого и левого глаз приведены в табл. 4.4.2.
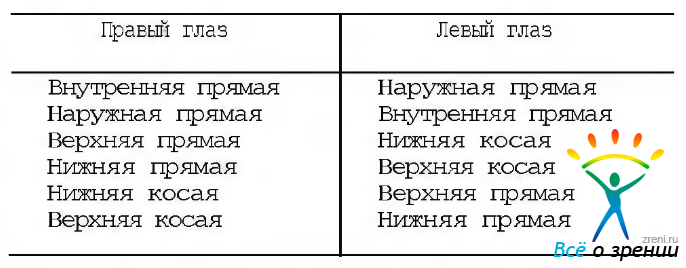
Таблица 4.4.2. Пары антагонистов наружных мышц глаза
Boeder считает, что сокращение одного члена пары антагонистов сопровождается удлинением противоположной мышцы. С механической точки зрения, наблюдаемое удлинение должно соответствовать степени сокращения антагониста. Когда функционируют мышцы пары одновременно, глаз располагается на пол-пути между двумя мышцами. При анализе действия пар мышц Boeder использует сферическую систему координат. В соответствии с этой моделью действие пар мышц рассматривается следующим образом.
Пары вертикальных прямых мышц. Первичным действием вертикальных прямых мышц является поднятие и опущение глаза, а вторичным — приведение и вращение. Имеется также относительная абдукция при расположении глаза несколько кверху или книзу.
Пары косых мышц. Первичным действием пары косых мышц является инторзия и поворот кнаружи. Инторзия не является вращением вокруг зрительной оси или оси Y, а, скорее, представляет собой движение относительно определенной точки на лимбе.
Из-за стабильности вращательной оси в глазнице сокращение одной из косых мышц (с расслаблением антагониста) приводит к вращению глаза вокруг оси, независимо от линии закрепления в горизонтальной плоскости. Вторичным действием пары косых антагонистов является перемещение глаза в горизонтальной и вертикальной плоскостях с увеличивающимся приведением глаза.
Пары горизонтальных мышц. Первичным действием горизонтальных антагонистов является приведение (аддукция) и отведение (абдукция). Вторичное их действие минимально.
При поднятом начальном положении отмечается поднятие (элевация) и эксциклодукция при приведении и инциклодукция при абдукции. Противоположная картина выявляется при опущенном глазном яблоке.
Использование концепции пар мышц позволило Boeder вычислить изменение длины мышц для каждого положения глаза.
Существуют и так называемые «количественные» модели, основанные на использовании компьютерной техники. Клиническая ценность этих более сложных моделей движения глаза пока не установлена.
Дав краткую характеристику строения наружных мышц глаза, их иннервации, мы остановимся на описании типов движения глаз и нейронном контроле этих движений. Но перед этим имеет смысл остановиться на значении в движении глаз взаимодействия между мышцами глаза и окружающими его соединительнотканными образованиями. Следует подчеркнуть, что движения глаз являются следствием взаимодействия статических и динамических сил. Для правильного понимания значения мышц в движении глаз необходимо сначала определить, каким образом ткани глазницы сопротивляются движениям. В первом приближении глаз в глазнице можно рассматривать в виде шара, подвешенного в сложной системе «эластических тяжей», обладающих тенденцией приводить его в центральное положение. Для смещения глаза (выведение его в эксцентричное положение) мышцы должны развить статическую силу, адекватно противодействующую соединительнотканным тяжам. Для перемещения глаза из одного эксцентричного положения в другое требуется дополнительная динамическая сила, которая способна преодолевать сопротивление тканей глазницы. Учитывая то, что статическая сила должна быть приложена к глазу постоянно для поддержания его положения в стационарном положении, динамическое усилие прилагается только во время движения глаза. Это динамическое усилие развивается при «разряжении» так называемых «физических» («пульсовых») мотонейронов. По это причине, основной задачей при дальнейшем изложении закономерностей контроля движений глаз центральной нервной системой необходимо определить нейроны, управляющие статическими и динамическими усилиями мышц. Возможно это только при использовании экспериментальных методов определения активности нейронов как в спокойном положении глаза, так и во время его движения. Благодаря этим исследованиям установлено, что в течение движения глаз одна пара мышц антагонистов сокращается благодаря активности «физических» нейронов, а вторая пара мышц расслабляется в результате торможения соответствующих им нейронов. Эта активность приводит глаз в новое статическое положение, поддерживаемое «тонической» активностью глазодвигательных нейронов. Таким образом, функцией глазодвигательных нейронов является постоянное вычисление необходимой активности нейронов для перемещения глаза на определенное расстояние, а также фиксация глаза в этом положении. При этом основной задачей системы является стабилизация взгляда, преобразуя сенсорную информацию различной модальности (зрительную, вестибулярную, слуховую и соматосенсорную) путем вычисления степени сокращения мышцы (сила и длина сокращения) — «пульс-шаг».
