Функциональная анатомия зрительной системы │ Часть 3


Содержание:
Описание
↑ Наружное коленчатое тело
Наружное коленчатое тело (corpus geniculatum laterale) является местом расположения так называемого «второго нейрона» зрительного пути. Через наружное коленчатое тело проходит около 70% волокон зрительного тракта. Наружное коленчатое тело представляет собой возвышенность, соответствующую месту расположения одного из ядер зрительного бугра (рис. 4.2.26—4.2.28).
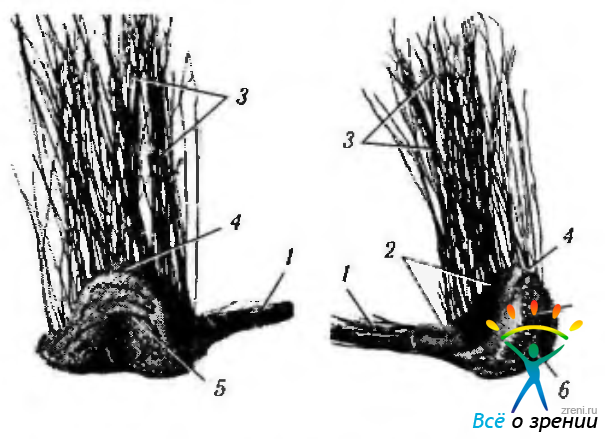
Рис. 4.2.26. Модель левого наружного коленчатого тела (по Wolff, 1951): а — вид сзади и изнутри; б — вид сзади и снаружи (1 — зрительный тракт; 2 — седло; 3 — зрительная лучистость; 4 — головка; 5 - тело; 6 - перешеек)
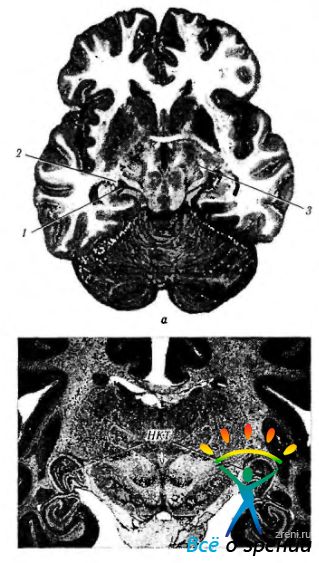
Рис. 4.2.27. Наружное коленчатое тело и его отношение к структурам головного мозга: а — горизонтальный срез мозга (1 — наружное коленчатое тело; 2— внутренняя капсула; 3—подушка зрительного бугра); б — сагиттальный срез мозга (гистологический срез, окрашенный гематоксилином и эозином) (НКТ—наружное коленчатое тело)
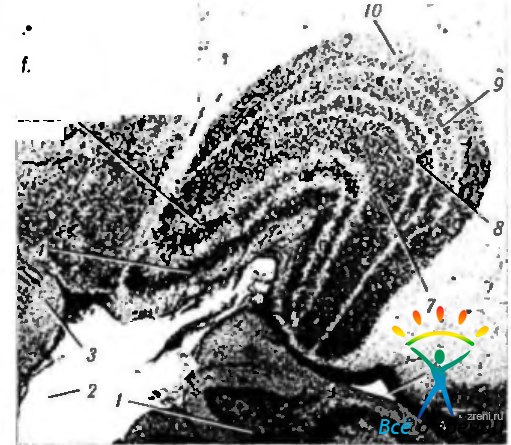
Рис. 4.2.28. Наружное коленчатое тело: 1 — гиппокамп; 2— субарохноидальное пространство; 3 — ножка мозга; 4 — слой 1; 5 — слой 2; б — нижний рог бокового желудочка; 7 — слой 3; 8 — слой 4; 9 — слой 5; 10 — слой 6. Наружное коленчатое тело является ядром зрительного бугра. Четко видно наличие шести темных слоев скопления нейронов, разделенных светлыми слоями, состоящими из нервных волокон. Слои 1 и 2 складываются из крупных нейронов (магноцеллюлярные), а слои 3—6 - из мелких клеток (парвоцеллюлярные)
Содержит оно около 1 800 000 нейронов, на дендритах которых заканчиваются аксоны ганглиозных клеток сетчатой оболочки.
Ранее предполагали, что наружное коленчатое тело представляет собой лишь «ретрансляционную станцию», передающую информацию от нейронов сетчатки через зрительную лучистость коре головного мозга. В настоящее время показано, что на уровне наружного коленчатого тела происходит довольно существенная и разноплановая обработка зрительной информации. О нейрофизиологическом значении этого образования речь пойдет несколько ниже. Первоначально необходимо остановиться на его анатомических особенностях.
Ядро наружного коленчатого тела представляет собой одно из ядер зрительного бугра. Располагается оно между вентропостериолатеральным ядром зрительного бугра и подушкой зрительного бугра (рис. 4.2.27).
