Передняя камера и дренажная система


Содержание:
Описание
При рассечении глазного яблока четко выявляются два отдела — передний, содержащий жидкость и находящийся впереди хрусталика, и задний, располагающийся позади хрусталика и выполненный стекловидным телом. В свою очередь, передний отдел разделяется радужкой на две камеры, переднюю и заднюю.Передняя камера глаза (camera anterior bulbi) спереди ограничена внутренней поверхностью роговицы, а по периферии — трабекулярной сетью (рис. 3.3.1, 3.3.2).

Рис. 3.3.1. Структурные образования переднего угла глазного яблока и границы лимбальной области: А — конъюнктива в области лимба; Б — влагалище глазного яблока (тенонова капсула); В - слой эписклеры; Г - склера области лимба; 1 — конъюнктивальные сосуды; 2— эписклеральные сосуды; 3 — глубокие склеральные сосуды; 4 — склеральная шпора; 5 — ресничная мышца; 6 — просвет канала Шлемма; 7— трабекулярная сеть; 8 — отростки радужной оболочки, переходящие в трабекулы; 9 — место прерывания боуменовой оболочки; 10 — место прерывания десцеметовой оболочки
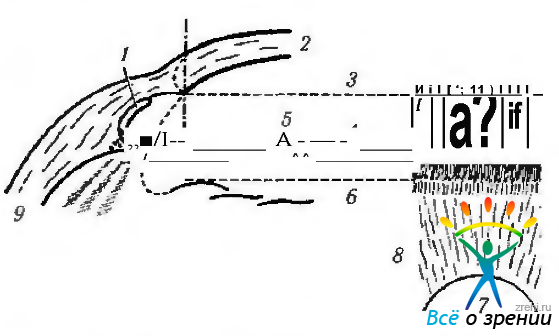
Рис. 3.3.2. Соответствие гониоскопической картины особенностям микроскопического строения структур угла передней камеры (по Fine, Yanoff, 1972): 1 — шлеммов канал; 2— роговица; 3 — линия Швальбе; 4 — трабекулярная сеть; 5 — склеральная шпора; 6 — рецессия угла; 7— зрачок; 8— передняя поверхность радужки; 9 — склера
Сзади она в пределах зрачка ограничена хрусталиком и передней поверхностью радужки, а по периферии — передней поверхностью ресничного тела. Передняя и задняя границы передней камеры глаза встречаются в углу дренажной системы. Передняя камера сообщается через зрачок с задней камерой глаза.
Объем передней камеры примерно равен 220 мкл, и средняя глубина — 3,15 мм (2,6—4,4 мм). Диаметр передней камеры колеблется от 11,3 до 12,4 мм.
Глубина камеры может быть различной, что хорошо выявляется при использовании гониоскопии. Когда угол между задней поверхностью роговой оболочки и передней поверхностью радужки менее 20°, камеру называют узкой. При этом высока вероятность контакта радужки с трабекулярной сетью, приводящего к блокаде дренажной системы.
Отмечено, что глубина камеры уменьшается на 0,01 мм в год. В гиперметропическом глазу это уменьшение выражено в большей степени, чем в близоруком (камера углубляется на 0,06 мм для каждой диоптрии в близоруком глазном яблоке). Отмечается изменение глубины камеры и при аккомодации. Это связано с увеличением кривизны передней поверхности хрусталика и его смещением кпереди.
Переходя к описанию строения системы оттока камерной влаги, необходимо первоначально остановиться на структурах, образующих дренажную систему (рис. 3.3.1, 3.3.2).
Край (лимб) роговицы (limbus соrпеае) представляет собой переходную зону шириной приблизительно 1,5 мм. Располагается эта зона между роговой оболочкой и склерой. Границей лимба является линия, соединяющая конец боуменовой оболочки и места прерывания десцеметовой мембраны. По периферии корнеосклеральное соединение отграничено параллельной линией, проходящей через склеральную шпору.
Лимб можно разделить на три слоя в зависимости от глубины расположения структур. Это «глубокие слои», в состав которых входят шлеммов канал и трабекулярная сеть; «средние слои», состоящие из «корнеосклеральной стромы», в которой располагается также интрасклеральное венозное сплетение. В состав «поверхностных слоев» входят эписклера, тенонова капсула, строма и эпителий конъюнктивы.
Существует еще ряд подходов в определении понятия лимба. Патологоанатомы считают задней границей лимба линию, проходящую в 1,5 мм от места прерывания боуменова слоя. «Хирургический» лимб имеет ширину 2 мм и может быть разделен на две зоны: переднюю светло-серую зону, надлежащую над прозрачной роговой оболочкой и распространяющуюся от боуменовой оболочки до линии Швальбе, и заднюю белую зону, надлежащую над трабекулярным аппаратом и распространяющуюся от линии Швальбе до склеральной шпоры или корня радужной оболочки. Эти ориентиры необходимо знать при экстракции катаракты и проведении антиглаукоматозных операций.
↑ Клиническая анатомия передней камеры
В норме угол передней камеры глаза не виден, поскольку наблюдается полное внутреннее отражение идущего от угла света передней поверхностью роговой оболочки. Специальные оптические системы (гониоскопы прямые и непрямые) позволяют увидеть угол передней камеры, что широко и используется в клинической практике. При помощи гониоскопа клинически можно увидеть ряд ориентиров, характеризующих структурные особенности угла (рис. 3.3.1—3.3.4).
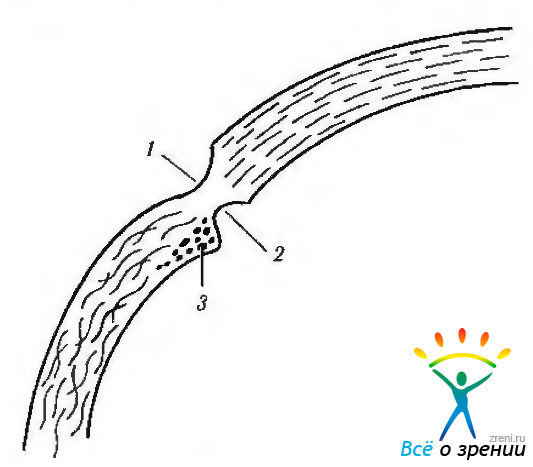
Рис. 3.3.3. Меридианальный срез корнеосклеральной области: 1 — наружная склеральная борозда; 2— внутренняя склеральная борозда; 3 — склеральная шпора

Рис. 3.3.4. Топография образований угла передней камеры (а) и их микроскопическое строение (б): 1 — венозный синус склеры (пшеммов канал); 2 — юкстаканали-купярная сеть; 3— задняя пограничная пластинка (десцеметова мембрана); 4 — корнеосклеральная часть трабекулярной сеточки; 5 — увеальная часть трабекулярной сеточки; 6 - склеральная шпора; 7 — корень радужной оболочки
Ресничная связка, являющаяся наиболее задней отметкой угла и представляющая собой темную полосу, соответствующую передней поверхности ресничного тела, и места прикрепления ресничной мышцы к склеральной шпоре.