После изложения основных принципов движения глаз можно остановиться и на нейронном контроле движений глаза.
↑ Нейронный контроль движений глаз
Контроль над положением глаз и координация сокращения наружных мышц глаза интенсивно изучались и изучаются до настоящего времени при помощи клинических, анатомических, физиологических и биоинженерных методов исследования. Интеграция данных этих дисциплин способствовала пониманию деятельности глазодвигательной системы животных и человека.
Основными функциями нейронного контроля движений глаза является обеспечение направления глаза к зрительной цели, поддержание функции бинокулярного зрения, пространственная локализация зрительной цели. Происходит это благодаря доставке центростремительной информации коре головного мозга сведений относительно положения глаза в глазнице, что, в свою очередь, помогает определить направление взора.
Афферентная система. Система, регулирующая движения глаза, состоит из приводящей части (афферентная система), передающей сенсорную информацию относительно положения глаза, а также отводящей части (эфферентная система), ответственной за его движение.
Чувствительные волокна, идущие от наружных мышц глаза в составе III, IV и VI нервов, направляются к нейронам, расположенным в ядре среднемозгового тракта тройничного нерва. Конечным пунктом передачи информации являются чувствительные ядра тройничного нерва, а тажrе вентральное основное ядро зрительного бугра.
Информация к перечисленным структурам нервной системы поступает от многочисленных механорецепторов, залегающих в паренхиме наружных мышц глаза.
К основным механорецепторам относятся:
- мышечное веретено,
- сухожильный аппарат Гольджи,
- палисадоподобные и спиральные нервные окончания.

Таблица 4.4.3. Чувствительный аппарат наружных мышц глаза человека
Мышечное веретено. В наружных мышцах глаза мышечное веретено обнаружено как у животных, так и у человека. Мышечное веретено наружных мышц глаза отличается от таковых скелетной мускулатуры.
Основной функцией этой структуры является точная регистрация положения глазного яблока в глазнице и передача этой информации вышерасположенным нервным центрам.
Мышечное веретено распределено неравномерно и чаще обнаруживается в узкой части мышцы. Число этих рецепторов в одной мышце колеблется от 22 до 71. Внутренняя прямая мышца глаза содержит 18,8 мышечного веретена; наружная прямая—19,3; нижняя прямая — 34,0; верхняя косая — 27,3; нижняя косая — 4,3.
Каждое веретено содержит в капсуле 1 — 15 интрафузальных волокон, а на полюсе обнаруживается концевая пластинка. В области экватора рецептора располагается множество мякотных и безмякотных нервных волокон, окружающих интрафузальное волокно. Толщина интрафузальных волокон различна и колеблется от 5 до 30 мкм (в среднем 10 мкм). Длина веретена равняется 350—725 мкм (в среднем — 500 мкм).
Рядом исследователей установлено, что мышечное веретено не регистрирует положение глазного яблока при отсутствии зрительного стимула. Fuchs, Kornhuber предполагают, что веретено поставляет информацию мозжечку относительно длины и конечного пункта саккадического движения глаза.
Сухожильный аппарат Гольджи регистрирует степень регидности мышцы. Длина каждой отдельной структуры равна примерно 1 мм, а ширина — 0,1 мм. В каждом образовании выявляется 15—20 экстрафузальных волокон. Капсула состоит из сплетения коллагеновых волокон, между которыми проходят приводящие аксоны.
Сокращение мышцы сжимает аксоны, в результате чего формируются сигналы, свидетельствующие об увеличении ригидности мышцы.
Палисадоподобные окончания. Палисадоподобные окончания (мышечно-сухожильные цилиндры) являются основным сенсорным аппаратом наружных мышц глаза млекопитающих. Они состоят из «скрытых» нервных окончаний, находящихся в месте перехода мышечной ткани в сухожилие. Эти образования представляют собой нежную сеть нервных волокон, контактирующих с мышечным волокном. Один мякотный аксон может отдавать ветви нескольким соседним мышечным волокнам. В большем количестве они обнаружены в горизонтальных прямых мышцах глаза. Нервный сигнал формируется в результате сжатия терминалов при сокращении мышцы.
Спиральные окончания. В средней трети наружных мышц глаза выявляются нервные волокна, спиралевидно окутывающие мышечные волокна. Они делают 3—8 поворотов вокруг мышечного волокна и заканчиваются структурой, напоминающей концевую пластинку.
