Об особенностях различных цветореагирующих аппаратов зрительного анализатора. Часть 1.


Содержание:
Описание
↑ Орган зрения в связи с организмом как целым. Метод непрямых раздражений
«Анализатор,— читаем мы у Павлова (1949),— есть сложный нервный механизм, начинающийся наружным воспринимающим аппаратом и кончающийся в мозгу то в низшем отделе его, то в высшем, в последнем случае бесконечно более сложным образом. Основным фактом физиологии анализаторов является то, что каждый периферический аппарат есть специальный трансформатор данной внешней энергии в нервный процесс. А затем идет длинный ряд или далеко, или совершенно не решенных вопросов». Вместе с тем Павлов всегда подчеркивал, что «деятельность нервной системы направляется, с одной стороны, на объединение, интеграцию работы всех частей организма, с другой — на связь организма с окружающей средой». Естественно поэтому было полагать, что и деятельность зрительного анализатора, осуществляя связь организма с окружающей средой, тесным образом связана со всем организмом как целым направлена на объединение, интеграцию всех его частей.
Имея в виду приблизиться к уяснению всех закономерностей, которые управляют деятельностью нашего органа зрения в связи с состоянием организма как целого, мы и наши сотрудники уже около двадцати лет назад при изучении функций зрения стали пользоваться методом непрямых раздражений. Применяя различные неадекватные для глаза стимулы и наблюдая за темп изменениями, которые эти стимулы вызывают в состоянии и функциях зрительного анализатора, мы получали возможность вскрывать зависимости зрения от изменений, происходящих в других участках нервной системы. Был проведен ряд специальных исследований цветового зрения методом непрямых раздражении. Полученные данные, хорошо укладываясь в схему трехкомпонентной теории цветового зрения, подтверждают ее и позволяют, как нам кажется, ближе конкретизировать природу отдельных цветоощущающих аппаратов, а также существующие между ними отношения.
Первые описания изменений цветового зрения под влиянием непрямых раздражителей, затрагивающих другие органы чувств, мы находим у Урбанчича (Urbantschisch, 1888). Он предлагал испытуемым смотреть на небольшие цветные пятна с такого расстояния, что цвета их почти или совсем не различались. При приближении к ушам испытуемых звучащих камертонов ранее не замечавшиеся цветные оттенки пятен становились зачастую хорошо видимыми. В иных случаях наблюдалось понижение цветовой чувствительности под влиянием звуков. Какой-либо определенной закономерности в изменениях цветового зрения при воздействии слуховых раздражений им, однако, установлено не было. Этим автором опыты ставились с влиянием на цветовое зрение не только слуховых раздражителей. Так, испытывалось действие раздражителей вкусовых, обонятельных и осязательно-температурных. Под действием всех этих агентов обычно наступали те или иные изменения в цветовых ощущениях. Однако опыты не вскрыли какой-либо определенной закономерности в реакциях глаза на применявшиеся непрямые раздражители. Недостатком методики здесь было то, что при определении порога видности в первые моменты действия непрямого раздражителя не применялся искусственный зрачок. От непрямых раздражителей зрачковое отверстие у испытуемых, несомненно, менялось, чем вызывались неучитываемые им изменения в условиях оценки глазом раздражителей, близких к пороговым.
↑ Влияние слуховых раздражений на цветовое зрение
Начиная с 1936 г. влияние слуховых раздражений на цветовое зрение исследовалось нами и нашими сотрудниками. Испытывались цветовая чувствительность центрального зрения, критическая частота мелькании и границы цветных полей зрения. Во всех случаях применялся искусственный зрачок.
Абсолютная цветовая чувствительность определялась установлением «порога исчезновения хроматичности», т. е. установлением той пороговой яркости цветового раздражителя, при которой испытуемый субъект уже переставал замечать в раздражителе какую-либо хроматичность. Во всех ниже описываемых результатах опытов величина этого порога бралась как величина относительная, даваемая величиной пропускания (в %) того места поглощающего свет фотоклина, которое оказывалось перед источником света.
Методика измерения критической частоты мельканий состояла, как обычно, в определении того минимального числа периодов светового раздражения в секунду, при котором прерывистый свет (при равенстве периодов света и темноты) уже переставал казаться мигающим.
Границы полей зрения для цветных раздражителей определялись на периметре в условиях световой адаптации глаза. Определения же абсолютной цветовой чувствительности глаза, как и определения критической частоты мельканий, производились в ходе 1,5—2-часовой темновой адаптации обоих глаз испытуемого.
В качестве прямых раздражителей применялись различные монохроматические лучи спектра. Непрямой слуховой раздражитель, создававшийся ламповым генератором, имел частоту в отдельных сериях опытов около 800 и 2000 герцев (колебаний в секунду) и постоянный уровень громкости. Звук подводился к обоим ушам испытуемого приблизительно на 40-й минуте темновой адаптации и длился 10 мин.
Опыты давали возможность определять уровень цветовой чувствительности глаза в отсутствии слухового раздражения (Е2) и во время звука (Е2). Оказалось, что величина E2/E1 изменяется в зависимости от длины волны раздражителя вполне закономерно (рис. 44).

Рис. 44. Изменение колбочковой чувствительности в зависимости от звукового раздражения (по Кравкову)
Кружки на рис. 44 обозначают найденные в опытах значения E2/E1 (среднее из данных нескольких испытуемых).
Чувствительность зрительного анализатора к зелено-голубым лучам под влиянием звуков заметно повышается, к лучам оранжевато-красным снижается. Обращает на себя внимание то, что для средней части спектра (для лучей желтых с длиной волны около 570 m?) цветовая чувствительность при воздействии звуков не меняется. Опыты с длинноволновыми (690 m?) и коротковолновыми (430 m?) лучами показали, что на чувствительность к этим, концевым лучам спектра слуховые раздражения не влияют.
На цветовую чувствительность влияют не только звуки, но и шумы и притом влияют тоже противоположным образом по отношению к зеленому и красно-оранжевому цветам.