Склеральная шпора выглядит тонкой светлой узкой полосой, расположенной между поверхностью ресничного тела и пигментированной зоной трабекулярной сети. Эта полоса очерчивает заднюю границу корнеосклеральной части трабекулярной сети.
Трабекулярная сеточка при гониоскопии располагается кпереди от склеральной шпоры и представляет собой широкую (750 мкм) полосу, довольно трудно различимую в слабо пигментированном глазе. Распространяется она от склеральной шпоры к кольцу Швальбе. Трабекулярная сеть прикрывает внутреннюю часть шлеммова канала.
Венозный синус склеры (шлеммов канал) (sinus venosus sclerae Schlemm) можно увидеть при гониоскопии только тогда, когда происходит ретроградный заброс в него крови (ре-флюкс). При этом канал выглядит розовой полосой, просвечивающей через трабекулы. Ретроградный заброс крови возможен при гониоскопии, поскольку при наложении на поверхность глаза гониоскопа затрудняется эписклеральный венозный дренаж и изменяется направление кровотока.
Пограничное кольцо (линия) Швальбе представляет собой переднюю границу дренажного утла. Она выглядит как нежная зубчатая линия, расположенная в месте прерывания мембраны Десцемета. Примерно у 15—20% людей эта линия может быть значительно утолщенной и проецироваться в виде тонкого блестящего гребня в переднюю камеру (задний эмбриотоксон). Кольцо Швальбе иногда слегка пигментировано.
«Углубление» угла (recess). Верхушка утла передней камеры глаза находится в плоскости, расположенной позади на 0,6—1,0 мм наиболее передней точки капсулы хрусталика. Поэтому радужка изгибается назад, образуя «углубление» утла передней камеры. Ширина этого «углубления» зависит от размера глаза, глубины передней камеры, состояния зрачка и других факторов.
Таким образом, при помощи гониоскопии можно определить состояние ряда образований —
- трабекулярной сети,
- радужной оболочки,
- ресничного тела,
- задней поверхности роговой оболочки,
- склеральной шпоры,
- зрачка.
Передняя камера глаза содержит структуры, обеспечивающие дренаж камерной влаги. Большая часть влаги оттекает через трабекулярную сеть в шлеммов канал, а затем в интра- и эписклеральные венозные сосуды. Появление препятствия на этом пути оттока приводит к повышению внутриглазного давления, состоянию, называемому глаукомой.
В тех случаях, когда передняя камера мелкая, повышение внутриглазного давления возможно при смещении корня радужки вперед. При этом происходит блокада утла. Подобное состояние называется первичной закрытоугольной глаукомой.
При другой форме глаукомы, так называемой первичной открытоугольной глаукоме, отток камерной влаги затруднен в связи с появлением препятствия оттоку влаги на уровне трабекулярной сети и шлеммова канала. В этом угол остается открытым.
↑ Дренажный аппарат
Дренажный аппарат состоит из:
- внутренней склеральной борозды;
- трабекулярной сети;
- шлеммова и коллекторных каналов.
Внутренняя склеральная борозда (склеральный валик) представляет собой расположенное циркулярно углубление на внутренней поверхности лимба (рис. 3.3.4—3.3.6).
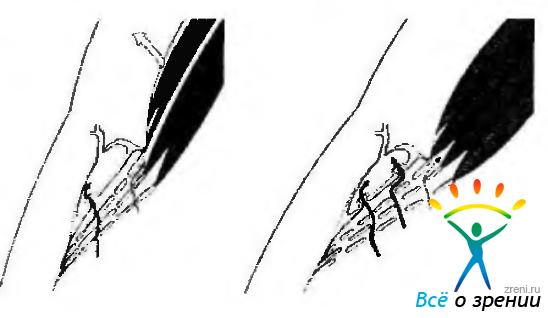
Рис. 3.3.5. Изменение проходимости дренажной системы при расслаблении (а) и сокращении (б) ресничной мышцы: сокращение мышцы приводит к ее утолщению, что сопровождается уменьшением пространства между мышечными волокнами и уменьшением объема увеасклерального пути оттока. В то же время сокращение мышцы приводит к натяжению склеральной шпоры и расширению, пространств между трабекулами, что способствует уменьшению резистентности трабекулярной сети оттоку камерной влаги
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Рис. 3.3.6. Строение дренажной системы при использовании сканирующей электронной микроскопии (по Fine, Yanoff, 1972): 1 — роговица; 2 — задняя поверхность роговицы; 3 — коллекторный канал; 4 — шлеммов канал; 5 — угол передней камеры; 6 — радужка; 7 — радужка на срезе; 8—пигментный эпителий радужки
Задней границей внутренней склеральной борозды являются пучки циркулярным образом расположенных коллагеновых волокон, которые формируют склеральную шпору или задний пограничный круг Швальбе. В борозде снаружи размещается шлеммов канал, а кнутри — «корнеосклеральная часть» трабекулярной сети (рис. 3.3.4—3.3.6).
Кольцо Швальбе, как указывалось выше, является передней границей трабекулярной области. Здесь коллагеновые волокна перемешиваются с эластическими волокнами. С возрастом появляются и спиралевидные коллагеновые волокна Кольцо Швальбе является местом перехода эндотелия роговой оболочки к клеткам, покрывающим трабекулы.
Склеральная шпора представляет собой клиновидный гребень, обращенный в сторону полости глаза и состоящий из циркулярным образом ориентированных коллагеновых и эластических волокон (рис. 3.3.4). К склеральной шпоре присоединяется сухожилие продольной ресничной мышцы. Здесь же присоединяется корнеосклеральная часть трабекулярного аппарата. Переднемедиальный край шпоры образует задний край склеральной борозды (рис. 3.3.4).
Коллагеновые волокна склеральной шпоры различного диаметра (от 35 до 80 нм). Толщина их увеличивается по мере приближения к склере.
Сокращение ресничной мышцы оттягивает склеральную шпору кзади. При этом открываются межтрабекулярные пространства (рис. 3.3.5). Предполагают, что этот механизм является одним из основных механизмов понижения внутриглазного давления при применении миотиков.
Недавно было показано, что в пределах склеральной шпоры имеются сократительные миофибробластоподобные клетки, в цитоплазме которых выявлено большое количество а-актинина и миозина. В клетках недостает десмина и микрофилламентов промежуточного типа, т. е. компонентов, характерных гладкомышечным клеткам ресничного тела. Миофибробластоподобные клетки склеральной шпоры контактируют с эластическими волокнами склеральной шпоры, а некоторые из них непрерывно переходят в смежные участки трабекулярной сети. Часть клеток трабекулярной сети также содержит а-актинин и актин гладких мышц. Таким образом, можно предположить, что сокращение этих клеток может изменять архитектонику трабекулярной сети и изменять сопротивление оттоку камерной влаги.
К некоторым миофибробластоподобным клеткам склеральной шпоры подходят безмякотные аксоны нейронов, тела которых лежат в супрацилиарном пространстве. Аксоны распространяются в склеральной шпоре циркулярно и параллельно соединительнотканным элементам. Их терминалы плотно контактируют с клеточными мембранами. Окончания нервов содержат зернистый материал и а гранулярные пузырьки, напоминающие таковые в адренэргических нервах. Тем не менее волокна не относятся к адренэргическим, что подтверждено иммуногистохимически. Tamm et al. выявили, что подобного типа пузырьки обнаруживаются в неадренэргических терминалах нервной системы кишечника.