Тракты проприорецепции. Проприорецептивные импульсы от наружных мышц глаза проходят через глазную ветвь тройничного нерва к полулунному ганглию, обладающему соматотопической организацией (рис. 4.4.11)
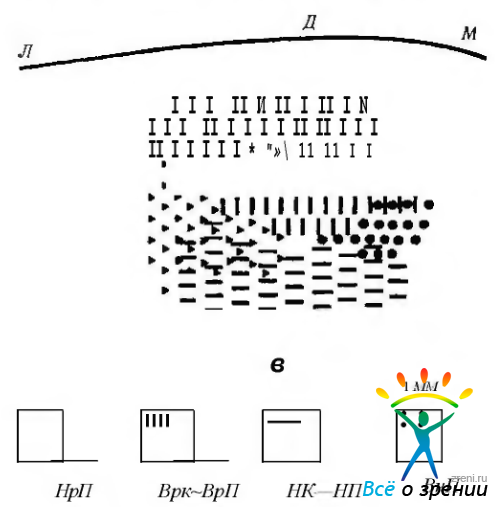
Рис. 4.4.11. Диаграмма, иллюстрирующая соматотопическую организацию афферентов проприорецептивной чувствительности наружных мышц глаза в полулунном ганглии (по Е. Manni, Е. V. Petrossi): Л — латеральная; Д — дорзальная; М — медиальная; В— вентральная поверхности полулунного ганглия; НрП—наружная прямая мышца; ВрК — верхняя косая мышца; ВрП—верхняя прямая мышца; НК—нижняя косая мышца; НП — нижняя прямая мышца; ВнП—внутренняя прямая мышца
Центральные отростки нейронов первого порядка заканчиваются в ипсилатеральном чувствительном ядре тройничного нерва. Они также проецируются на более каудальную часть ядра тройничного нерва.
От ядра тройничного нерва проприорецептивные импульсы передаются крыше и покрышке среднего мозга и посредством медиальной петли (lemniscus medialis) поступают к вентрально расположенному комплексу ядер зрительного бугра. К другим путям, вовлеченным в анализ проприорецептивной информации, относятся наружное коленчатое тело, зрительная кора головного мозга. Проприорецептивные волокна от наружных мышц глаза подходят также к V, VI и VII долям коры мозжечка посредством мшистых волокон. Участвуют в обработке информации и верхние холмики четверохолмия. При этом проприорецептивные афференты к верхнему четверохолмию обладают тормозным влиянием.
Эфферентная система. Цель глазодвигательной системы состоит в том, чтобы навести глаза на выбранный объект наиболее эффективным и скоординированным способом. В предыдущих главах нами показано, что степень остроты зрения, в определенной мере, определяется плотностью расположения фоторецепторов в специализированной части сетчатой оболочки, называемой центральной ямкой. Естественно, что в эволюционном развитии параллельно с формированием центрального зрения развивалась и система наведения изображения именно на этот наиболее чувствительный участок сетчатки. Раньше развились системы, позволяющие фиксировать неподвижное изображение. Подобный тип фиксации взора характерен для низших позвоночных. У более сложно организованных животных появилась необходимость следить за движущимися целями и при этом фиксировать изображение на наиболее чувствительный участок сетчатой оболочки. Подобный тип фиксации взора характерен для высших млекопитающих, включая человека. Благодаря столь сложному эволюционному развитию системы слежения за зрительной целью и сформировалось пять систем, функционирующих содружественно и дополняющих одна другую. Это саккадная система, система плавного слежения, вергентная система, система обеспечения фиксации и вестибулярная система. Функционирование первых четырех систем зависят от информации, получаемой от органа зрения, в то время как формирование последней (вестибулярной) зависит от информации, получаемой от вестибулярных и отолитовых рецепторов, рецепторов шеи и, возможно, от наружных мышц глаза. Информация от всех перечисленных систем объединяется, обрабатывается и направляется к ядрам, управляющим наружными мышцами глаза (табл. 4.4.4).