Если под влиянием звукового раздражения происходит известная «перестройка» нашего цветоощущающего аппарата, то можно было бы ожидать, что перестройка скажется на таком чувствительном показателе состояния нашего цветового зрения, каким является уравнение Релея, устанавливаемое на аномалоcкопе Нагеля. Специальные опыты (Кравков, 1937) действительно показали, что в условиях звукового раздражения большой громкости испытуемые берут в смеси красного с зеленым, подравниваемой желтому, меньше зеленых лучей, чем в условиях тишины. Это подтверждает, что чувствительность к зеленым лучам при звуке оказывается повышенной. В табл. 7
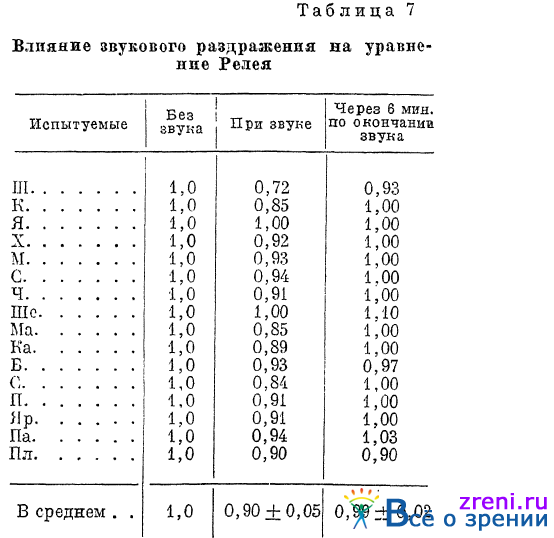
приведены соответствующие данные опытов, проведенных на 16 лицах. Цифры в таблице обозначают величины отношений количества зеленых лучей к красным, нужные для получения равенства цвета смеси с монохроматическим желтым цветом. Отношение, устанавливавшееся в опытах без звука, для всех испытуемых принято за единицу.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Совершенно такие же изменения цветовой чувствительности наблюдались Яковлевым (1940) при изучении границ цветовых полей зрения при звуках и шумах. Границы полей зрения для синего и зеленого цветов от слуховых раздражений расширяются, для оранжевого суживаются, для крайнего красного остаются без перемен (рис. 45).
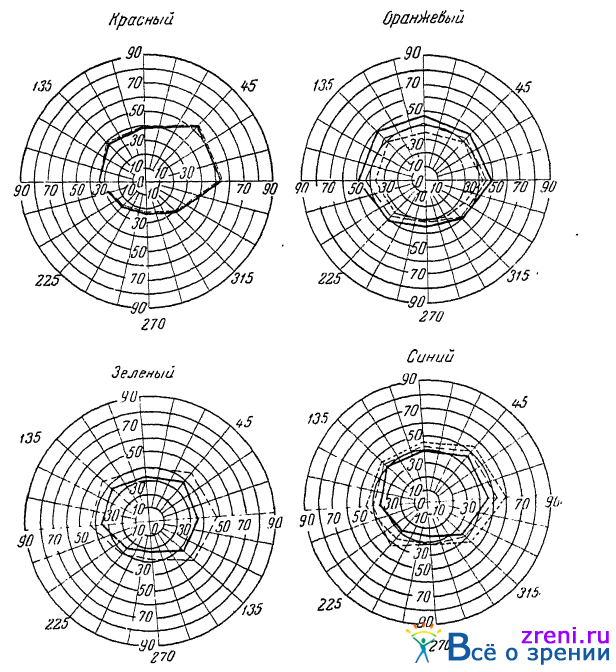
Рис. 45. Изменение границ цветовых шлей зрения в зависимости от слухового раздражения (по Яковлеву)
Сплошная линия —без слухового раздражения; прерывистая — при звуке; пунктирная —при сильном шуме. Цифрами указаны угловые градусы
Сплошная линия —без слухового раздражения; прерывистая — при звуке; пунктирная —при сильном шуме. Цифрами указаны угловые градусы
Далее проводились опыты с целью выяснить, в какой степени изменяемость цветовой чувствительности под влиянием звуков зависит от уровня громкости этих последних (Кравков, 1939). Определялась цветовая чувствительность к монохроматическим оранжевым (590 m?) и зеленым лучам (530 m?). Слуховым раздражителем служил звук от лампового генератора с частотой 775 герцев. Уровень громкости изменялся в пределах 25—95 децибел. Оказалось, что, несмотря да столь значительную разницу в громкости, направление изменений цветовой чувствительности под влиянием звуков оставалось тем же, т. е. повышение чувствительности к зеленому и понижение к оранжевому. В зависимости от громкости лишь менялась степень изменения цветовой чувствительности. На рис. 46

Рис. 46. Влияние громкости звукового раздражителя на колбочковую чувствительность глаза (по Кравкову)
приведены результаты этих опытов По абсциссе отложены громкости звука в децибелах, по ординате значения E2/E1. Крестики — результат для ? = 530 m?, кружки для ? = 590 m?.
В опытах Добряковой (1944) определялось, как изменяется под влиянием звукового раздражения критическая частота слития мельканий различных монохроматических лучей. Опыты проводились в условиях темнотой адаптации. Испытуемый смотрел монокулярно в окуляр спектроскопа через постоянный искусственный зрачок. Размер раздражающего поля равнялся 1,3°. Яркость же его соответствовала критической частоте, равной 16—20 пер/сек в первые минуты темновой адаптации. Звук, громкостью около 70 децибел и с частотою около 800 герцев, подводился к обоим ушам испытуемого и длился 10 мин. Средние данные опытов па трех лицах приведены на рис. 47.
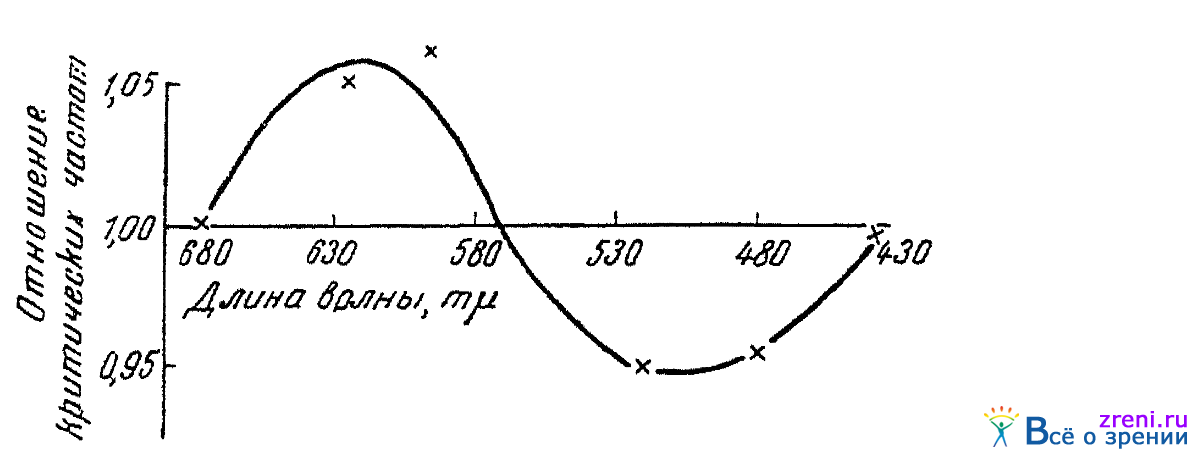
Рис. 47. Влияние звукового раздражения на критическую частоту слияния мельканий спектральных лучей (по Добряковой)
По вертикали отложены отношения критической частоты слияния мельканий, найденной при слуховом раздражении, к критической частоте, найденной без слухового раздражителя. По горизонтали показаны длины волн световых раздражителей.