Необходимо принять во внимание то, что аксоны, иннервирующие миофибробластоподобные клетки склеральной шпоры у человека, относятся к аминэргическим, пептидэргическим и нитрэргическим. Они в то же время не дают положительной реакции при проведении иммуногистохимической реакции для выявления ацетилхолинэстеразы. Важно отметить и то, что задние участки трабекулярной сети иннервируются аналогичным образом. Парасимпатические пептидэргические и нитрэргические волокна, подходящие к склеральной шпоре, исходят из крылонебного ганглия, а также нервных волокон сосудистой оболочки.
Предполагают, что пептидэргическая и нитрэргическая иннервация миофибробластоподобных клеток является основной в регуляции сопротивления оттоку камерной влаги посредством контакта миофибробластоподобных клеток с эластическими волокнами трабекулярной сети. Введение обезьянам нитровазодилятаторов вызывает увеличение оттока камерной влаги.
Трабекулярная сеточка (зубчатая связка; reticulum trabeciilare; lig. pectinatum; spongium iridocorneale).
На меридианальных срезах глаза видна скудная коллагеновая сеть, выполняющая внутреннюю склеральную борозду и распространяющаяся к корню радужки в виде веера (рис. 3.3.1, 3.3.4). Ручка этого веера располагается несколько кпереди от места прерывания десцеметовой мембраны. Именно в этом месте коллагеновые волокна веера проникают в глубокие периферические слои стромы роговицы и переплетаются с ними.
Трабекулярную сеть можно разделить воображаемой линией на две части. Эту линию необходимо провести от склеральной шпоры к месту прерывания десцеметовой мембраны. Часть трабекулярной сети, лежащую снаружи линии и расположенную между роговой оболочкой и склерой, обозначают роговично-склеральной частью (pars corneoscleralis reticulum trabeciilare). Часть трабекулярной сети, расположенную кнутри и прилежащую к радужной оболочке, обозначают сосудистооболочечной (увеальной) частью (pars uvealis) (рис. 3.3.1, 3.3.4).
Ширина трабекулярной сети сзади, вблизи склеральный шпоры, равняется 120—180 мкм. Она более широкая при близорукости, чем при гиперметропии.
Между корнеосклеральной частью трабекулярной сети и эндотелиальнои выстилкой шлеммова канала располагается богатая клетками зона — пери- или юкстаканаликулярная соединительная ткань.
Пространства радужно-роговичного угла, расположенные между трабекулами (фонтановы пространства; spatia anguli iridocornealis Fontana), содержат гидрофильные гликозаминогликаны и коллагеновый материал, которые влияют на отток камерной влаги.
Сосудистооболочечная (увеальная) часть трабекулярной сети. Внутренняя часть увеальной трабекулярной сети (1—2 слоя) состоит из переплетающихся трабекул. Самые внутренние трабекулы могут распространяться от ресничной мышцы к кольцу Швальбе. Сзади определяется 2—5 слоев трабекул, внешние слои которых ориентированы циркулярно.
Сзади, трабекулы могут соединяться с циркулярными и радиальными мышечными волокнами ресничной мышцы. Спереди «увеальные трабекулы» постепенно сближаются, и заканчиваются в месте прерывания десцеметовой мембраны, т.е. внутренней части кольца Швальбе. Эндотелиальное покрытие трабекул постепенно переходит в эндотелий роговой оболочки.
Трабекула увеальной части трабекулярной сети имеет диаметр 4—6 мкм. Она утолщается кзади и сужается кпереди. Ширина межтрабекулярных пространств колеблется от 20 до 75 мкм.
Роговично-склеральная часть трабекулярной сеточки. Роговично-склеральная часть трабекулярного аппарата представляет собой решетчатую уплощенную структуру, состоящую из трабекул. Толщина каждой трабекулы приблизительно 5—12 мкм. Расстояние между трабекулами равняется 5—20 мкм. При этом меж-трабекулярные пространства внешних слоев роговично-склеральной части колеблются между 2 и 20 мкм, т. е. пространства более узкие, чем в увеальной части.
Между трабекулами, расположенными на разных уровнях, обнаруживаются межтрабекулярные «связки», толщиной от 2 до 5 мкм.
Количество слоев трабекул в роговичносклеральной части колеблется от 8 до 15, а общая ее толщина равна 120—150 мкм. Передние слои роговично-склеральной части трабекулярного аппарата сходятся и сливаются с роговичными пластинами.
Трабекула. Основной структурой увеальной и роговично-склеральной частей трабекулярного аппарата являются трабекулы. В трабекуле различают кортикальную зону и стержень. Снаружи трабекула покрыта одним слоем клеток (рис. 3.3.7—3.3.9).
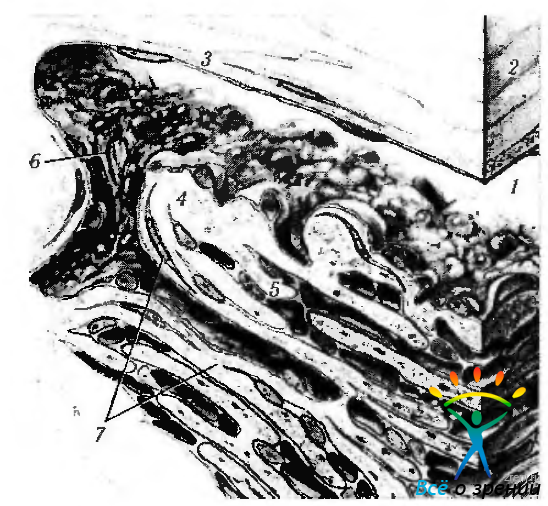
Рис. 3.3.7. Трехмерное схематическое изображение венозного синуса склеры (шлеммова канала) и трабекулярной сети (по Hogan et al., 1971): 1 — просвет канала; 2 — эндотелиальная клетка; 3 — наружная стенка канала; 4 — внутренняя стенка канала; 5 — межтрабекулярные пространства; 6 — внутренние соединительные каналы; 7—корнеосклеральные трабекулы
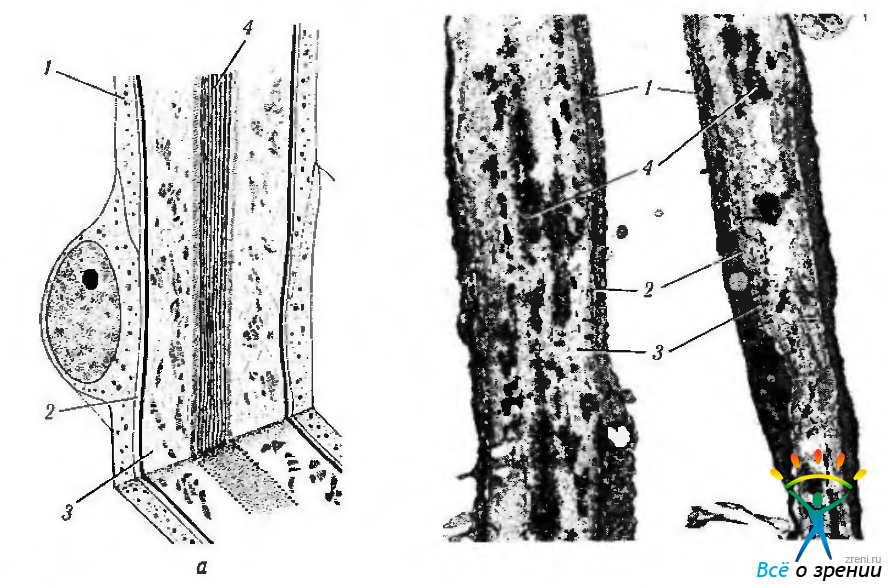
Рис. 3.3.8. Схематическое изображение структурной организации (а) и электронномикроскопическое строение (б) трабекулы: 1 — эндотелиальная клетка; 2— базальная мембрана; 3— кортикальная зона; 4 — стержень трабекулы

Рис. 3.3.9. Ультраструктурная организация юкстаканаликулярной соединительной ткани (по Fine, Yanoff, 1972): 1 — венозный синус склеры (шлеммов канал); 2 — эндотелиальные клетки, выстилающие шлеммов канал: 3 — юкстаканаликулярная сеть; 4— межтрабекулярные пространства; 5 — трабекула
Клетки трабекулы располагаются вдоль длинной оси трабекулы. Толщина их порядка 4—8 мкм, а длина 120 мкм. Соседние клетки контактируют между собой посредством отростков. Они также соединяются при помощи десмосом и щелевых контактов. Несмотря на наличие межклеточных контактов, радиоактивные трейсеры (ферритин) свободно проникают вглубь трабекулы по межклеточным пространствам.