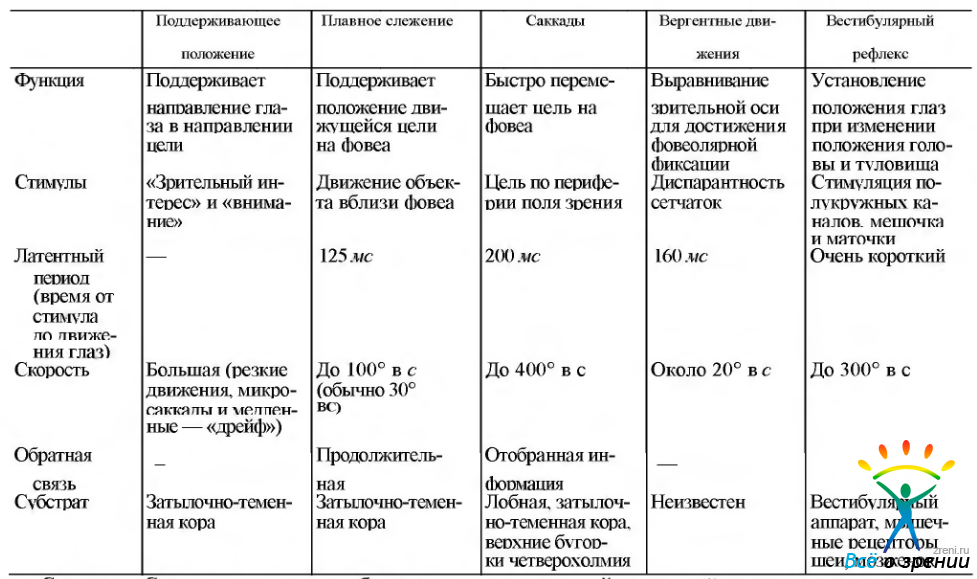
Таблица 4.4.4. Характеристика различных типов движения глаз
Саккады. Саккадная система обеспечивает обнаружение зрительной цели и выведение ее на наиболее чувствительную часть сетчатой оболочки (центральная ямка), исправляя при этом существующую «угловую ошибку» между ними. При формировании саккад используется зрительная, соматосенсорная и слуховая информация. Информация о величине и направлении необходимого для фиксации изображения на сетчатке изменения положения глаза («двигательная ошибка») передается стволу мозга, в котором и вычисляются статические и динамические силы, необходимые для выполнения этого движения.
Саккады являются быстрыми скачкоподобными движениями глаза из одной точки фиксации к другой. Они чередуются с периодами фиксации, длящимися примерно 0,15—2,0 с. Амплитуда этих движений колеблется от нескольких угловых минут до нескольких градусов. Средняя угловая скорость при этом тем выше, чем больше амплитуда саккады, и доходит до 200—600° в секунду. Длительность саккады варьирует от 10 до 80 мс. Саккады с амплитудой 10—15° обычно сопровождаются дополнительным поворотом головы.
Типичное саккадическое движение глаза у человека равно десяти градусам и продолжается 45 мс. При произвольных саккадах длительность ее равняется 200 мс. Непроизвольные саккады более быстрые. Они бывают во сне, а также в течение быстрой стадии оптокинетического и вестибулярного нистагма, во время зрительной моторной реакции при неожиданном появлении цели по периферии поля зрения. Нередко в процессе саккадического движения глазное яблоко как бы «промахивается» относительно цели, т. е. цель не фиксируется в центральной ямке, а располагается дальше. Саккадное «промахивание» (дисметрическая саккада) встречается даже в норме. После дисметрической саккады всегда наступает корректирующее движение. В норме встречается два типа корректирующих движений: корректирующая саккада с определенным интервалом времени ожидания между наступившей и предыдущей саккадами и корректирующее «скользящее» движение, развивающееся без интервала при завершении оригинала саккады. «Скользящее» движение не требует нового управляющего импульса, идущего от мозга.
Саккадические движения нередко нарушаются при различных заболеваниях, сопровождающихся поражением головного мозга и глаза. Саккады отсутствуют при тяжелой прогрессивной надъядерной офтальмоплегии, при поражении лобно-среднемозговых (мезенцефалических) трактов. Нарушение временных характеристик саккад, их скорости и точности обнаруживается у больных с рассеянным склерозом в 57%. К нарушению саккадических движений может привести применение ряда фармакологических препаратов.
Анатомический субстрат саккад. Традиционно считают, что генерирует саккадические движения кора лобной области головного мозга после поступления в нее необходимой информации от органа зрения и наружных мышц глаза. Это добавочные поля первичной лобной зрительной коры, включая дополнительную зрительную кору и дорзальную прeлобную кору. Контролируют саккады также поле 7а затылочной коры, теменная кора, латеральное внутризатылочное поле, наружное коленчатое тело и базальные ганглии. Все вышеперечисленные области проецируются на верхние бугорки четверохолмия (рис. 4.4.12, 4.4.13).