Специальные опыты Кравкова (1939) установили, что изменения критической частоты мельканий под влиянием непрямых раздражителей — ее повышения или снижения — в условиях данных опытов обратны изменениям абсолютной чувствительности.
Таким образом, наблюдавшиеся понижение критической частоты мельканий при воздействии звука для зелено-синих лучей и ее повышение для лучей оранжево-красных означают, что чувствительность к зелено-синим лучам от звука повышалась, а к оранжево-красным снижалась.
↑ Влияние обонятельных раздражении на цветовое зрение
Изучалось, далее, и то, как влияют на колбочковую чувствительность различные обонятельные раздражители. Было исследовано действие средней силы запахов бергамотового масла, гераниола и индола. В качестве основного метода мы пользовались здесь определением критической частоты мельканий. Приблизительно на 40-й минуте темповой адаптации мы подносили к носу испытуемого флакон с пахучим веществом. Обонятельный раздражитель оставался перед испытуемым 10—12 мин., в течение которых производилось несколько определений критической частоты мельканий. По прекращении непрямого раздражителя критическая частота мельканий определялась еще в течение 20—30 мин. Так мы находили критическую частоту мельканий при действии непрямого обонятельного раздражения, а равно и ту величину ее, которая соответтвует условиям без обонятельного раздражения. Прерывающимся световым раздражителем были монохроматические лучи различных длин волн. Их яркость соответствовала в начале темновой адаптации критической частоте слияния мельканий, равной 12—20 пер/сек.
Опыты показали, что критическая частота мельканий при действии запахов бергамотового масла и гераниола изменяется для различных лучей спектра вполне закономерно. Эту закономерность можно хорошо видеть на рис. 48,
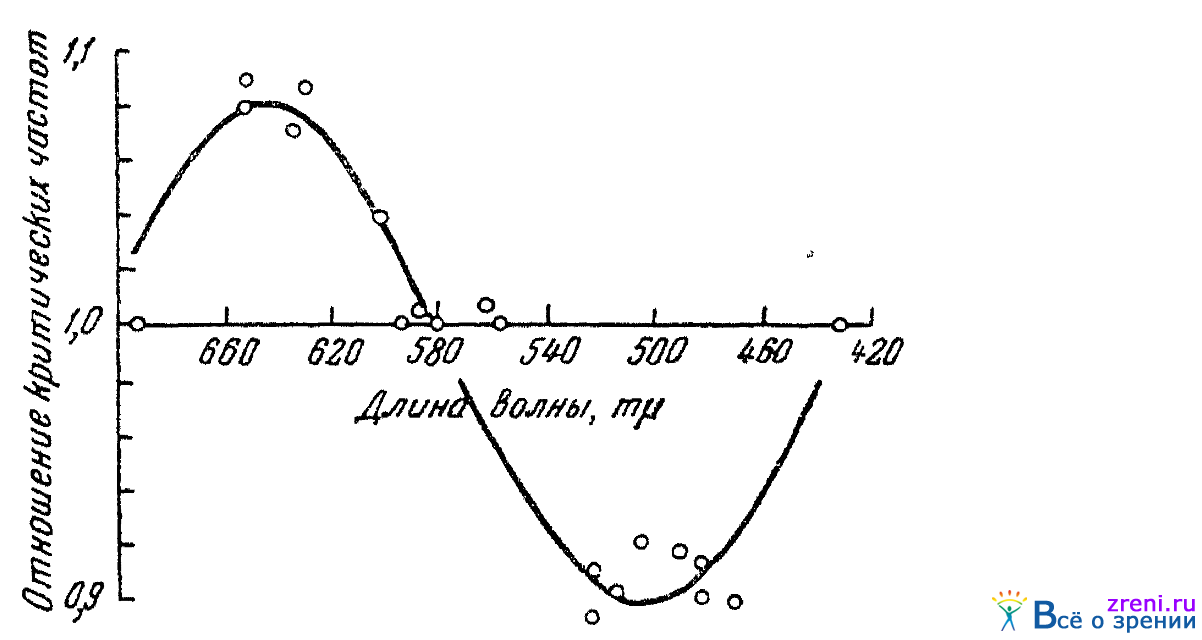
Рис. 48. Влияние обонятельного раздражения (запаха бергамотового масла) на критическую частоту слияния мельканий спектральных лучей (по Кравкову)
где приведены данные опытов с бергамотовым маслом с двумя испытуемыми. По ординате отложена относительная величина критической частоты слияния мельканий; за единицу принято значение, найденное без обонятельного раздражителя. Мы вновь видим, что спектр в отношении реакций глаза на непрямые раздражители распадается на две половины — зелено-синюю и оранжево-красную. То же самое наблюдалось и при применении запаха камфоры: снижение критической частоты для лучей зеленых и повышение ее для лучей оранжевато-красных.
Иной результат, а именно отсутствие каких-либо заметных изменений, был получен при применении запаха индола. По интенсивности запах этот был не слабее прочих и вызывал обычно у испытуемых чувство неприятного.