Поверхность трабукулярных клеток покрыта макромолекулами, богатыми сиаловыми кислотными остатками. Между трабекулами гиалуроновый гель не обнаруживается.
Трабекулярные клетки содержат обычные органоиды и большое количество пиноцитозных пузырьков. Обнаруживаются также филаменты цитоскелета. Клетки трабекул отличаются высокой синтетической активностью. Они синтезируют материал базальных мембран, коллаген и гликозаминогликаны.
Наиболее важной функцией трабекулярных клеток является их барьерная функция на пути камерной влаги. Эта функция обеспечивается структурными особенностями клеток и зависит от биологической их активности. Одной из функций является также синтез межклеточного материала и его лизис. Последняя функция вытекает из необходимости постоянного лизиса материала, освобождающегося в трабекулярной сети по мере прохождения через нее камерной влаги. О синтетической активности клеток свидетельствуют экспериментальные исследования по культивированию изолированных клеток in vitro. Трабекулярные клетки при этом синтезируют внутри- и внеклеточные гликозаминогликаны (гепарансульфат, гиалуроновая кислота, дерматансульфат).
Получены убедительные данные, свидетельствующие о способности трабекулярных клеток синтезировать волокнистый материал, особенно после травмы или применения кортикостероидов. Трабекулярные клетки обладают также фибринолитическими свойствами. В культуре ткани трабекулярные клетки синтезируют в определенном количестве активатор плазминогена.
Трабскулярныe клетки обладают высокой фагоцитарной активностью. Нередко в них можно найти зерна пигмента и другие частицы, количество которых увеличивается с возрастом. Введенные в эксперименте частицы (коллоидное золото, пероксидаза хрена, витальные красители) моментально фагоцитируются клетками и, таким образом, выводятся из камерной влаги. Для переваривания фагоцитированного материала цитоплазма трабекулярных клеток содержит достаточно большое количество лизосом. У некоторых животных (кошка) после фагоцитоза трабекулярные клетки гибнут и восстанавливаются только спустя 150 дней, а у человека поглотившие пигмент клетки сохраняются длительно.
В последние годы установлено, что трабекулярные клетки синтезируют многочисленные биологически активные вещества, некоторые из которых участвуют в регуляции внутриглазного давления. К ним относятся простагландин F2, ингибитор тканевой и матричной металлопротеиназы. Причем увеличивается синтез этих веществ, и они высвобождаются в камерную влагу при механической деформации клеток, что происходит при колебаниях внутриглазного давления.
Интересные данные были получены при изучении синтеза в трабекулярных клетках оксида азота, вещества, обладающего многими функциями. Окись азота обладает иммуномодулирующим свойством, участвует в процессах сокращения и расслабления мышечной ткани, обладает нейромодуляторными свойствами. Образуется окись азота благодаря ферментативной активности синтетазы оксида азота, которая генерирует окисль азота из L-аргинина и является короткоживущим свободным радикалом. Показано, что интенсивность синтеза оксида азота зависит от колебания внутриглазного давления. Колебания давления деформируют трабекулярные клетки, что и является причиной активации синтетазы оксида азота. Такая связь между активацией синтеза оксида азота и обратимой деформацией клеток свойственна не только трабекулярным клеткам. Она характерна для эндотелиальной выстилки шлеммова канала, эндотелиальных клеток сосудов, хондроцитов, остеоцитов.
Оксид азота способен расслаблять трабекулярную сеть и ресничную мышцу. Происходит это благодаря существованию различных механизмов. Так, оксид азота, синтезируемый трабекулярными клетками, может включать гуанилил циклазу и различные аутокринные и паракринные механизмы, приводя к увеличению концентрации циклического GMP в трабекулярных клетках. О роли оксида азота, синтезируемого трабекулярными клетками, свидетельствуют наблюдения снижения активности синтетазы оксида азота при глаукоме.
Непосредственным механизмом влияния оксида азота на регуляцию внутриглазного давления является его влияние на концентрацию в цитоплазме трабекулярных клеток ионов кальция. Показано, что в трабекулярных клетках при повышении ВГД до 20—30 мм Hg изменяется концентрация внутриклеточного кальция, поскольку синтетаза оксида азота (bNOS и eNOS) активизирует комплекс кальций / кальмодулин (комплекс кальция с кальмодулином является месседжером. изменяющим активность многих ферментов, регулирующих кальциевый насос, различные специфические белковые киназы, циклические нуклеотидные фосфодиэстеразы, гистоны и тубулин).
Кортикальная зона. Кортикальная зона состоит из окрашивающегося положительно ШИФФ-реактивом пластинчатого материала, присоединенного к трабекулярным клеткам при помощи полудесмосом. Внутренняя граница этой зоны не очень хорошо видна и инфильтрирована соединительнотканными элементами коры.
В пределах базальной пластинки найдены скопления веретенообразных коллагеновых волокон с периодичностью, колеблющейся от 30—40 до 80—120 нм.
Стержень. Стержень каждой трабекулы образован коллагеном I, II и IV типов. Он также содержит фибронектин, эластин, хондроитинсульфат, дерматансульфат и спиралевидный коллаген.
Коллагеновые фибриллы (толщина 30— 50 нм) ориентируются вдоль длинной оси трабекул. В трабекулах увеальной части они формируют компактный стержень. Ориентация коллагеновых фибрилл в трабекулах, вероятно, определена направлением приложения силы при сокращении мышц ресничного тела.