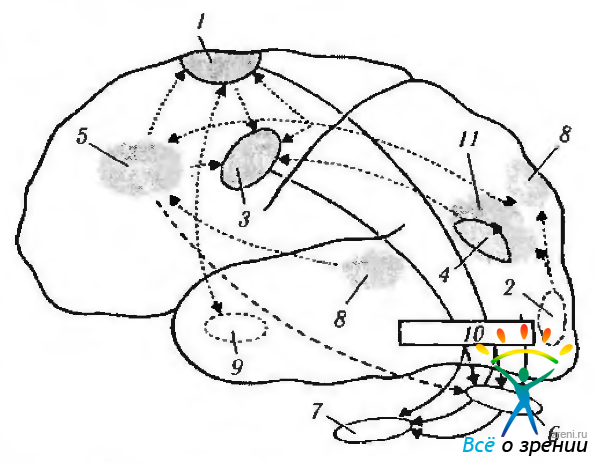
Рис. 4.4.12. Гипотетическая схема взаимодействия различных участков коры головного мозга в процессе саккадического движения глаз (по Pierrot—Deseillignv, Ann Neurol 1995, [37, 557—567]): 1—дополнительное глазодвигательное поле; 2 — затылочная кора: 3—лобное глазодвигательное поле: 4—теменное глазодвигательное поле; 5 — прелобная кора: 6 — верхние бугорки четверохолмия; 7—парамедианная ретикулярная формация варольева моста; 8—вестибулярная кора; 9—гиппокамп; 10 — зрительный бугор; 11 — теменная задняя кора
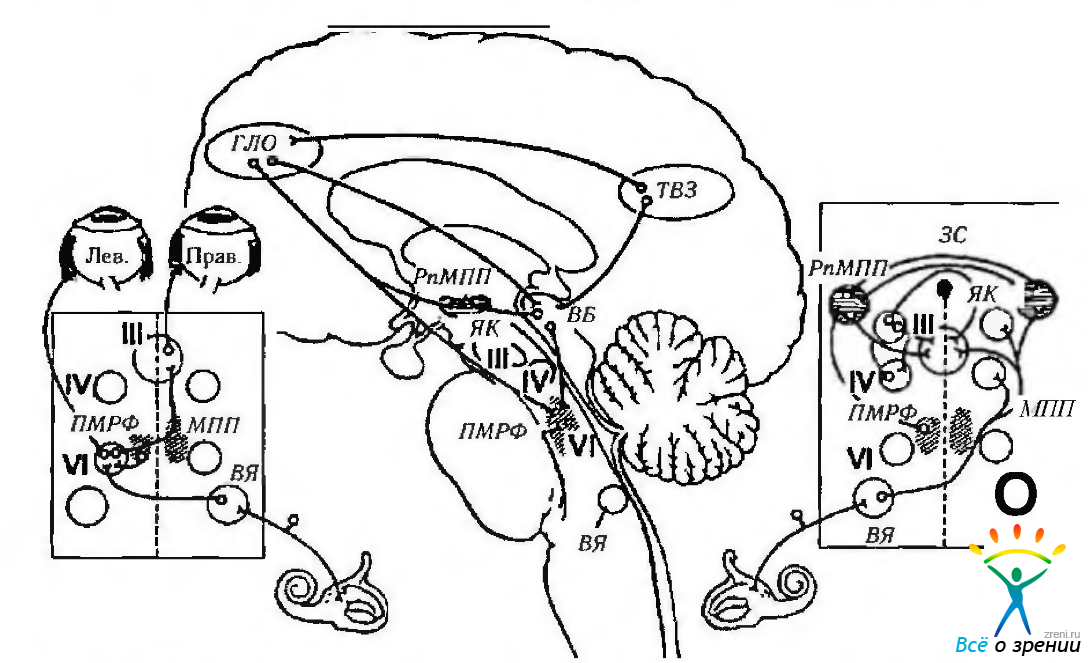
Рис. 4.4.13. Обобщенная схема нейрального контроля движений глазных яблок (по Miller, 1985): рисунок в центре показывает связь между глазной лобной областью (ГЛО) и теменно-височно-затылочной (ТВЗ) областью с верхними бугорками четверохолмия (ВБ), а также ростральным промежуточным ядром медиального продольного пучка (рпМПП) и парамедианной ретикулярной формацией варольева моста (ПРФА1). ГЛО и ВБ продуцируют саккады, в то время как ТВЗ является наиболее важным участком коры в контроле плавного слежения. Схематическое изображение, приведенное слева, иллюстрирует тракты в области ствола мозга, контролирующие горизонтальный взгляд. Аксоны клеточных тел, локализованных в ПРМФ, подходят к ипсилатеральному ядру отводящего нерва (VI), где они образуют синапс с мотонейронами отводящего нерва, аксоны которых, перекрещиваясь по средней линии, направляются в составе медиального продольного пучка (МПП) к часта ядра глазодвигательного нерва, иннервирующей внутреннюю прямую мышцу (ВП) противоположной стороны. Схематическое изображение справа иллюстрирует тракт ствола мозга вертикального взгляда. К наиболее важным структурам относятся рпМПП, ПРФМ, промежуточное ядро Кахала (ЯК) и задняя спайка (ЗС). Аксоны нейронов, расположенных в вестибулярном ядре (ВЯ), проходят мимо ядра отводящего нерва и посредством медиального продольного пучка подходят к ядрам глазодвигательного (III) и блокового (IV) нервов
Существуют данные относительно того, что перечисленные области мозга формируют сигнал для «пакетных нейронов». Участие и взаимодействие того или иного участка коры головного мозга зависит от типа саккадического движения, что проиллюстрировано рис. 4.4.12, 4.4.13.