Наружное коленчатое ядро состоит из дорзального и филогенетически более древнего вентрального ядер. Вентральное ядро у человека сохранено в виде рудимента и состоит из группы нейронов, расположенных ростральней дорзального ядра. У низших млекопитающих это ядро обеспечивает наиболее примитивные фотостатические реакции. Волокна зрительного тракта к этому ядру не подходят.
Дорзальное ядро составляет основную часть ядра наружного коленчатого тела. Представляет оно собой многослойную структуру в виде седла или асимметричного конуса с округленной верхушкой (рис. 4.2.26—4.2.28). На горизонтальном срезе видно, что наружное коленчатое тело связано спереди со зрительным трактом, с латеральной стороны — с ретролентикулярной частью внутренней капсулы, медиально — со средним коленчатым телом, сзади с гиппокампальной извилиной, а постериолатерально — с нижним рогом бокового желудочка. К ядру наружного коленчатого тела сверху прилежит подушка зрительного бугра, антериолатерально — темпоропонтинные волокна и задняя часть внутренней капсулы, латерально — зона Вернике, а с внутренней стороны — медиальное ядро (рис. 4.2.27). Зона Вернике является самой внутренней частью внутренней капсулы. Именно в ней и начинается зрительная лучистость. Волокна зрительной лучистости располагаются с дорзолатеральной стороны ядра наружного коленчатого тела, в то время как волокна слухового тракта — с дорзомедиальной.
Наружное коленчатое тело соединяется с верхним четверохолмием при помощи связки, называемой передним плечом.
Даже при макроскопическом исследовании наружного коленчатого тела выявляется, что это образование обладает слоистым строением. У обезьян и человека четко различается шесть полос «серого вещества» и расположенные между ними «белые» прослойки, состоящие из аксонов и дендритов (рис. 4.2.28). Первым слоем обозначен слой, расположенный с вентральной стороны. Два внутренних слоя состоят из клеток большого размера (магноцеллюлярные слои 1 и 2). Получили они такое название по той причине, что состоят из крупных нейронов с эксцентрично расположенным ядром и большим количеством в цитоплазме вещества Ниссля. Аксоны нейронов магноцеллюлярного слоя формируют не только зрительную лучистость, но также направляются к верхним бугоркам четверохолмия. Четыре наружных слоя состоят из маленьких и среднего размера клеток (парвоцеллюлярные слои, 3—6). Они содержат нейроны, получающие информацию от сетчатки и передающие ее только зрительной коре головного мозга (формируют зрительную лучистость). Обнаруживаются и нейроны, обеспечивающие связь между нейронами наружного коленчатого тела. Это так называемые «вставочные нейроны» (интернейроны). Предполагают, что два слоя, состоящие из мелких нейронов (парвоцеллюлярные слои), появляются в связи с развитием центрального зрения.
Важно отметить, что на перечисленные слои нейронов проецируются волокна, идущие от различных участков сетчатки обоих глаз. Так, перекрещенные волокна зрительного тракта заканчиваются в 1, 4 и 6-м слоях, а неперекрещенные — во 2, 3 и 5-м (рис. 4.2.29).
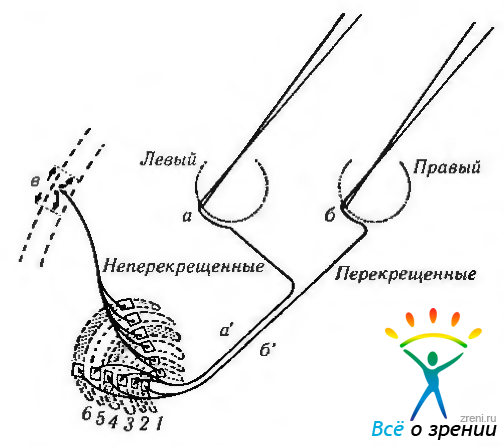
Рис. 4.2.29. Представительство сетчатой оболочки в наружном коленчатом теле: Импульсы от корреспондирующих точек (а, б) двух сетчаток проходят в зрительный тракт. Неперекрещенные волокна (а') заканчиваются во 2, 3 и 5-м слоях наружного коленчатого тела. Перекрещенные волокна (б') заканчиваются в слоях 1, 4 и 6. Импульсы после прохождения НКТ (в') проецируются на кору головного мозга
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Это происходит таким образом, что волокна от корреспондирующих частей двух половин сетчатки (например, правая височная и левая назальная половины сетчатки) заканчиваются в соседних слоях. Приведенные особенности проекции на наружное коленчатое тело установлены на основании использования разнообразных методов исследования. Так, в случаях разрушения контрлатерального зрительного нерва или предшествовавшего удаления глазного яблока развивается дегенерация нейронов 1, 4 и 6-го слоев наружного коленчатого тела (рис. 4.2.30).