В «увеальной» трабекуле эластические волокна располагаются, главным образом, в центре стержня. Эти эластические волокна отличаются по строению от эластических волокон других тканей организма. Состоят они из волокнистого и аморфного компонентов. В этой области иммуногистохимически определяется большое количество микрофибриллярного белка, близкого к эластину,— фибриллина. Ультраструктурно показано, что только центральная зона эластического волокна содержит эластин и тропоэластин. Эти компоненты погружены в электронноплотный материал неизвестной природы.
Эластический компонент трабекулы играет определенную роль в способности трабекулы к сокращению, что было показано на изолированной трабекуле быка.
Сокращению способствует наличие в трабекулярных клетках миофиламентов (актин). Именно эта особенность позволяет отнести трабекулярные клетки к миофибробластам. Показано, что у человека количество таких клеток уменьшается с возрастом. Сохраняются они лишь вблизи склеральной шпоры.
Особого внимания заслуживают вопросы возрастных изменений трабекулярных клеток. С возрастом пролиферативная активность трабекулярных клеток снижается. Кроме того, на протяжении жизни количество клеток постоянно линейно уменьшается со скоростью потери 0,56% клеток в год. Количество клеток у 20-летнего индивидуума равняется примерно 763 000, а у 80-летнего — всего лишь 403 000. При этом количество клеток уменьшается ежегодно примерно на 6000. Интересно, что скорость потери трабекулярных клеток различна в различных участках трабекулярной сети. Наибольшая потеря клеток отмечается в центральных участках.
Немаловажное практическое значение имеет выявление репаративных возможностей трабекулярной сети. Трабекулярные клетки in vitro не способны регенерировать. Тем не менее при повреждении трабекулярной ткани отмечаются признаки регенерации клеток, принимающих кубовидную форму. При этом увеличивается их число. Подобную регенерацию трабекулярных клеток выявляли после трабекулоэктомии или после лазерной трабекулопластики.
Отростки радужной оболочки представляют собой однородные треугольной формы «связки», идущие от корня радужки до трабекул «увеальной» части трабекулярного аппарата. с которыми они и сливаются. Иногда отростки достигают склеральной шпоры, а иногда и линии Швальбе. Количество их незначительно. Обнаруживаются они у трети индивидуумов. У индивидуумов с карими глазами эти отростки пигментированы. Строение отростков аналогично строению стромы радужки. Иногда отростки прикрывают угол передней камеры.
Клетки Швальбе. В месте перехода между роговой оболочкой и трабекулярной сетью рядом исследователей обнаружены клетки, отличающиеся хорошо выраженной эндоплазматической сетью, большим количеством митохондрий и многочисленных электронноплотных гранул. Эти клетки были названы клетками Швальбе. Предполагают, что клетки Швальбе обладают секреторной активностью, о чем свидетельствует не только обнаружение в цитоплазме гранул, дающих положительную реакцию при выявлении нейрон-зависимой энолазы, гиулоронат-синтетазы. Происхождение и функция этих клеток пока неизвестны.
Пери- или юкстаканаликулярная соединительная ткань распространяется вдоль всего шлеммова канала (рис. 3.3.9). Толщина этой зоны колеблется от 2,0 до 20,0 мкм, и располагается она между эндотелиальной выстилкой канала и лежащей кнутри «корнеосклеральной» частью трабекулярного аппарата. Эта зона складывается из 2—5 клеточных слоев, погруженных в межклеточное вещество (рис. 3.3.9). Клетки обладают длинными отростками и соединяются между собой при помощи зон замыкания, десмосом и щелевых контактов. Между клетками определяются промежутки шириной 10 мкм, через которые проникает камерная влага по направлению эндотелиальной выстилки шлеммова канала. Между этими клетками и эндотелием шлеммова канала располагается базальная мембрана.
Периканаликулярные клетки обладают важными функциями — фагоцитарной и синтетической. Эта ткань представляет собой наиболее мощное препятствие на пути оттекающей влаги передней камеры глаза. Связано это не только с тем, что межклеточные пространства узкие и извилистые, но, в первую очередь, с присутствием внеклеточно расположенных протеогликанов и гликопротеидов.
Зона, контактирующая с внешней стенкой шлеммова канала, содержит меньше клеток, чем прилегающая к ней трабекулярная ткань. Состоит она из 4—8 плотно упакованных слоев фиброцитоподобных клеток. Толщина этой зоны порядка 5—15 мкм Помимо клеток, в ней определяются неравномерно распределенные коллагеновые, эластические волокна и мелкозернистый материал. Коллаген относится к VI типу.
Имеется также и переходная зона, толщиной 20—30 мкм, располагающаяся между этой юкстаканаликулярной тканью и склерой. Она состоит примерно из 10 коллагеновых пластин, практически идентичных склеральным пластинам.
Межклеточное вещество. Главными компонентами межклеточного вещества являются коллаген I, III, IV, V и VI типов, фибронектин, хондроитин- и дерматансульфат. Обнаруживается также гиалуроновая кислота и эластическая ткань. Многие из этих макромолекул (коллаген VI типа, фибронектин, хондроитин- и дерматансульфат) содержат сиаловую кислоту. Обнаружен и фибриллин. Особенностью межклеточного вещества является наличие эластических волокон, образующих густую объемную сеть («решетчатое сплетение»). Поскольку эластические волокна связаны с сухожилиями мышцы ресничного тела и базальной мембраной эндотелиальных клеток шлеммова канала, они могут влиять на проходимость этой области для камерной влаги.
↑ Шлеммов канал и коллекторные каналы
Венозный синус склеры (шлеммов канал; sinus venosus sclerae Schlemm). Шлеммов канал (Schlemm, 1830) представляет собой узкую трубку или систему трубок длиной 36 мм (рис. 3.3.7, 3.3.10, 3.3.11).

Рис. 3.3.10. Сканограмма вскрытого венозного синуса склеры (шлеммов канал). Стрелками указаны трабекулы
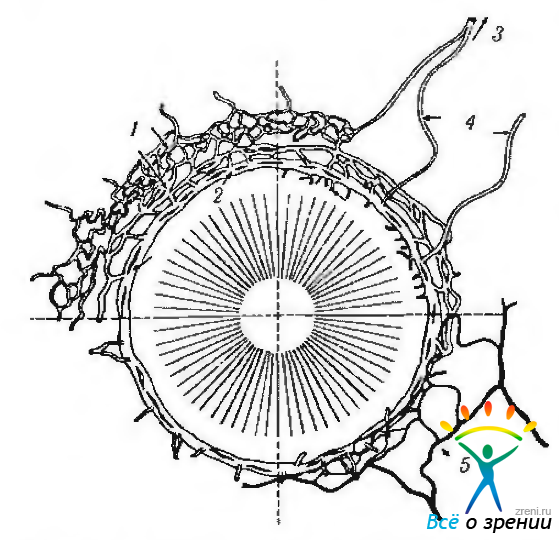
Рис. 3.3.11. Схематическое изображение венозного синуса склеры (шлеммового канала) и его отношение к артериальной и венозной системам (по Tripathi et ai, 1982): 1 — интрасклеральное венозное и глубокое склеральное сплетения; 2 — шлеммов канал; 3 — эписклеральное венозное сплетение; 4 водяные вены; 5 артериальный круг
Внутренняя ее стенка выстлана эндотелием. Шлеммов канал располагается в наружной части внутренней склеральной борозды. Его основной функцией является отведение камерной влаги из трабекулярной сети в эписклеральную венозную сеть посредством коллекторных каналов. Юкстаканаликулярная соединительная ткань отделяет внутренние и внешние стенки шлеммова канала от трабекулярной сети и склеры.