Наиболее важной областью формирования саккад является кора лобной глазной области. В отличие от теменной коры и базальных ганглиев, кора лобной глазной области проецируется непосредственно на центры движения глаз, расположенные в мосту. После повреждения верхних бугорков четверохолмия только лобная кора способна генерировать саккады.
Как выше указано, нисходящий тракт от лобной коры сводится к прямой проекции на мост, а именно на его парамедианную ретикулярную формацию. Существует и непрямая проекция. При этом образуются синапсы в наружном коленчатом теле, и только затем проецируется на ретикулярную формацию (рис. 4.4.12, 4.4.13). Парамедианная ретикулярная формация играет решающую роль в генерации глазных движений. Располагается она между глазодвигательными ядрами по срединной линии ствола мозга. Предполагают, что парамедианная ретикулярная формация в физиологическом отношении состоит из двух частей. Большая часть расположена рострально. Функционирует она как «генератор импульсов», поскольку в эксперименте обнаружены в этой области нейроны, формирующие «пакет» потенциалов действия, начинающихся приблизительно за 8—10 мс перед движением глаза. Эти нейроны названы «пакетными» нейронами (рис. 4.4.14).
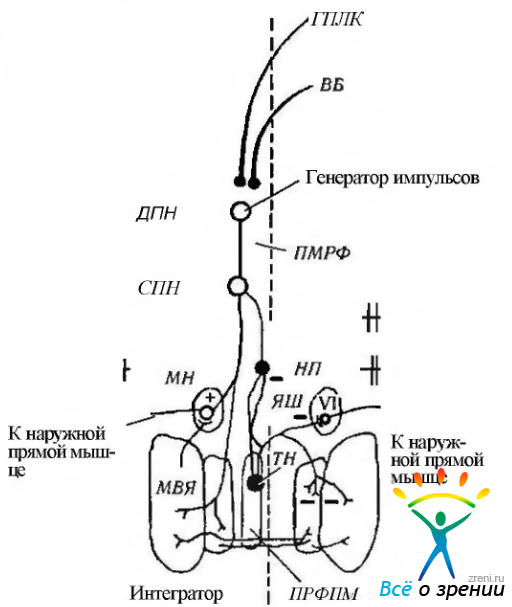
Рис. 4.4.14. Схематическое изображение генератора саккадических движений глаз, расположенного в стволе мозга (по Albert, 1995): изменение положения глаза (Е) и картина активности нейронов при саккадическом движении в левую сторону. Вход к стволу мозга происходит от верхних бугорков четверохолмия (ВБ) и глазного поля лобной коры (ГПЛК). Эти сигналы «перекрещиваются» в стволе мозга и возбуждают «пакетные» («физические») нейроны контрлатеральной парамедианной ретикулярной формации моста (ПМРФ). Длительно функционирующие «пакетные» нейроны (ДПН) разряжаются и возбуждают нейроны, функционирующие со средней длительностью (СПН). Именно эти нейроны кодируют так называемый «скоростной сигнал» саккад. Нейроны «паузы» (НП) расположены в вентрокаудальной части парамедианной ретикулярной формации в nucleus raphe interpositus (ядро шва — ЯШ). Эти клетки тормозят «пакетные» нейроны, функционирующие только с целью генерации саккад. Тормозные «пакетные» нейроны (771) лежат каудальней ядра отводящего нерва (VI) в дорзолатеральной части ретикулярной формации продолговатого мозга (ПРФПМ) и тормозят контрлате-рапьные мотонейроны. Сигнал о положении глаза генерируется «нейральным интегратором» в медиагьном вестибулярном ядре (МВЯ) и nucleus prepositus hypoglossy. Разряжение мотонейронов (МН) кодирует как скорость саккадического движения, так и его амплитуду («шаг»). Заштрихованные кружки — тормозные нейроны, а полые — возбуждающие
Выявлено также, что число импульсов этих нейронов линейно связано с амплитудой саккады в горизонтальной плоскости.
Каудальная часть парамедианной ретикулярной формации, лежащая вблизи ядер отводящего нерва, выполняет функцию «нейрального интегратора» и содержит «тонические» нейроны и «нейроны паузы». Располагаются клетки «нейрального интегратора» в nucleus prepositus hypoglossi и медиальном вестибулярном ядре. Нейроны этого ядра получают входы от премоторных «саккадных полей» ствола мозга и проецируются на ядра глазодвигательных нервов, мозжечок и вестибулярные ядра (рис. 4.4.14).