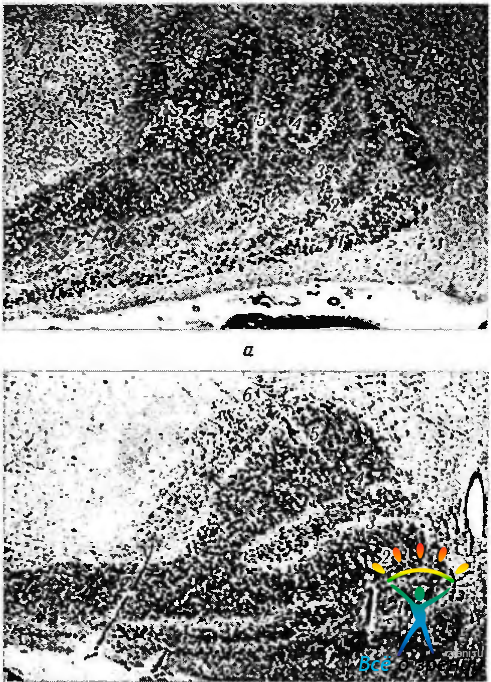
Рис. 4.2.30. Изменение микроскопического строения наружного коленчатого тела с двух сторон при удалении одного глазного яблока (no Alvord, Spence, 1997): а — наружное коленчатое тело (НКТ), расположенное ипсилате-рально относительно энуклеированного глаза; б — НКТ, расположенное контрлатерально относительно энуклеированного глаза (После смерти больного, у которого задолго до смерти было удалено глазное яблоко, микроскопически исследованы наружные коленчатые тела. После нарушения нормальной проекции ганглиозных клеток сетчатой оболочки на нейроны НКТ наступает атрофия последних. При этом интенсивность окрашивания слоев уменьшается. На рисунке видно, что 3-й и 5-й слои НКТ, расположенные ипсилатерально относительно удаленного глаза, значительно слабее окрашены гематоксилином и эозином. В то же время слои 3 и 5 НКТ, расположенные контрлатерально относительно удаленного глаза, окрашены более интенсивно, чем слои 4 и 6. Можно также отметить, что слои 1 и 2 поражены в наименьшей степени)
При разрушении гомолатеральных волокон зрительного нерва наступает дегенерация нейронов 2, 3 и 5-го слоев. Это явление называется транссинаптической дегенерацией. Установлено также, что если при рождении котенку сшить веки одного глаза, то через три месяца наступит дегенерация 25—40% нейронов наружного коленчатого тела. Подобной формой транссинаптической дегенерации можно объяснить некоторые механизмы развития амблиопии, развивающейся при врожденном косоглазии.
О различной проекции на наружное коленчатое тело перекрещенных и неперекрещенных волокон свидетельствуют и экспериментальные исследования. В этих исследованиях в одно из глазных яблок вводится радиоактивная аминокислота, распространяющаяся трансаксонально по направлению наружного коленчатого тела и накапливающаяся в его нейронах (рис. 4.2.31).
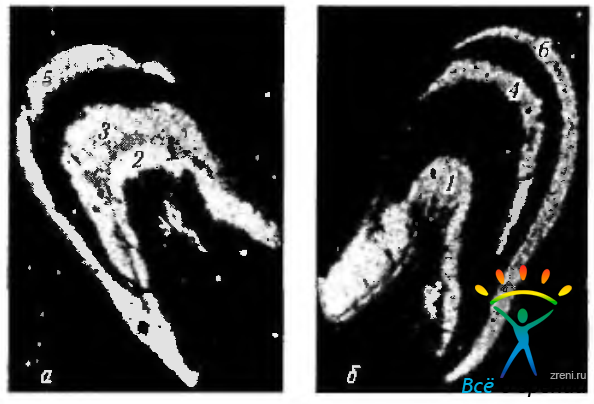
Рис. 4.2.31. Распределение радиоактивной метки в наружных коленчатых телах после введения в левое глазное яблоко обезьяны радиоактивной аминокислоты: а - левое наружное коленчатое тело; б - правое наружное коленчатое тело. (Аминокислота поглощается ганглиозными клетками сетчатой оболочки и транспортируется по аксонам через зрительный нерв, зрительный перекрест и зрительный тракт к наружному коленчатому телу. Иллюстрация указывает на то, что слои 2, 3 и 5 получают информацию от ипсилатерального глаза, а слои 1, 4 и 6 — от контрлатерального глаза)
Особенности проекции сетчатки на наружное коленчатое тело. В последнее время выявлены особенности проекции сетчатой оболочки на наружное коленчатое тело. Сводятся они к тому, что каждая точка половины сетчатки точно проецируется на определенную точку ядра наружного коленчатого тела («точка к точке»). Таким образом, пространственное возбуждение в слое ганглиозных клеток сетчатки «картируется» пространственным распределением возбуждения нейронов в разных слоях наружного коленчатого тела. Строгий топографический порядок связей наблюдается и между клетками различных слоев. Проекции каждой точки поля зрения во всех слоях находятся непосредственно одна под другой, так что можно выделить колонкообразный участок, пересекающий все слои наружного коленчатого тела и соответствующий проекции локальной области поля зрения.