Под влиянием обонятельных раздражителей могут изменяться и границы цветовых полей зрения. Это было показано Селецкой (1949), применявшей запахи розмарина и индола. Поля зрения определялись для маленьких квадратиков красного и голубовато-зеленого цвета, по своему цветовому тону соответствовавших 640 и 496 m? Границы полей зрения отмечались по четырем меридианам. Опыты были проведены на 13 лицах и определенно показали, что под влиянием розмарина границы полей зрения для зеленого расширяются, а для красного суживаются, под влиянием же запаха индола наблюдалось обратное, т. е. сужение границ поля зрения для зеленого и расширение границ для красного. Последнее, впрочем, имело место лишь при применении испытательных объектов малого размера. Типичный результат опытов Селецкой иллюстрирован на рис. 49,
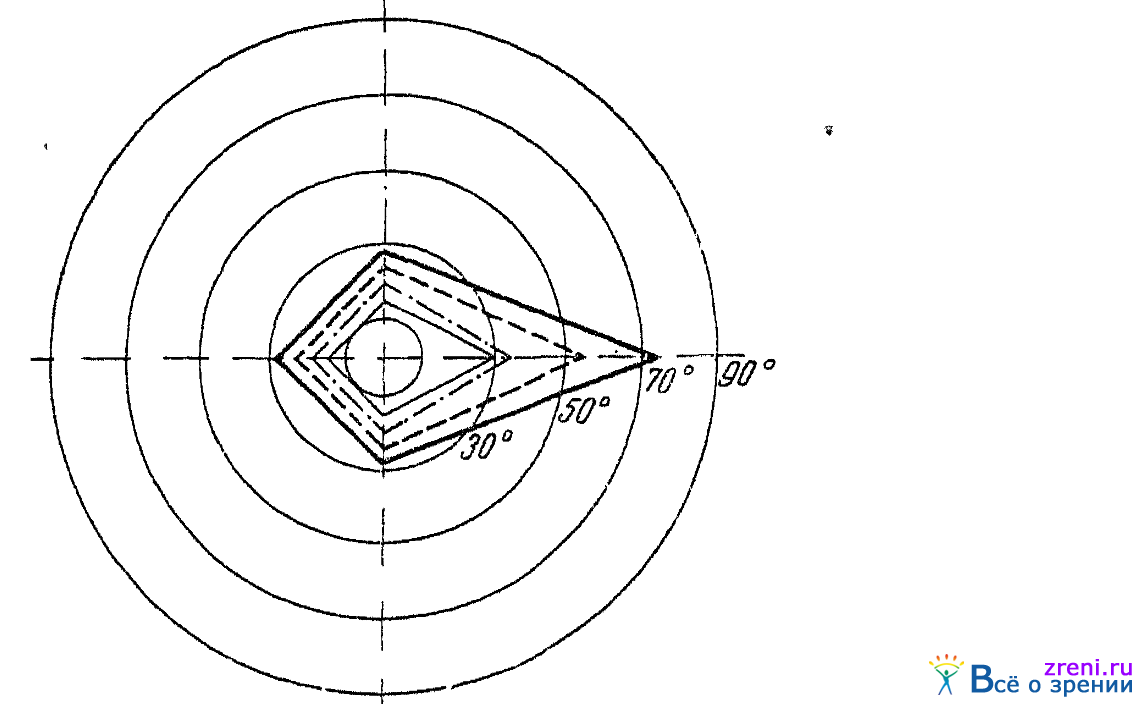
Рис. 49. Влияние обонятельного раздражения (запахом розмарина) на границы цветовых полей зрения правого глаза (по Селецкой)
— зеленый (без обонятельного раздражения)
------ зеленый (при обонятельном раздражении)
— красный (без обонятельного раздражения)
— — — красный (при обонятельном раздражении)
— зеленый (без обонятельного раздражения)
------ зеленый (при обонятельном раздражении)
— красный (без обонятельного раздражения)
— — — красный (при обонятельном раздражении)
где показаны границы цветовых полей зрения для красного и зеленого цветов, имевшиеся до обонятельного раздражения розмарином и при таком раздражении.
↑ Влияние температурных, вкусовых и постуральных раздражений на цветовое зрение
Имеются работы и по выяснению того, как сказываются на цветовой чувствительности глаза непрямые раздражители, действующие на температурный и вкусовой рецепторы (Добрякова, 1941). Температурным раздражителем в опытах Добряковой было тепло (40—45°) от электрической печи, действовавшее на кисти рук испытуемых. Вкусовым раздражителем служил сахар. Кусок сахара испытуемый держал в течение 10 мин. во рту, после чего рот ополаскивался водой комнатной, температуры.
Результаты этих опытов дали ту же картину, что и при испытании в качестве непрямых раздражителей действия звука и некоторых запахов, т. е. повышение цветовой чувствительности к области зелено-синего и снижение ее к лучам оранжево-красным. В области желтого (около 570 m?) опять-таки имелась нейтральная зона, где никаких изменений цветовой чувствительности не было обнаружено. К крайним лучам спектра чувствительность глаза также остается без перемен.
Обратное изменение цветовой чувствительности к зеленому и красному цветам в ответ на один и тот же непрямой раздражитель было описано и Шварц (1947). В Институте психологии ею исследовалось влияние постуральных раздражений (прямое положение головы и запрокинутое). При запрокинутой голове цветовая чувствительность к зеленому цвету (520 m?) заметно снижается, доходя через 90 мин. до 25 % исходной величины; к оранжево-красному цвету (610 m?) цветовая чувствительность, напротив, несколько повышается (рис. 50).

Рис. 50. Влияние запрокинутого положения головы на колбочковую чувствительность (по Шварц)
↑ Вегетативная противоположность двух групп непрямых раздражений
Описанные выше опыты, таким образом, согласно показывали, что одни непрямые раздражители повышают колбочковую чувствительность к зелено-синим лучам и понижают к лучам оранжево-красным, другие, наоборот, понижают ее к лучам зелено-синим и повышают к лучам оранжево-красным. В качестве непрямых раздражителей первого рода следует назвать слуховые раздражения, запахи бергамотового масла, гераниола, камфоры и розмарина, вкусовое раздражение сладким, легкое тепловое раздражение. В качестве же непрямых раздражителей второго рода проявили свое действие запах индола и положение запрокинутой головы.
Естественно поставить вопрос, что есть общего для группы первых и для группы вторых раздражителей. Нам представлялось вероятным допущение, что общим является характерное действие той и другой группы непрямых раздражителей на вегетативную нервную систему. Первая группа непрямых раздражителей (слуховые раздражения, запахи гераниола и др.) возбуждают преимущественно симпатический отдел вегетативной нервной системы. Вторая группа непрямых раздражителей (запах индола, положение запрокинутой головы) возбуждает преимущественно парасимпатический отдел вегетативной нервной системы.
Первая группа раздражителей ускоряющим образом действует на пульс. Слуховые раздражители сосудосуживающим образом, т. е. симпатикотропно, влияют на сосуды сетчатки. Последнее было продемонстрировано в свое время офтальмоскопически московским отиатром Штейном (Stein, 1910). Парасимпатикомиметическое действие непрямых раздражителей второй группы проявляется в обратном (замедляющем) действии их на пульс, а также и в таких симптомах, как тошнотные движения. Вегетативную противоположность запахов бергамотового масла и гераниола, с одной стороны, и индола, с другой — отмечает между прочим Мартине (Martinet, 1925).
↑ Изменения цветового зрения под влиянием вегетотропных медикаментов
Предположение о зависимости цветовой чувствительности глаза от изменений в состоянии вегетативной нервной системы нашло подтверждение в дальнейших опытах по изучению действия на колбочковую чувствительность нашего зрительного анализатора разного рода вегетотропных веществ.