Просвет шлеммова канала на поперечном разрезе овальной формы. Он может быть разделен перегородками на отделы и состоять из многочисленных каналов.
Ширина шлеммова канала в поперечных плоскостях 120—400 мкм и 10—25 мкм. Существуют довольно широкие колебания размеров шлеммова канала в зависимости от возраста, наличия предшествоваших заболеваний глаза, что необходимо учитывать при проведении антиглаукоматозных операций.
Эндотелиальная выстилка шлеммова канала располагается на базальной мембране, которая местами прерывается. Подобный характер базальной мембраны позволяет предположить, что мембрана не может обеспечить существенного сопротивления потоку камерной влаги.
Главными компонентами базальной мембраны являются коллаген IV типа, ламинин, фибронектин, гепаран сульфат протеогликан.
В шлеммовом канале видны отростчатые расширения в виде дивертикулов, направленные в сторону юкстаканаликулярной ткани и трабекулярного аппарата (каналы Сондерманна).
Эндотелиальная выстилка. Стенка шлеммова канат, обращенная в сторону глаза. На протяжении длительного времени продолжались споры относительного того, существует или нет прямое сообщение между передней камерой и шлеммовым каналам. Теперь точно известно, что передача влаги осуществляется посредством переноса ее через цитоплазму эндотелиальных клеток. Морфологическим проявлением этого процесса является присутствие в цитоплазме эндотелиоцитов вакуолей.
Шлеммов канал выстлан одним слоем эндотелиальных клеток. На внутренней поверхности канала они имеют длину 40—120 мкм, ширину 4—12 мкм, а толщину 0,2 мкм. Скреплены они между собой при помощи десмосом. Встречаются и редкие щелевые контакты, располагающиеся между эндотелиальными клетками и клетками юкстаканаликулярной ткани.
Межклеточные контакты занимают незначительную площадь мембраны. Они не могут предотвратить прохождение лейкоцитов или макрофагов. Плотность расположения межклеточных контактов не изменяется при изменении внутриглазного давления.
На апикальной поверхности эндотелиальных клеток видны микроворсинки. В цитоплазме эпителиоцитов содержатся многочисленные свободные рибосомы и микрофиламенты, а также множество пиноцитозных пузырьков.
Наиболее явной особенностью внутренней стенки шлеммова канала является наличие гигантских вакуолей. Ширина их от 4 до 6 мкм, а длина до 25 мкм. Возникают они в результате инвагинации базальной плазматической мембраны эндотелиальных клеток, обеспечивая, таким образом, возможность проникновения камерной влаги в юкстаканаликулярную ткань.
Меньшая часть влаги может проникать через поры, образованные в цитоплазме клеток («трансцеллюлярные каналы»). Поры могут быть до 2,5 мкм в диаметре, в то время как базальные инвагинации имеют ширину до 4 мкм. Плотность расположения пор в норме равняется 850 пор/мм2 (Johnson et al., 2002). причем их плотность у меньшается при развитии глаукомы.
Использование меченных изотопами веществ и частиц различного диаметра позволило выяснить, что многие вещества могут проходить через «трансцеллюлярные каналы» из передней камеры в шлеммов канал (рис. 3.3.12).
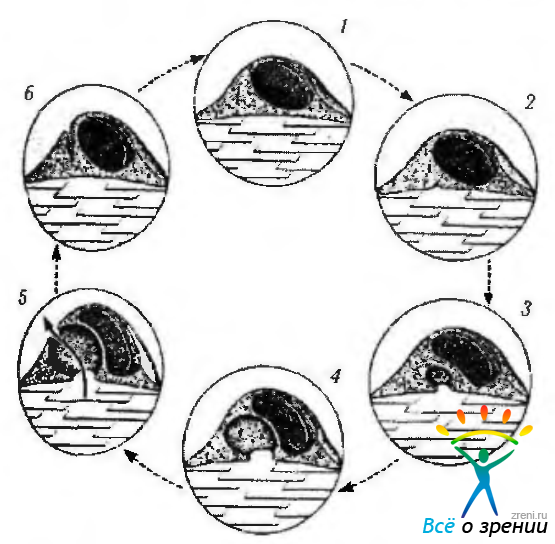
Рис. 3.3.12. Схематическое изображение концепции Tripathi et al. (1977) относительно механизма формирования трансцеллюлярных каналов в эндотелиальных клетках шлеммова канала при выведении камерной влаги (цикл образования вакуолей в эндотелиальных клетках)
Эта возможность выявлена для торотраста, ферритина, золота и пероксидазы хрена. Через эти каналы могут проходить даже такие клетки, как эритроциты.
Выявлена закономерность, которая сводится к тому, что формирование вакуолей в эндотелиальных клетках зависит от уровня внутриглазного давления. Причем при нарастании давления число вакуолей увеличивается.
Tripathi считает, что при увеличении внутриглазного давления в эндотелиальных клетках внутренней стенки шлеммова канала появляется способность «циклически» пропускать камерную влагу, образуя внутрицитоплазматические вакуоли и «трансцеллюлярные каналы». До сих пор непонятно, является ли этот процесс активным, использующим энергию, или протекает пассивно. Тем не менее важно знать, что камерная влага поступает в шлеммов канал только через эндотелиальные клетки и только 1% общего объема влаги проникает между эндотелиальными клетками. При этом вся эндотелиальная выстилка шлеммова канала обеспечивает только 5—10% сопротивления оттоку камерной влаги.
Эндотелиальные клетки наружной стенки шлеммова канала более длинные и более плоские. Апикальная их поверхность гладкая. Они прочно соединены между собой при помощи зон замыкания. В цитоплазме клеток редко выявляются гигантские вакуоли. Лежат эндотелиоциты на толстой базальной мембране. В соответствии с особенностями строения наружной стенки можно предположить, что ее пропускная способность низкая. Тем не менее использование изотопных меток выявило высокую пропускную способность.
Коллекторные канты. Коллекторные каналы в количестве 25—35 начинаются у внешней стенки шлеммова канала (рис. 3.3.11). Посредством этих каналов влага оттекает в три венозных сплетения: глубокое, среднее склеральное и эписклеральное. До 8 каналов отводят влагу непосредственно в эписклеральное венозное сплетение. Известны эти каналы как «водяные вены». Они были обнаружены Ашером (Ascher) в 1942 году, а их связь со шлеммовым каналом выявлена Эштоном.