Приведенная закономерность проекции выявлена на основании экспериментальных исследований. Так, показано, что локальное точечное повреждение сетчатки приводит к развитию транснейронной дегенерации небольших, но четко очерченных скоплений клеток в трех слоях наружного коленчатого тела с обеих сторон. Фокальное повреждение зрительной коры или введение в нее радиоактивного трейсера приводит к «маркировке» клеток или волокон, расположенных на линии, простирающейся поперек всех слоев наружного коленчатого тела на том же самом уровне. Эти участки соответствуют «рецептивным полям» наружного коленчатого тела и получили название «проекционная колонка».
В этом месте изложения материала целесообразно остановиться на особенностях рецептивных полей наружного коленчатого тела. Рецептивные поля наружного коленчатого тела напоминают таковые ганглозных клеток сетчатой оболочки. Различают несколько основных типов рецептивных полей. Первый тип характеризуется наличием ON-ответа при возбуждении центра и OFF-ответа при возбуждении периферии (ON/OFF-тип). Второй тип рецептивных полей характеризуется обратным отношением — OFF/ON-тип. ДЛЯ наружного коленчатого тела характерно также и то, что в слоях 1 и 2 обнаруживается смесь рецептивных полей первого и второго типов. В то же время в слоях 3—6 обнаруживается только один тип рецептивных полей (в двух слоях поля первого типа, а в других двух — второго типа). Обнаруживаются также линейные рецептивные поля с различным соотношением ON- и OFF-центров. Использование электрофизиологических методов позволило выявить, что рецептивные поля наружного коленчатого тела обладают более выраженной оппонентной реакцией, чем рецептивные поля ганглиозных клеток сетчатки. Именно это предопределяет большое значение наружного коленчатого тела в усилении контраста. Выявлены также явления пространственно-временной суммации поступающих сигналов, анализа спектральных характеристик сигнала и т. д. Нейроны наружного коленчатого тела, участвующие в кодировании цвета, локализуются в парвоцеллюлярных слоях, где сконцентрированы цветооппонентные клетки «красно-зеленого» и «сине-желтого» цвета. Как и для ганглиозных клеток сетчатки, для них характерна линейная суммация колбочковых сигналов по площади сетчатки. Магноцеллюлярные слои также состоят из оппонентных нейронов с пространственно распределенными в рецептивных полях входами от колбочек разного типа. Необходимо отметить, что анатомическая сегрегация нейронов с различными функциональными свойствами наблюдается уже в сетчатке, где отростки биполяров и ганглиозных клеток ON- и OFF-типов локализуются в разных подслоях внутреннего плексиформного слоя. Такое «анатомическое обособление» нейронных систем, образующих разные каналы передачи информации, является общим принципом в построении анализаторных структур и наиболее выражено в колончатой структуре коры, на чем мы остановимся несколько ниже.
Продолжая описание особенностей проекции сетчатки на наружное коленчатое тело, необходимо отметить, что периферические височные участки сетчатки противоположного глаза проецируются на слои 2, 3 и 5 и называются монокулярным полумесяцем.
Наиболее полные данные о ретинотопической организации волокон зрительного нерва, зрительного перекреста и ядер наружного коленчатого тела у человека и обезьяны получены Brouewer, Zeeman, Polyak, Hoyt, Luis. Первоначально мы опишем проекцию нeмакулярных волокон. Неперекрещивающиеся волокна, идущие от верхне-височного квадранта сетчатки, в зрительном перекресте располагаются дорзо-медиально и проецируются на медиальую часть ядра наружного коленчатого тела. Неперекрещивающиеся волокна, идущие от нижне-височного квадранта сетчатки, в зрительном перекресте располагаются снизу и латерально. Они проецируются на наружную часть наружного коленчатого тела (рис. 4.2.29). Макулярная область сетчатки проецируется на клиновидный сектор, расположенный в задних двух третях или трех четвертях наружного коленчатого тела (рис. 4.2.34, 4.2.35).