Нами (Кравков, 1941, 1949) испытывалось изменение цветовой чувствительности глаза к различным монохроматическим лучам спектра под влиянием закапывания (инстилляции) в конъюнктивальный мешок адреналина. Применялся раствор 1 : 1000 и инстилляции производились обычно около 60-й минуты темповой адаптации, когда чувствительность устанавливалась уже на достаточно постоянном уровне. Прослеживался дальнейший ход адаптационной кривой вплоть до 100—120-й минуты.
Уровень чувствительности, обнаруживаемый на 100-й минуте, и учитывался нами как показатель действия адреналина. Относительная величина этого уровня показана в качестве ординаты на рис. 51.
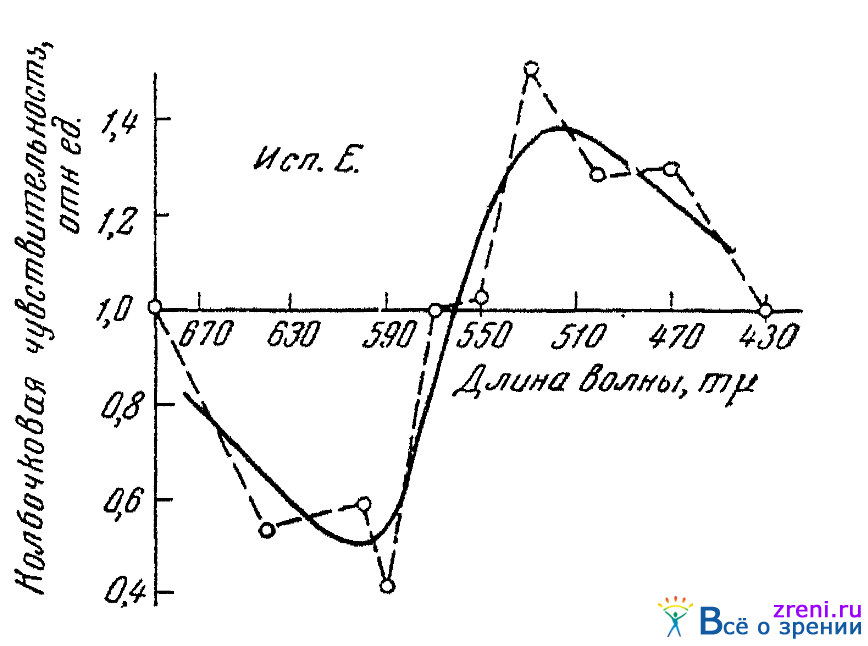
Рис. 51. Влияние инстилляции адреналина на колбочковую чувствительность (по Кравкову)
Рис. 52

Рис. 52. Влияние инстилляции адреналина (Л) на колбочковую чувствительность в ходе темновой адаптации (по Кравкову)
показывает ход адаптационных кривых, иллюстрируя картину отдельных опытов с лучами с длинами волн 530 и 600 m?. По абсциссе отложено время, по ординате — чувствительность. Время инстилляции адреналина отмечено стрелкой с буквою А. В описываемых опытах испытуемые должны были отмечать порог исчезновения хроматичности поля, видимого ими центральной ямкой сетчатки (размер поля 1,3°).
Адреналин является, как известно, возбудителем по преимуществу симпатического отдела вегетативной нервной системы. К подобным же симпатикотропным веществам относится и эфедрин. Опыты с эфедрином проводились Селецкой (1948). Эфедрин (0,025 г) принимался внутрь в виде таблеток. Для глаз, адаптированных к небольшой яркости, определялся порог исчезновения видности маленького цветного поля, видимого фовеально. Опыты были проведены с раздражителями красного и зеленого цвета. В приборе применялись для этого специальные светофильтры красного и зеленого цвета. В согласии с тем, что сообщено нами выше о действии адреналина, опыты Селецкой показали, что чувствительность к зеленому от эфедрина повышалась (в 26 случаях из 29), чувствительность же к красному понижалась (в 26 случаях из 32).
Другое симпатикотропное вещество—кордиамин, испытывавшееся в опытах Шварц (1948), показало совершенно подобный же эффект. К зелено-синим лучам спектра колбочковая чувствительность после приема кордиамина повышалась, к оранжево-красным понижалась. Кордиамин принимался внутрь (20 капель 25%-ного водного раствора).
С другой стороны, применение фармакологических средств, по преимуществу парасимпатикотропной природы, установило обратную картину изменения цветовой чувствительности, т. е. как раз ту, которая описана выше как результат действия запаха индола и запрокинутой головы.
Наши опыты (Кравков, 1941) установили, что закапывание в конъюнктивальный мешок 2—3 капель 1%-ного раствора пилокарпина заметно повышает критическую частоту слияния мельканий для лучей зеленых (520 m?) и заметно снижает ее для лучей оранжевых (590 m?). Применительно к чувствительности, как мы уже говорили выше, это обозначает повышение чувствительности к оранжевому и снижение ее к зеленому цвету. Рис. 53
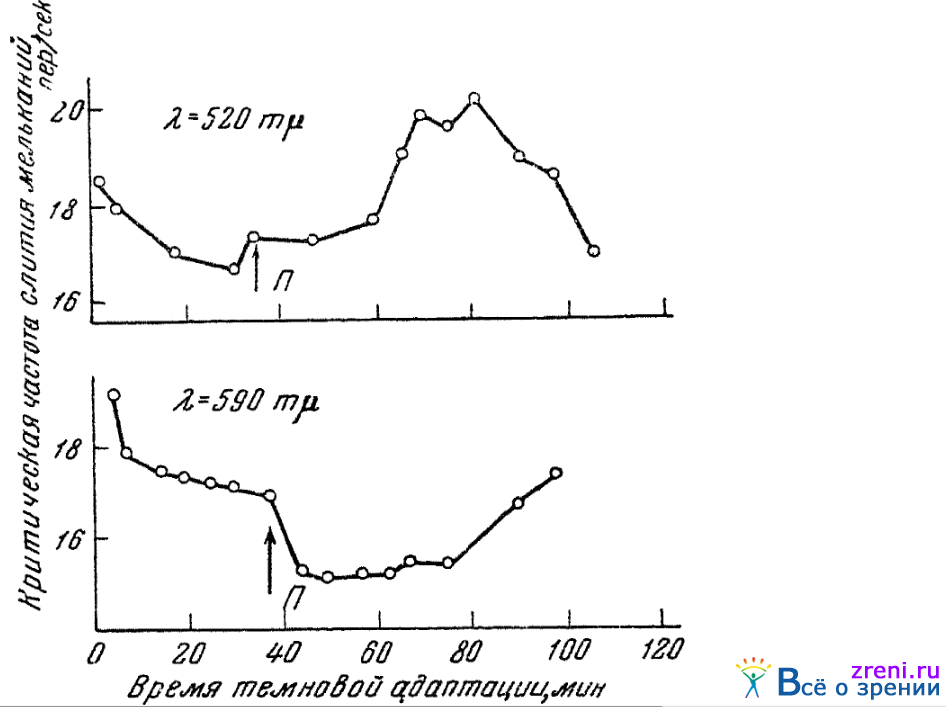
Рис. 53. Влияние инстилляции пилокарпина (П) на критическую частоту слияния спектральных лучей (? = 520 и 590 m?) (по Кравкову)
иллюстрирует сказанное, воспроизводя данные отдельных опытов.