Подобное взаимодействие позволяет клеткам «нейрального интегратора» объединять получаемую сенсорную информацию и формировать сигнал, определяющий частоту и амплитуду саккады. При этом амплитуда саккад кодируется в премоторных центрах, обеспеченных топографическими сенсорными картами стриарной коры, лобной глазной области и верхних бугорков четверохолмия.
Особое место занимают верхние бугорки четверохолмия. Еще в конце XIX века выявлено, что стимуляция верхних бугорков приводит к возникновению саккадоподобных движений глаз. В 70-е годы XX столетия на основании экспериментальных электорофизиологических исследований установлены важные факты. Во-первых, верхние бугорки не генерируют саккады. Они определяют точность движения глаз по направлению к цели, а также определяют направление и амплитуду движения независимо от первоначального положения глаз в орбите, поскольку содержат «топографическую карту» движений глаза. Подтверждается это тем, что стимуляция соседних участков бугорков приводит к смещению взгляда к смежным точкам цели. Во-вторых, верхние бугорки четверохолмия участвуют и в координации движений глаза в ответ на звуковой раздражитель, соматосенсорную информацию, обеспечивая, таким образом, направление взгляда в сторону звукового сигнала или тактильного раздражителя.
Теперь мы несколько подробней остановимся на нейронном контроле саккад в горизонтальной плоскости. Формируется премоторная команда горизонтальной саккады, как и других движений в горизонтальной плоскости, в нейронах парамедианной ретикулярной формации моста, а затем проецируется на ипсилатеральное ядро отводящего нерва (рис. 4.4.13—4.4.15).
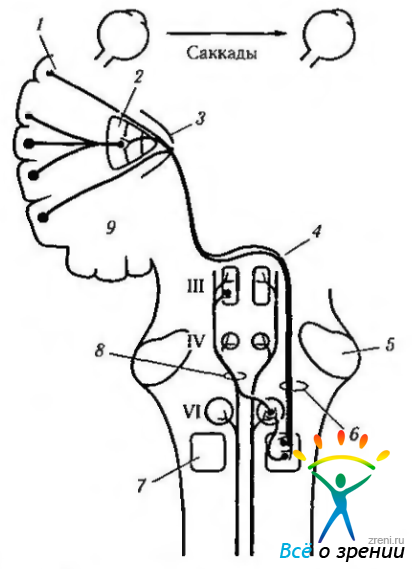
Рис. 4.4.15. Надъядерный контроль саккадических движений глаза, горизонтального взгляда в правую сторону (цель расположена в контрлатеральном поле) (по Reeh el al., 1985): 1 - лобное глазное двигательное поле (8); 2 — базальные ганглии; 3—внутренняя капсула; 4 — среднемозговой (мезанцефалический) перекрест; 5 — ножка мозжечка; 6—кортикобульбарный тракт (кортикотегментальный тракт); 7 — парамедианная ретикулярная формация варольева моста; 8—медиальный продольный пучок; 9—дополнительное двигательное поле (7). Основной путь показан красным цветом, а дополнительный путь, который может модифицировать горизонтальный взгляд, показан черным цветом
Ядро отводящего нерва лежит непосредственно по срединной линии в дорзальной части моста, сразу под дном четвертого желудочка. Оно содержит два типа нейронов. Большие мотонейроны ядра отдают аксоны, которые проходят вентрально через мост и выходят на внутренней поверхности ствола мозга, иннервируя наружную прямую мышцу глаза.
Ядро содержит также другие, маленькие, нейроны, чьи аксоны не покидают ствол мозга, а, перекрещиваясь, переходят на противоположную сторону, где входят в состав медиального продольного пучка и заканчиваются на нейронах ядер глазодвигательного нерва (рис. 4.4.14, 4.4.15). Именно благодаря этой связи и возбуждаются мотонейроны внутренней прямой мышцы глаза противоположной стороны, сокращение которой переводит взгляд.
«Тонические» нейроны ядра отводящего нерва возбуждаются перед началом саккадического движения и сохраняют такое состояние на протяжении всего движения.
Тормозные «тонические» нейроны располагаются несколько каудальней ядра отводящего нерва и обеспечивают взаимную иннервацию, посылая тормозные сигналы к контр латеральному отводящему ядру. Такая нейронная структура отвечает принципу антагонистической иннервации.
Вертикальные саккады также регулируются нейронами парамедианной ретикулярной формации моста. Но связь между этими нейронами и глазодвигательными ядрами не является прямой, а осуществляется через ретикулярную формацию среднего мозга. Кроме того, формируют вертикальный взгляд промежуточное (интерстициальное) ядро медиального продольного пучка, лежащее вблизи зрительного бугра с вентральной его стороны, а также ядро Кахала. Нейроны интерстициального ядра проецируются на нейроны глазодвигательного ядра, контролирующие верхнюю и нижнюю прямые мышцы и нижнюю косую мышцу. Более подробно нейронная связь между ядрами представлена на рис. 4.4.13 и 4.4.16.