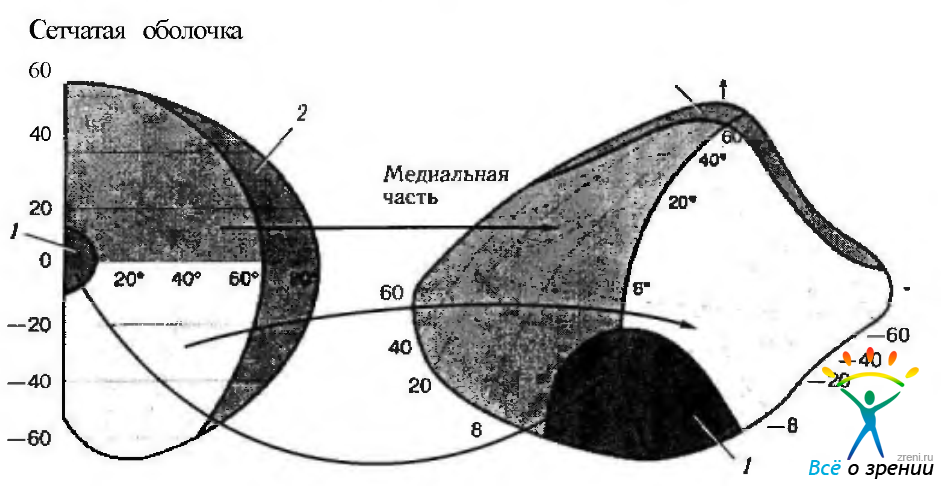
Рис. 4.2.34. Проекция сетчатой оболочки на наружное коленчатое тело: 1 - макула; 2 - монокулярный полумесяц
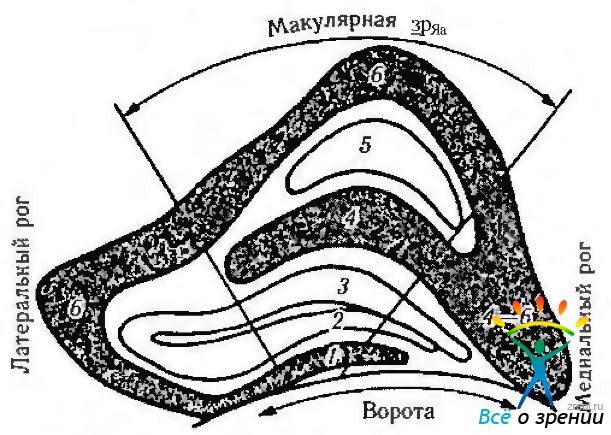
Рис. 4.2.35. Схематическое изображение коронарного среза через наружное коленчатое тело (вид сзади) (по Miller, 1985): обращает на себя внимание большое представительство в наружном коленчатом теле макулярной области (1—6—номера слоев НКТ)
Отмечено, что представительство зрительных полуполей в зрительном тракте как бы «поворачивается» на уровне наружного коленчатого тела таким образом, что вертикальное сечение становится горизонтальным. При этом верхняя часть сетчатки проецируется на медиальную часть, а нижняя на латеральную часть наружного коленчатого тела. Этот поворот полностью изменяется в зрительной лучистости таким образом, что, когда волокна достигают зрительной коры, квадрант верхней части сетчатки располагается в верхней части тракта, а нижний квадрант — снизу.
Синаптические взаимодействия нейронов наружного коленчатого тела. Ранее предполагали, что аксон ганглиозной клетки контактирует только с одним нейроном наружного коленчатого тела. Благодаря электронной микроскопии установлено, что афферентные волокна образуют синапсы с несколькими нейронами, (рис. 4.2.36).
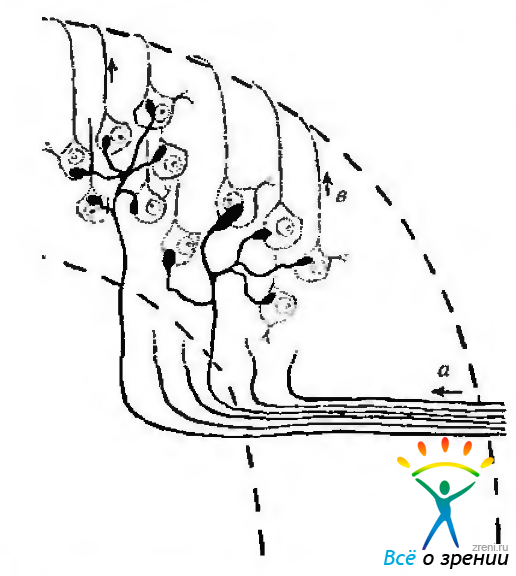
Рис. 4.2.36. Схематическое изображение взаимодействия терминалов аксонов ганглиозных клеток сетчатки с нейронами наружного коленчатого тела у обезьяны (по Glees, Le Gros, Clark, 1941): пучок волокон зрительного нерва (а) входит в клеточный слой (б) наружного коленчатого тела (НКТ) справа. Некоторые волокна отдают 5—6 ветвей, подходят к телу нейронов НКТ и образуют синапс. Аксоны клеток НКТ (в) покидают клеточный слой НКТ, проходят через волокнистый слой и формируют зрительную лучистость
В то же время каждый нейрон наружного коленчатого тела получает информацию от нескольких ганглиозных клеток сетчатки. На основании ультраструктурных исследований выявлены также разнообразные синаптические контакты между ними. Аксоны ганглиозных клеток могут заканчиваться как на теле нейронов наружного коленчатого тела, так и на первичных или вторичных их дендритах. При этом формируются так называемые «клубочковые» окончания (рис. 4.2.37).