Селецкая (1948) применяла в своих опытах парасимпатикотропное вещество — карбохолин. Испытуемые выпивали 7 см3 раствора карбохолина (при разведении 1 : 10 000). Измерялась колбочковая чувствительность в условиях световой адаптации. В большинстве опытов после приема карбохолина наблюдалось ухудшение чувствительности к зеленому и улучшение чувствительности к красному.
Упомянем здесь и об опытах Галочкиной (1948) с китайским лимонником. Китайский лимонник — вещество по своему действию на сердечно-сосудистую систему парасимпатикотропное. Он вызывает урежение пульса и падение кровяного давления. На цветовой чувствительности китайский лимонник сказывается подобно парасимпатикомиметическим веществам, повышая чувствительность к красному и снижая ее к зеленому (рис. 54).
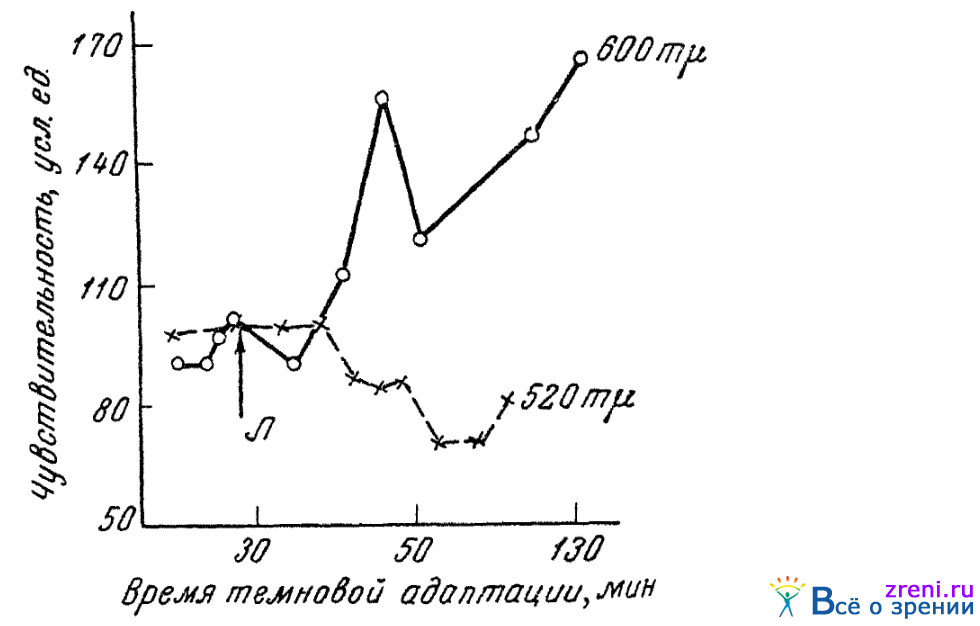
Рис. 54. Влияние китайского лимонника (Л) на колбочковую чувствительность (по Галочкиной)
Итак, опыты с медикаментозными воздействиями вместе с ранее описанными результатами опытов с непрямыми раздражителями говорят за противоположную вегетативную природу цветоощущающих аппаратов, возбуждаемых коротко- и длинноволновыми лучами спектра. Если зелено- и синеощущающие аппараты глаза повышают возбудимость при повышении тонуса симпатического отдела вегетативной нервной системы, то аппарат красноощущающий в этом случае делается менее чувствительным. Повышение же тонуса парасимпатического отдела вегетативной нервной системы в обратном направлении изменяет чувствительность цветоощущающих аппаратов нашего зрительного анализатора: красноощущающий аппарат оказывается более возбудимым, а зелено- и синеощущающие аппараты обнаруживают пониженную чувствительность.
Вегетативные же изменения, несомненно, зависят и от состояния коры головного мозга. Достаточно здесь вспомнить побледнение или покраснение лица, возникающие под влиянием тех или иных мыслей. Можно предполагать поэтому, что вегетативные сдвиги являются одним из путей влияния коры головного мозга на цветовое зрение.
↑ Изменения цветового зрения под влиянием ан- и катэлектротонических раздражений
Описанные новые факты, устанавливающие связи нашего цветового зрения с состоянием того или иного отдела вегетативной нервной системы, нашли себе дальнейшее углубленное толкование в ряде последующих работ, выполненных в наших лабораториях.
Были поставлены опыты (Кравков и Галочкина, 1947) по выяснению того, как сказывается на колбочковой чувствительности постоянный ток, пропускаемый через глазное яблоко.
Ток брался через кенотронный выпрямитель с фильтром и потенциометром. Один из электродов в форме овального кольца прикладывался к глазу; индифферентный электрод брался испытуемым в руку противоположной стороны. В некоторых сериях опытов индифферентный электрод ставился испытуемому сзади на шею. Исследуемый глаз мог быть открытым и во время электризации. Сила тока обычно была всего около 0,02 mА, иногда же около 0,2 и 0,5 mА. Продолжительность действия тока 3—15 мин. Опыты протекали в условиях темновой адаптации.
Ток включался около 30-й минуты адаптации. Исследуемым глазом фовеально определялись пороги видности различных монохроматических лучей по методу гашения (отмечалось полное исчезновение света). Контрольные опыты показали, что при определении порога на исчезновение хроматичности результаты оказываются теми же самыми.