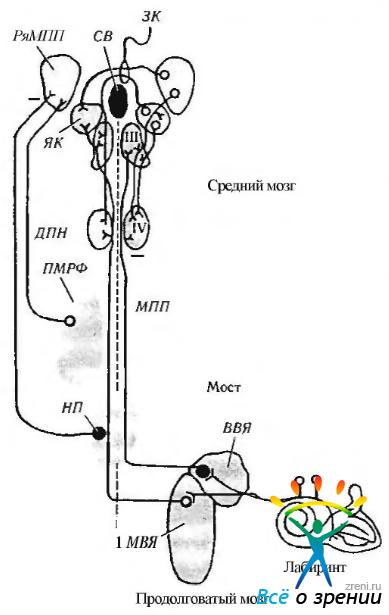
Рис. 4.4.16. Нейронный контроль содружественных вертикальных движений глаз: вестибулярные афференты от передних и задних полукружных каналов проецируются на верхнее (ВВЯ) и медиальное (МВЯ) вестибулярные ядра. Проекции от вестибулярных нейронов поднимаются в составе медиального продольного пучка (А1ПП) и соединительного плеча (не показано) к мотонейронам блокового нерва (IV), а также к ядру глазодвигательного нерва (III) и ядру Кахала (ЯК). Вестибулярные проекции, проходящие в ипсилатеральном МПП, ингибированы и исходят из верхнего вестибулярного ядра. Аксоны длительно функционирующих «пакетных» нейронов (ДПН) восходят из парамедианной ретикулярной формации моста (ПА1РФ) и направляются к ростральному промежуточному (интерстициальному) ядру медиального продольного пучка (ряАШП). Нейроны последнего кодируют вертикальные саккады и быструю стадию нистагма. Восходящие аксоны перекрещиваются несколько ростральней сильвиевого водопровода (СВ) в задней комиссуре (ЗК), в го время как нейроны, иннервирующие мотонейроны депрессора, проходят несколько латеральней и через ядро Кахала. Ядро Кахала и его реципроктные связи с вестибулярными ядрами формируют «нейральный интегратор» вертикальных движений. Команды плавного слежения проецируются на клочок (флоккулус) мозжечка от вестибулярного ядра. Заштрихованные кружки — тормозные нейроны, полые — возбуждающие; НП — нейроны паузы
Необходимо отметить, что нейроны парамедианной ретикулярной формации моста возбуждаются за несколько миллисекунд до начала вертикальной саккады. При этом они «кодируют» направление саккад. На эти клетки проецируются на верхние бугорки четверохолмия. кору лобной области и ядро клиновидного пучка (n. cuneatus), лежащее с латеральной стороны ретикулярной формации среднего мозга.
Нейроны ядра шва (n. raphes), расположенные около ядра отводящего нерва, тормозят «тонические» клетки между саккадами и в момент фиксации. Если эти нейроны, называемые нейронами «паузы», стимулировать во время саккады, движение глаза прерывается. Новое положение глаза в конце саккад поддерживается потоком импульсов, являющихся результатом функционирования «тонических» нейронов.
Плавное слежение. Плавные (следящие) движения глаз необходимы для длительной фиксации движущегося объекта и слежения за ним. При этом угловая скорость вращения глаз примерно соответствует скорости движения цели, если скорость последней не превышает 60—80° в секунду. Наиболее точно система плавного слежения работает тогда, когда скорость перемещения цели ниже 30° в секунду. После того как цель выбрана, система работает вне волевого контроля.
На плавные движения глаз накладываются коррекционные саккады. В этом случае изображение фиксируемой точки предмета остается в пределах 2° от центральной ямки. При скорости перемещения зрительного объекта более 80° в секунду следящие движения глаз происходят значительно медленнее, чем движение предмета. По этой причине изображение передвигается по сетчатке. Если скорость движения изображения не превышает 180° в секунду, то смещение изображения по сетчатке компенсируется дополнительными саккадическими движениями большей амплитуды. Координация таких следящих движений глаз обычно осуществляете я за счет последующих движений головы. Координация движений головы и глаз обычно осуществляется в центрах управления взгляда, расположенных в стволе мозга. Информация относительно положения головы и положения глаз поступает в эти центры не только от вестибулярных рецепторов внутреннего уха, но и от механорецепторов суставов шеи.
Вышеперечисленные сложные функции слежения, сочетающиеся с саккадами и находящиеся под контролем вестибулярной системы и мозжечка, обеспечиваются сложной системой нервного контроля, о чем подробно будет изложено ниже.
Продолжение в следующей статье: Движения глаз ? Часть 2
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0