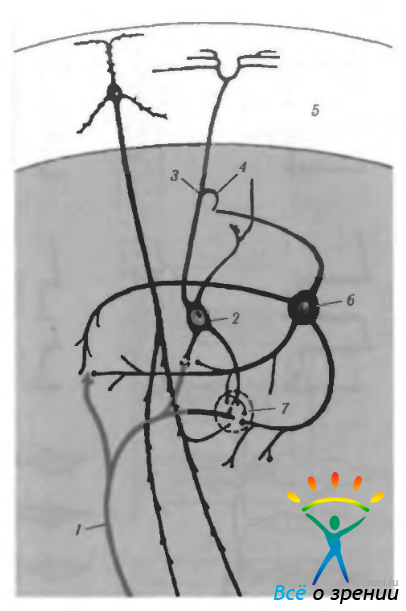
Рис. 4.2.37. Схематическое изображение нейронных связей гломерулы наружного коленчатого тела (НКТ) (по Carpenter, 1976): 1 — первичный афферент сетчатки; 2 — нейрон наружного коленчатого тела; 3 — аксон; 4 — коллатераль аксона; 5 — зрительная кора; б — клетка Гольджи II типа; 7 — гломерула. Первичные афференты сетчатки окрашены в красный цвет, а нейроны НКТ — в голубой. Кортикофугальные волокна и клетки Гольджи II типа (интернейроны) окрашены в черный цвет. Гломерула состоит из терминала первичного афферента сетчатки, нескольких терминалов нейрона НКТ, кортикофутального волокна и дендрита клетки Гольджи II типа. Этот комплекс окружен глиальной капсулой
У кошек «клубочки» отделены от окружающих образований тонкой капсулой, состоящей из отростков глиальных клеток. Подобная изоляция «клубочков» отсутствует у обезьян.
Синаптические «клубочки» содержат синапсы аксонов ганглиозных клеток сетчатки, синапсы нейронов наружного коленчатого тела и вставочных нейронов («интернейронов»). Эти синаптические образования напоминают «триады» сетчатой оболочки.
Каждый «клубочек» состоит из зоны плотно упакованных нейронов и их терминалов. В центре этой зоны расположен аксон ганглиозной клетки сетчатки, являющийся пресинаптическим. Он образует синапсы с нейроном наружного коленчатого тела и вставочными нейронами. Дендриты нейронов наружного коленчатого тела поступают в «клубочки» в виде шипа, который непосредственно и образует синапс с аксоном сетчатки. Дендрит вставочных нейронов (интернейронов) образует синапс со смежным «клубочком», формируя между ними последовательные синапсы.
Lieberman выделяет пре- и постсинаптические «ингибирующие» и «возбуждающие» дендритические и «клубочковые» синапсы. Они представляют собой сложное скопление синапсов между аксонами и дендритами. Именно эти синапсы структурно обеспечивают феномен торможения и возбуждения рецептивных полей наружного коленчатого тела.
Функции наружного коленчатого тела. Предполагают, что к функциям наружного коленчатого тела относятся: усиление контраста изображения, организация зрительной информации (цвет, движение, форма), модуляция уровня обработки зрительной информации с их активацией (посредством ретикулярной формации). Обладает наружное коленчатое тело и бинокулярными рецептивными полями. Важно отметить, что на функции наружного коленчатого тела влияют и более высоко расположенные центры мозга. Подтверждением роли наружного коленчатого тела в обработке информации, идущей от высших отделов мозга, является обнаружение проекции на него эфферентных волокон, исходящих из коры головного мозга. Возникают они в VI слое зрительной коры и проецируются на все слои наружного коленчатого тела. По этой причине незначительное повреждение зрительной коры вызывает атрофию нейронов во всех шести слоях наружного коленчатого тела. Терминалы этих волокон небольшого размера и содержат многочисленные синаптические пузырьки. Оканчиваются они как на дендритах нейронов наружного коленчатого тела, так и на вставочных нейронах («интернейронах»). Предполагают, что посредством этих волокон кора головного мозга модулирует деятельность наружного коленчатого тела. С другой стороны, показано, что изменение активности нейронов наружного коленчатого тела избирательно активизируют или тормозят нейроны зрительной коры мозга.
Существуют и другие связи наружного коленчатого ядра. Это связь с подушкой зрительного бугра, вентральным и латеральным ядрами зрительного бугра.
Кровоснабжение наружного коленчатого тела осуществляется задней мозговой и задней ворсинчатой артериями. Основным сосудом, кровоснабжающим наружное коленчатое тело, особенно задне-внутреннюю его поверхность, является задняя мозговая артерия. В ряде случаев от этой артерии отходит ветвь — задняя ворсинчатая (хориоидальная) артерия. При нарушении кровообращения в этой артерии обнаруживаются нарушения поля верхнего гомонимного квадранта сетчатой оболочки.
Передняя ворсинчатая (хориоидальная) артерия почти полностью кровоснабжает переднюю и боковую поверхности наружного коленчатого тела. По этой причине нарушение кровообращения В ней приводит К поражению волокон. исходящих из нижнего квадранта сетчатки. Эта артерия отходит от внутренней сонной артерии (иногда от средней мозговой артерии) сразу дистальней места выхода задней соединительной артерии. При достижении передней части наружного коленчатого тела передняя ворсинчатая артерия отдает различное число ветвей перед вступлением в нижний рог бокового желудочка.
Часть наружного коленчатого тела, на которую проецируются волокна, исходящие из желтого пятна, кровоснабжается как передней, так и задней ворсинчатой артериями. Кроме того, от хорошо развитой системы анастомозов, расположенных в мягкой и паутинной оболочках мозга, отходят многочисленные артериолы, проникающие в наружное коленчатое тело. Там они образуют густую сеть капилляров во всех его слоях.