При помощи щелевой лампы «водяные вены» видны в виде прозрачных сосудов, содержащих как камерную влагу, так и кровь. Наиболее часто их можно обнаружить субконъюнктивально на расстоянии 2 мм от лимба книзу и назально. Перед впадением в эписклеральные вены они распространяются на протяжении 1,0—10,0 мм. Коллекторные каналы выстланы эндотелием. Клапаны в них отсутствуют.
Глубоко расположенное склеральное венозное сплетение представлено ветвями передних ресничных вен, которые соединяются со средним склеральным сплетением. При этом в лимбальной области образуется интрасклеральная венозная сеть. Эта система получает кровь также и от ресничного венозного сплетения.
Из интрасклерального сплетения влага оттекает в эписклеральное сплетение и далее к передним ресничным венам. Эписклеральное венозное сплетение, кроме того, получает кровь от вен конъюнктивы, дренирующих перилимбальную область.
Кровоснабжение дренажной системы. Кровоснабжается шлеммов канал сосудами малого крута кровообращения радужки, получающего, в свою очередь, ветви из поверхностных и глубоких ответвлений передних ресничных артерий. Иногда артериолы проходят вблизи шлеммова канала, отделенные от него только адвентицией.
Иннервация дренажной системы. Иннервация дренажной системы осуществляется волокнами надресничного и ресничного сплетений, расположенных в области склеральной шпоры
В трабекулярной сети обнаруживаются как миелинизированные, так и немиелинизированные нервные волокна. Миелинизированные волокна образуют дугу, прилегающую к задней поверхности трабекулярного аппарата. Нервные окончания обильны, как в юкстаканаликулярной ткани, так и в трабекулярной сети (рис. 3.3.13).

Рис. 3.3.13. Распределение нервных окончаний (треугольники) в области трабекулярной сети и венозного склерального синуса (по Tripathi et al., 1982)
Ruskell нашел немиелинизированные волокна на всем протяжении трабекулярной сети и шлеммова канала. Наиболее часто они встречались в юкстаканаликулярной ткани, а также вблизи эндотелиальной выстилки шлеммова канала.
В настоящее время не совсем ясно, к какому типу относятся обнаруживаемые в этой области нервные волокна. Это во многом связано с тем, что нейротрансмиттерами являются многочисленные вещества. Нервные волокна могут быть аминэргическими, нитрэргическими и пептидэргическими. Nomura, Smelser, Ruskell считают, что симпатические волокна составляют 30%. Располагаются они в трабекулярной сети и передней части продольной ресничной мышцы и относятся к адренэргическим. Количество подобных волокон уменьшается с возрастом, а также при хронической простой глаукоме. Некоторые симпатические нервы иммунореактивны к нейропептиду Y.
Парасимпатическая иннервация угла передней камеры посредством волокон ресничного ганглия выявлена Holland, von Saliman. Collins. Ruskell установил, что у обезьян парасимпатические волокна поступают с лицевым нервом, образующих синапсы в крылонебном ганглии. В глазницу они поступают посредством rami orbitales.
Имеются данные, свидетельствующие о том, что нервы, исходящие из крылонебного ганглия, иммуноактивны в отношении вазоинтерстициального полипептида (VIP). Эти волокна также иннервируют заднюю часть увеального тракта глаза человека.
Чувствительные волокна тройничного нерва, содержащие Р вещество, выявлены в структурах угла глаза обезьян и человека, а также в увеальных и корнеосклеральных частях трабекулярной сети, юкстаканаликулярной ткани и шлеммовом канале. Иннервируются пептид-, нитр- и аминэргическими волокнами и миоэпителиальные клетки.
Отдельно необходимо остановиться на механорецепторах, обнаруживаемых в дренажной системе. Формируются они следующим образом. Внутренние слои глаза млекопитающих иннервируются сенсорными нервами, исходящими из тройничного нерва. Большинство волокон относится к волокнам типа С, а некоторые из них специфически окрашиваются на субстанцию Р.
Многочисленные ветви тройничного нерва проникают в склеру. При этом часть миелинизированных волокон образуют склеральное сплетение. Именно от этого сплетения отходят ветви к трабекулярной сети, теряя при этом миелиновую оболочку. Заканчиваются эти волокна нервными окончаниями типа механорецепторов. Рядом исследователей показано, что по строению механорецепторы трабекулярной сети наиболее близки к барорецепторам. Эти рецепторы специфически окрашиваются на наличие белков нейрофиламентов и синаптофизин, т. е. маркер синаптических пузырьков. Рецепторы трабекулярной сети подобны висцеральным механорецепторам других частей тела — каротидного синуса, дуги аорты, эндокарда, системы органов дыхания, пищевода, кожи, сухожилий.
Количество и плотность расположения механорецепторов трабекулярной сети увеличиваются с возрастом, а также при хронической простой глаукоме.
Существует три гипотезы, объясняющие роль механорецепторов, расположенных в области дренажной системы. Они могут выполнять функцию проприоцепции сухожилий ресничной мышцы, влиять на сокращение миофибробластоподобных клеток склеральной шпоры. Кроме того, они могут функционировать как барорецепторы при изменении внутриглазного давления.
↑ Увеосклеральный путь оттока
Передняя часть ресничного тела, увеосклеральная часть трабекулярного аппарата, передняя поверхность радужки являются потенциальными местами распространения камерной влаги в супрахориоидею, что и было показано многими исследователями. После проникновения камерной влаги в строму перечисленных структур она поступает в супрахориоидею, а затем распространяется через склеру в сосудистую систему, включая вортикозные вены. Предполагают, что около 10% объема оттока камерной влаги происходит именно этим путем.
↑ Регуляция внутриглазного давления
Механизмы регуляции внутриглазного давления до сих пор не совсем ясны. Очевидно только, что как секреция камерной влаги, так w сопротивление ее оттоку регулируются. Повышение внутриглазного давления, в конечном итоге, тормозит секрецию камерной влаги. Кроме того, повышение внутриглазного давления должно сопровождаться «раскрытием» пустей оттока, т. е. увеличивать количество трансцеллюлярных каналов, расположенных во внутренней стенке шлеммова канала.
Хотя холинэргические препараты (типа пилокарпина) и препараты с а-адренэргическим действием (типа адреналина) лишь незначительно понижают давление в норме, степень снижения давления при открытоугольной глаукоме значительно выше, что используется в ее лечении. В нормальном глазу влияние этих препаратов на давление контролируется гомеостатическими регулирующими механизмами: оба препарата снижают сопротивляемость дренажных структур оттоку. Пилокарпин увеличивает количество трансцеллюлярных пор в эндотелиальной выстилке шлеммова канала. Препарат также действует на ресничную мышцу, которая посредством своих сухожилий прикрепляется к склеральной шпоре и увеосклеральной части трабекулярного аппарата. Этот механизм был показан многими исследователями. Каким образом сокращение ресничной мышцы приводит к усилению оттока камерной влаги? Rohen показал, что сухожилия ресничных мышц присоединяются к волокнам трабекулярной сети. Выделяют три типа сухожилий.
- Первый тип сухожилий исходит из наиболее отдаленных пучков продольного слоя ресничной мышцы и прикрепляется к склере или склеральной шпоре.