Данные многих подопытных лиц позволили установить здесь вполне определенную закономерность. Под влиянием тока цветовая чувствительность обнаруживает заметные сдвиги, причем направление сдвигов чувствительности зависит от направления тока (от того, какой полюс находится на глазном яблоке). По прекращении тока обычно наблюдались изменения чувствительности в обратном направлении по сравнению с тем, что было во время прохождения тока через глаз. На рис. 55
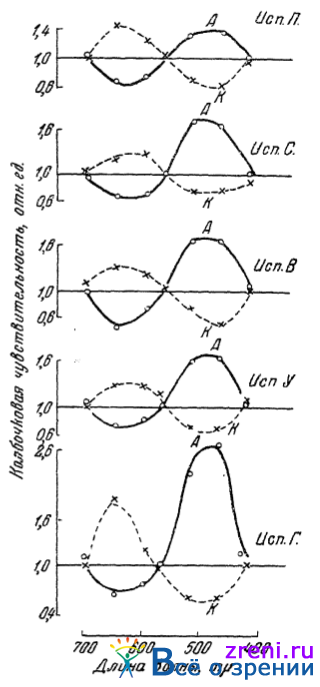
Рис. 55. Влияние ан- и катэлектротона на колбочковую чувствительность глаза (по Кравкову и Галочкиной)
приведены типичные результаты опытов с пятью лицами. По ординате отложена относительная величина цветовой чувствительности на 3-й минуте действия тока на глаз. За единицу принят уровень чувствительности перед включением тока.
Обнаруживающаяся закономерность выражена весьма явно. Именно, под влиянием анэлектротона (анод на глазе) колбочковая чувствительность темноадаптированного глаза к зеленосиним лучам повышается, к оранжево-красным падает; под влиянием катэлектротона (катод на глазе) чувствительность к зелено-синим падает, к оранжево-красным повышается. К желтым (около 570 m?) и концевым лучам спектра (около 700 и 425 m?) чувствительность под влиянием тока обоих направлений совсем не изменяется.
↑ Роль изменения концентраций ионов в глазе. Опыты с ионофорезом калия кальция, натрия и магния
Попытки объяснить найденные закономерности действия тока на глаз привели к мысли, что здесь имеет значение происходящее вблизи полюсов изменение концентраций ионов в средах глаза. Естественно было подумать прежде всего об изменениях в относительной концентрации ионов калия и кальция, поскольку известно важное значение этих катионов для возбудимости живых тканей. Одновалентные ионы калия в поле постоянного тока скорее передвигаются к катоду, чем двухвалентные ионы кальция. Поэтому ионный показатель К/Са у катода возрастает, а у анода уменьшается.
Чтобы проверить это предположение, Кравковым и Галочкиной (1947) были поставлены опыты с введением в глазное яблоко ионов калия и кальция путем ионофореза. Подушечки, смоченные растворами хлористого кальция или иодистого калия, помещались на анод, накладываемый на веки закрытого глаза. Сила тока равнялась 0,5 mА. Ионофорез длился 20 мин. Опыты протекали в условиях темновой адаптации. Подопытные определяли испытываемым глазом порог исчезновения, фовеально фиксируемого, монохроматического света. При измерении порога во время ионофореза электрод с подушечкой сдвигался на короткое время с век к виску.
Результаты опытов подтвердили правильность предположения о роли ионов калия и кальция. При ионофорезе калия или кальция в цветовой чувствительности глаза наступали изменения противоположного характера, несмотря на то, что в обоих случаях активным электродом на глазном яблоке был, конечно, анод. При введении в темноадаптированный глаз ионов кальция чувствительность к зеленому повышается, к оранжевокрасному падает. По прекращении ионофореза наступают сдвиги чувствительности в обратном направлении. При введении в глаз ионов калия происходят изменения прямо противоположные: чувствительность к зеленому падает, к оранжевокрасному возрастает. На рис. 56
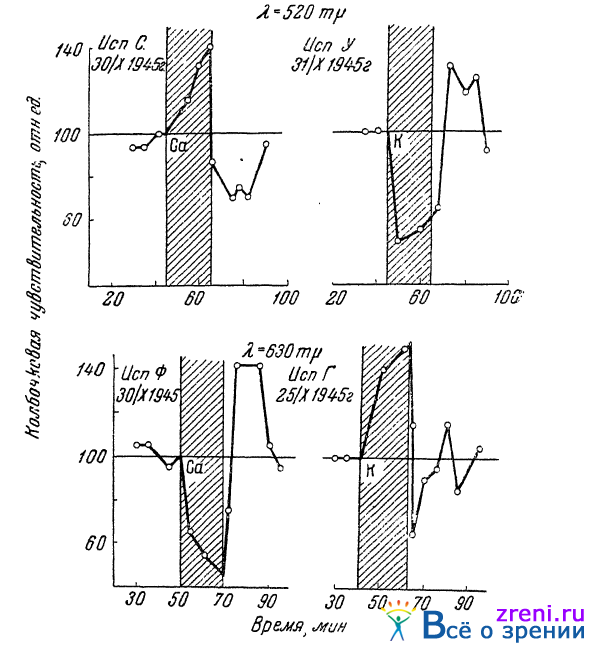
Рис. 56. Влияние ионофореза калия и кальция на колбочковую чувствительность глаза; заштрихованное пространство обозначает время ионофореза (по Кравкову и Галочкиной)
показаны типичные результаты отдельных опытов.
При изучении ионофореза ставился следующий дополнительный опыт, с нашей точки зрения весьма показательный. На один глаз помещалась подушечка, смоченная раствором калия, а на другой — подушечка, смоченная раствором кальция. Обе подушечки контактировали с положительным полюсом постоянного тока. Индифферентный электрод (катод) помещался на затылочной стороне шеи испытуемого. Пороги колбочковой чувствительности к зеленым лучам (520 m? измерялись поочередно то одним, то другим глазом. Оказалось, что в обоих глазах наступали изменения цветовой чувствительности явно обратного направления. В глазе, к которому был приложен раствор кальция, чувствительность повышалась, в глазе, к которому был приложен раствор калия, она падала. Эти данные говорят за то, что в наших опытах с ионофорезом решающую роль играло изменение условий в периферической (глазничной) части зрительного анализатора.
В специальных опытах нами было прослежено действие ионов калия (вводимых в глаз посредством ионофореза) на колбочковую чувствительность к излучениям ряда длин волн спектра (Кравков, 1947). Полученные результаты приведены на рис. 57,
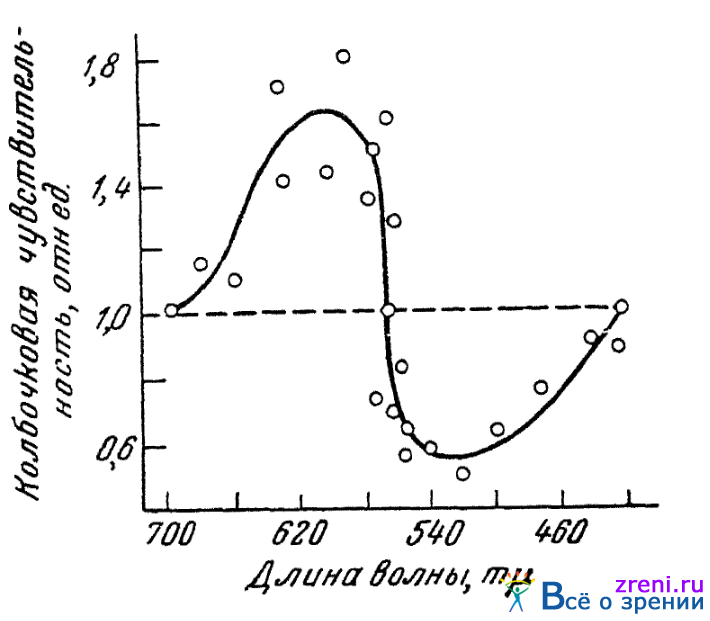
Рис. 57. Влияние ионофореза калия на колбочковую чувствительность по отношению к различным спектральным лучам (по Кравкову)
где по абсциссе показаны длины волн того света, чувствительность к которому испытывалась, а по ординате — относительное значение чувствительности, найденное при введении в глаз ионов калия; за единицу принят уровень чувствительности, установившийся в условиях темновой адаптации перед ионофорезом.