↑ Зрительная лучистость
Зрительная лучистость (radiatio optica; Грасиоле, Gratiolet) является аналогом других лучистостей зрительного бугра, таких как слуховая, затылочная, теменная и лобная. Все перечисленные лучистости проходят через внутреннюю капсулу, соединяющую полушария мозга и стволовую часть мозга, спинной мозг. Внутренняя капсула находится латеральней зрительного бугра и боковых желудочков мозга и медиальней чечевицеподобного ядра (рис. 4.2.40, 4.2.41).

Рис. 4.2.40. Зрительная лучистость. Боковой вид мозга после удаления части левого полушария: 1 — центральная извилина; 2 — средняя лобная извилина; 3 — прецентральная извилина; 4 нижняя лобная извилина; 5—дугообразный пучок; 6 — зрительная лучистость; 7 — затылочная доля; 8 — мозжечок; 9 — височная доля
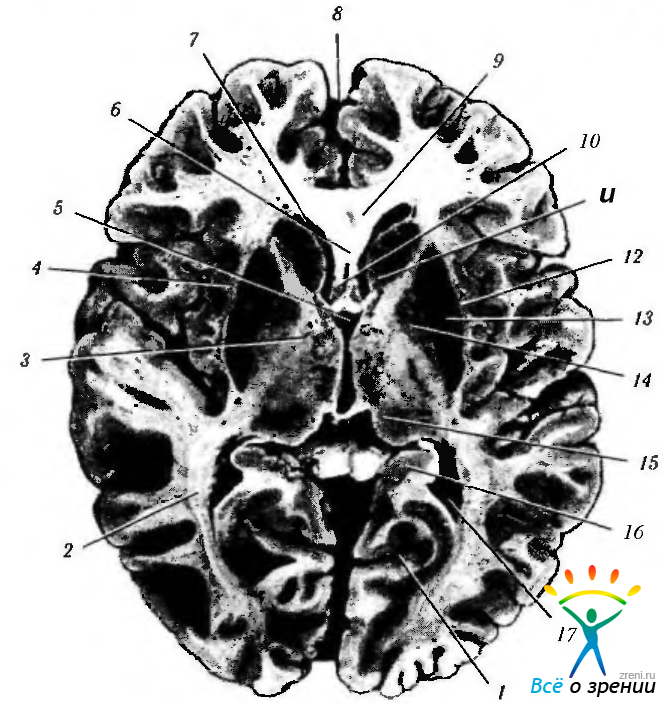
Рис. 4.2.41. Горизонтальный срез мозга на уровне расположения зрительной лучистости: 1 — шпорная борозда; 2 — зрительная лучистость; 3 — внутренняя капсула; 4 — наружная капсула; 5 — четвертый желудочек; 6 — пластинка прозрачной перегородки; 7 — передний рог бокового желудочка; 8 — продольная щель мозга; 9 — колено мозолистого тела; 10 полость прозрачной перегородки; 11 — головка хвостатого ядра; 12 — ограда; 13 — скорлупа; 14 — бледный шар; 15 — зрительный бугор; 16 — гиппокамп; 17 — заднее колено бокового желудочка
Наиболее задняя часть внутренней капсулы содержит волокна слуховой и зрительной лучистости и нисходящие волокна, идущие от затылочной коры к верхним бугоркам четверохолмия.
Зрительная лучистость соединяет наружное коленчатое тело с корой затылочной доли головного мозга. При этом ход волокон, исходящих от различных отделов наружного коленчатого тела, довольно существенно отличается. Так, волокна, идущие от нейронов латерального отдела наружного коленчатого тела, огибают нижний рог бокового желудочка, расположенный в височной доле, а затем, направляясь кзади, проходят под задним рогом этого желудочка, достигая нижних отделов зрительной коры, вблизи шпорной борозды (рис. 4.2.40, 4.2.41). Волокна от медиального отдела наружного коленчатого тела идут несколько более прямым путем к первичной зрительной коре (поле 17 по Бродману), расположенной в медиальной части затылочной доли. Волокна этого пути отклоняются латерально, пройдя непосредственно кпереди от входа в боковой желудочек, а затем поворачивают кзади, идут в каудальном направлении, огибая сверху задний рог этого желудочка и оканчиваются в коре, расположенной вдоль верхнего края шпорной борозды.
Верхние волокна, покидающие наружное коленчатое тело, направляются прямо к зрительной коре. Нижние волокна делают петлю вокруг желудочков мозга (петля Меера) и направляются к височной доле. Нижние волокна плотно прилежат к чувствительным и двигательным волокнам внутренней капсулы. Даже небольшой инсульт, возникающий в этой области, приводит к верхним гемианопсическим дефектам поля зрения и гемипарезу (контрлатеральному) .