- Второй тип сухожилий передает нагрузку от склеральной шпоры волокнам, расположенным в трабекулярной сети. Они состоят из эластоподобных волокон, которые распределяются в наружной части трабекулярной сети и соединяются с волокнами, лежащими под эндотелием.
- Третий тип волокон — коллагеновые. Они образуют широкие длинные полосы, проходящие через трабекулярную сеть и прикрепляющиеся к строме роговицы. Подобное прикрепление сухожилий при сокращении ресничной мышцы разворачивает трабекулы так, что межтрабекулярные пространства увеличиваются. Расширяется и просвет шлеммова канала, что сопровождается увеличением площади фильтрации жидкости и, естественно, снижением сопротивления оттоку. Описанный механизм роли ресничной мышцы подтвержден в экспериментальных исследованиях.
Каким образом реализуется влияние адреналина, остается неясным, хотя предполагают, что он действует непосредственно на трабекулярные клетки и на некоторые сосуды, обеспечивающие дренаж камерной влаги на уровне коллекторных сосудов. Адренэргические бета-блокаторы и ингибиторы карбонагидразы, используемые в лечении глаукомы, уменьшают скорость секреции ресничным телом камерной влаги ЭТ0.
В последние годы благодаря разработке новых методов анализа (иммуноморфология, методы молекулярной генетики) проводятся интенсивные исследования механизмов регуляции внутриглазного давления. Особое внимание при этом уделяется выявлению роли щелевых контактов между трабекулярными клетками и микротрубочками, роли биологически активных веществ, состава и состояния межклеточного вещества, особенно юкстаканаликулярной ткани. Особое внимание уделяется изменению объема трабекулярных клеток в результате изменения ионного состава клеток, с чем связывают регуляцию оттока камерной влаги. Участие в регуляции внутриглазного давления принимают такие биологические активные вещества, как интерлейкин-6, препротахикинин-1, секретогранин-П, катепсин-L, стромелизин-1, тимозин, тубулин, fi-кристалин, глицеральде-гид-3-фосфатдегидрогеназа и Cu/Zn дисмутаза перекиси водорода, миоцилин, простагландины и др. Изменение эспрессии перечисленных метаболитов обнаруживается в эспериментальных условиях изменения внутриглазного давления. Столь интенсивные исследования биохимических и физиологических механизмов регуляции внутриглазного давления связаны с практической необходимостью создания новых лекарственных средств в лечении глаукомы. Пока эти исследования находятся в стадии накопления данных.
↑ Старение глаза и открытоугольная глаукома
С возрастом развиваются структурные изменения дренажной системы, увеличивающие сопротивление оттоку камерной влаги и способствующие развитию глаукомного процесса. Степень структурных изменений дренажной системы коррелирует со степенью изменений сосудов организма при общих сосудистых заболеваниях. Это отмечено при атеросклерозе, гипертонической болезни, сахарном диабете и др.
При старении в два-три раза утолщаются трабекулы, главным образом в результате накопления спиралевидного коллагена. Увеличивается количество базального материала. Однако количество протеогликанов (хондроитинсульфат) уменьшается. Исчезает микрофибриллярный компонент эластических волокон.
Показано, что в процессе физиологического старения в дренажной зоне глаза происходят незначительные нарушения в виде мукоидного набухания. Эти изменения могут привести к нарушению оттока камерной влаги, но глаукома не развивается, так как гомеостатические механизмы, обеспечивающие поддержание внутриглазного давления на физиологическом уровне, компенсируют этот сдвиг.
Рядом авторов выявлены дегенеративные изменения трабекулярных клеток, число которых прогрессивно снижается. Слой клеток истончается, трабекулы «сливаются». Этот процесс расценивают как «гиалиноз» трабекулярного аппарата, который приводит к увеличению сопротивляемости оттоку камерной влаги и повышению внутриглазного давления. Отмечается и уменьшение числа клеток в юкстаканаликулярной ткани. В ней накапливается материал, являющийся продуктом распада эластических волокон и других молекул типа спиралевидного коллагена. Содержание в этой области гиалуроновой кислоты с возрастом также снижается. Биохимическими исследованиями показано увеличение количества фибронектина, коллагена VI типа и тромбоспондина. При этом уменьшается количество ламинина, который, тем не менее, в повышенном количестве обнаружен под эндотелиальной выстилкой шлеммова канала.
Приведенные выше изменения выявлены и при развитии первичной открытоугольной глаукомы.
Трабекулярная сеть при глаукоме. Наиболее ранние изменения трабекулярного аппарата при открытоугольной глаукоме пока не установлены. Исследование участков трабекулярного аппарата, удаленного во время операции на поздних стадиях глаукомы, позволило Rohen и Witmer выявить материал в виде «бляшки», располагающийся в сетчатой части трабекулярной сети и под эндотелиальными клетками шлеммова канала. Они различают 3 типа «бляшек».
- Первый тип «бляшек» преимущественно располагается у шлеммова канала и состоит из гомогенного или мелкозернистого материала.
- «Бляшки» второго типа выглядят на тангенциальных срезах в виде точек. При электронномикроскопическом исследовании они представляют собой центральные участки эластоподобных волокон, разрезанных поперек.
- «Бляшки» третьего типа состоят из электронноплотного материала, содержащего зернистый компонент и исчерченные фибриллы.
Количество всех трех типов «бляшкоподобного» материала увеличивается с возрастом, а при открытоугольной глаукоме количество этого материала значительно больше независимо от возраста больного. Накопление «бляшкоподобного» материала может являться препятствием на пути оттока камерной влаги, особенно при локализации его вблизи эндотелиальных клеток шлеммова канала.
Микроскопически также выявлено, что при открытоугольной глаукоме возможно спадение наружной и внутренней стенок шлеммова канала. При этом отсутствует эндотелиальная выстилка. Сочетались эти изменения со значительным скоплением «бляшкоподобного» материала.
При открытоугольной глаукоме нередко обнаруживаются также признаки воспаления, проявляющиеся инфильтрацией трабекулярной сети лимфоцитами. Подобная инфильтрация выявляется только на поздних стадиях развития заболевания.
При глаукоме уменьшается также количество трабекулярных клеток. Уменьшение количества трабекулярных клеток сопровождается появлением в сохранившихся клетках так называемых матричных пузырьков, представляющих собой морофлогическую форму лизосом. Прогрессивное уменьшение количества трабекулярных клеток может стать причиной «слипания» трабекул между собой.
Нередко при глаукоме наступает гиалиноз корнеосклеральных и увеальных трабекул. Этому, как правило, предшествует накопление базальноподобного материала. Подобные изменения довольно сильно напоминают возрастные, что ряду авторов дает основание предполагать наличие единых механизмов, лежащих в основе старения и возникновения открытоугольной глаукомы. Подтверждении тому являются данные о нарушении процессов перекисного окисления, как при старении, так и при глаукоме. Показано первичное повреждение продуктами перекисного окисления клеточных мембран эндотелиальных клеток, что может явиться пусковым механизмом развития сосудистых заболеваний глаза и глаукомы.
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0