Сопоставление результатов, полученных в опытах с применением электротона и ионофореза с результатами ранее описанных опытов, позволяет видеть, что симпатикотропные раздражители и медикаменты влияют совершенно так же, как анод, или ионы кальция, раздражители же и медикаменты парасимпатикотропные меняют колбочковую чувствительность глаза подобно катоду или ионам калия. В этой установленной нами на явлениях цветового зрения закономерности нельзя не видеть подтверждения общей физиологическом закономерности, гласящей, что действие симпатического нерва в очень многих случаях подобно действию кальция, а действие парасимпатического, блуждающего, нерва подобно действию калия. Этот факт подчеркивается рядом физиологов. Например, Тинель (Tinel, 1937) пишет: «Известно, что соли кальция повышают тонус симпатической нервной системы... и увеличивают активность адреналина. Отсутствие же кальция, наоборот, может устранить и даже извратить эффект адреналина... соли калия, напротив, антагонистичны действию симпатического нерва; они повышают тонус блуждающего нерва и эффект ацетил-холина... Можно сказать, что кальций действует как симпатический нерв, а калий, как блуждающий». Это же, в итоге анализа ряда физиологических фактов, подчеркивает и Цондек (Zondek, 1923). Он приходит к выводу, что эффект калия подобен эффекту раздражения блуждающего нерва, а эффект кальция — эффекту раздражения симпатического нерва. При отсутствии ионных сдвигов не подействует и раздражение вегетативного нерва: «ионы — орудия, посредством которых нерв осуществляет свою функцию».
Было интересно далее выяснить значение для цветовой чувствительности глаз и других важных для жизни живой клетки катионов — ионов натрия и магния. Выяснению этого вопроса, поэтому, были у нас посвящены работы Поликаниной(1949). Ею показано, что ионы натрия на цветовой чувствительности глаза сказываются так же, как ионы калия, а ионы магния вызывают такие же изменения, как ионы кальция. Ионы вводились в глаз посредством ионофореза, как описано выше. Цветовая чувствительность, в условиях темповой адаптации, определялась к зеленым (520 m?) и красным (630 m?) лучам спектра. Типичные результаты опытов Поликаниной иллюстрируют данные нескольких опытов, приведенные на рис. 58.
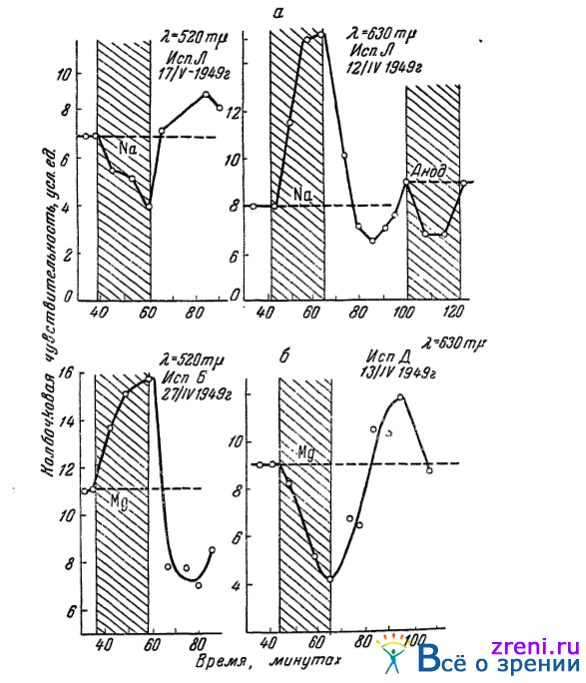
Рис. 58. Влияние ионофореза натрия (а) и магния (б) на колбочковую чувствительность глаза; время ионофореза обозначено заштрихованным пространством (по Поликаниной)
Признание решающей роли ионов в процессах зрительного возбуждения лежит, как известно, в основе ионной теории возбуждения, развивавшейся академиком П. П. Лазаревым. Согласно этой теории, начало возбуждения определяется величиною отношения С1/С2+? , где С1 и С2— концентрации антагонистических ионов, ?—некоторая постоянная. В качестве антагонистических ионов могут быть рассматриваемы одновалентные ионы К и Na, с одной стороны, и двухвалентные ионы Са и Mg, с другой. Согласно Лебу (1926), сокращения мышц наступают лишь при определенной величине ионного отношения [K]+[Na]/[Ca]+[Mg]. Можно сказать, что возбудимость, чувствительность ткани бывает (при прочих равных условиях) тем большей, чем ближе к этой оптимальной величине ионного отношения состояние ткани.
Если Лазаревым вопрос о значении ионного отношения для возбудимости зрительного прибора решался в самой общей форме и без какого-либо различения отдельных цветоощущающих аппаратов нашего зрительного анализатора, то в настоящее время мы имеем возможность сделать уже более конкретные утверждения. Вся совокупность описанных выше опытных данных позволяет признать, что порогу возбудимости зелено-и синеощущающего аппаратов глаза соответствует меньшая
[К] + [Na]/[Ca]+[Mg]
величина отношения в средах зрительного анализатора, чем порогу возбудимости красноощущающего аппарата. Чувствительность зелено- и синеощущающих аппаратов глаза обычно выигрывает от увеличения в глазе ионов Са или Mg. Чувствительность же красноощущающего аппарата, напротив, повышается при увеличении в глазе ионов К или Na. Можно поэтому в этом смысле говорить о кальцио- и магниофильности зелено- и синеощущающих аппаратов и о калио- и натриофильности аппарата красноощущающего.
Продолжение в следующей статье: Об особенностях различных цветореагирующих аппаратов зрительного анализатора. Часть 2.
---
Статья из книги: Цветовое зрение | Кравков С. В.

Комментариев 0