Наиболее передние волокна обнаруживаются приблизительно в 5 см позади вершины височной доли. Отмечено, что лобэктомия, при которой мозговую ткань иссекают в 4 см от вершины височной доли, не приводит к появлению дефекта поля зрения. При повреждении более обширной области (глубоко расположенные опухоли, темпоральная декомпрессия по поводу травмы или инфекционного заболевания) развиваются гомонимные верхнеквадрантные гемианопсии. Наиболее типичные формы дефекта поля зрения при повреждении зрительной лучистости приведены на рис. 4.2.43.

Рис. 4.2.43. Схема распространения волокон в зрительном тракте, наружном коленчатом теле и зрительной лучистости. Нарушение поля зрения при повреждении участков, расположенных после зрительного перекреста: 1 — сдавление зрительного тракта — гомонимная гемианопсия с нечетким краем; 2 — сдавление проксимальной части зрительного тракта, наружного коленчатого тела или нижней части зрительной лучистости — гомонимная гемианопсия без сохранения макулярного поля с четким краем; 3 — сдавление передней петли зрительной лучистости — верхняя квадрантная анопсия с нечеткими краями; 4—сдавление верхней части зрительной лучистости — нижняя квадрантная анопсия с нечеткими краями; 5 — сдавление средней части зрительной лучистости — гомоним ная гемианопсия с нечеткими краями и выпадением центрально го зрения; 6 — сдавление задней части зрительной лучистости — конгруентная гомонимная гемианопсия с сохранением центрального зрения; 7 — сдавление передней части коры в области шпоры — темпоральное выпадение поля зрения с противоположной стороны; 8 — сдавление средней части коры в области шпоры — гомонимная гемианопсия с сохранением центрального зрения со стороны поражения и сохранением темпорального поля зрения с противоположной стороны; 9 — сдавление задней части коры за тылочной области — конгруентная гомонимная гемианопсическая скотома
Как указано выше, зрительная лучистость содержит 3 главные группы волокон. Верхняя часть содержит волокна, обслуживающие нижние поля зрения, нижняя часть — верхние поля. Центральная часть содержит макулярные волокна.
Ретинотопическая организация волокон наружного коленчатого тела распространяется и на зрительную лучистость, но с некоторыми изменениями в положении волокон (рис. 4.2.42).

Рис. 4.2.42. Топографические особенности проекции различных отделов сетчатой оболочки на разных уровнях зрительного пути: 1 — макулярная область; 2 — верхний наружный квадрант сетчатки; 3 — нижний наружный квадрант сетчатки; 4 — «монокулярный полумесяц»
Дорзальный пучок волокон, представляющий верхний периферический квадрант сетчатки, исходит из медиальной части наружного коленчатого тела и проходит к дорзальной губе птичьей шпоры. Вентральный пучок волокон представляет периферию нижнего квадранта сетчатки. Проходит он в латеральной части наружного коленчатого тела и подходит к вентральной губе птичьей шпоры. Предполагают, что эти проекции периферии сетчатки лежат в зрительной лучистости медиальней проекции макулярных волокон. Макулярные волокна распространяют вперед, занимая большую центральную часть зрительной лучистости в виде клина. Затем они направляются кзади и сходятся в области верхней и более низко расположенной губ птичьей шпоры.
В результате разделения периферических и центральных проекций повреждение зрительной лучистости может привести к квадрантным выпадениям поля зрения с наличием четкой горизонтальной границы.
Расположенные наиболее периферически назальные проекции сетчатки, представляющие собой «монокулярный полумесяц», собираются вблизи верхних и более низких границ дорзальных и вентральных пучков зрительной лучистости.
Нарушения в области зрительной лучистости приводят к ряду специфических нарушений полей зрения, часть которых приведена на рис. 4.2.43. Характер выпадения поля зрения во многом определяется уровнем повреждения. Причиной подобных нарушений могут быть разнообразные заболевания мозга. Наиболее часто это нарушение кровообращения (тромбоз, эмболия при гипертонической болезни, инсульт) и развитие опухоли (глиома).
В связи с тем, что нарушение структуры и функции зрительной лучистости нередко связано с нарушением кровообращения, важно знать об особенностях кровоснабжения этой области.
Кровоснабжение зрительной лучистости осуществляется на 3 уровнях:
- Часть зрительной лучистости, проходящей латерально и выше нижнего рога бокового желудочка, кровоснабжается ветвью передней ворсинчатой (хориоидальной) артерии.
- Часть зрительной лучистости, расположенной позади и латеральней рога желудочка, кровоснабжается глубокой глазной ветвью средней мозговой артерии. Последняя прони-кает в эту область через переднее перфорированное вещество совместно с латеральными полосчатыми артериями.
- При подходе зрительной лучистости к коре головного мозга кровоснабжение осуществляется перфорирующими артериями коры, главным образом, ветвями артерии птичьей шпоры. Артерия птичьей шпоры отходит от задней мозговой артерии, а иногда и от средней мозговой артерии.
Все перфорирующие артерии относятся к так называемым концевым артериям.
Продолжение в следующей статье: Функциональная анатомия зрительной системы ? Часть 4
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0