Функциональная анатомия зрительной системы │ Часть 4


Описание
Зрительная кора
Как было указано выше, системы нейронов сетчатки и наружного коленчатого тела анализируют зрительные стимулы, оценивая их цветовые характеристики, пространственный контраст и среднюю освещенность в различных участках поля зрения. Следующий этап анализа афферентных сигналов выполняется системой нейронов первичной зрительной коры (visul cortex).
Выявление участков коры головного мозга, отвечающих за обработку зрительной информации, имеет свою довольно длительную предысторию. Еще в 1782 г. студент-медик Francesco German описал белую полосу, проходящую через серое вещество затылочной доли. Именно он впервые предположил, что кора может содержать анатомически отличающиеся области. До открытия Gennari анатомы предполагали, что кора представляет собой однородную пластину ткани. Gennari даже не представлял, что он наткнулся на первичную зрительную кору. Прошло более столетия, пока Henschen доказал, что полоска Gennari соответствует первичной зрительной коре.
В связи с наличием полоски первичная зрительная кора часто называется «полосатой корой». В последующие годы, изучая цитоархитектонические особенности различных участков коры, Бродман описал 47 полей. Он произвольно обозначил первичную зрительную область под номером 17. Затем были обнаружены и другие зрительные области. При этом первичная зрительная кора получила прозаическое обозначение поля «VI» (зрительная область 1), а смежные участки (экстрастриарные зрительные области) были названы V2, V3, V4 и так далее. Таким образом, названия «первичная зрительная кора», «стриарная (полосатая) кора», «поле 17» и «VI» являются синонимами.
Этот раздел посвящен строению и функциям зрительной коры.
В настоящее время существует несколько способов изучения особенностей структурной организации и функций зрительной коры мозга. Наиболее старым является гистоморфологическое исследование коры мозга, направленное на выяснение особенностей послойного ее строения и цитологических особенностей составляющих ее нейронов. Используя этот метод, авторы до сих пор пытаются выявить границы между участками коры, отличающихся цитоархитектоникой и, естественно, функциями. Позже использовали методы, которые сводились к нанесению небольших повреждений коры экспериментальных животных. При этом спустя длительное время после травмы развивается дегенерация нервных волокон, проецирующихся на поврежденные участки, которая легко выявляется после импрегнации срезов серебром. Благодаря этому методу изучены существующие связи коры с более низко расположенными центрами.
В последующие годы разработаны методы введения антероградных трейсеров. Наиболее часто используются меченные изотопом свободные аминокислоты. После введения аминокислоты она распространяется трансаксонально, проникая через синаптические образования. Последующее проведение радиоаутографии срезов легко визуализует ход нервных волокон. К другому типу трейсеров (ретроградный) относят фермент — пероксидазу хрена. Если антероградный трейсер распространяется от периферии (глазное яблоко) к центру (зрительная кора), то пероксидаза хрена распространяется от центральных отделов ЦНС к периферическим.
Понятно, что перечисленные методы исследования можно использовать преимущественно в эксперименте. Основным методом исследования функции коры головного мозга до сих пор остается анализ симптомов, возникающих при поражениях того или иного участка коры мозга. В последние годы существенную помощь в проведении подобного анализа оказала ЯРМ-томография и электроэнцефалография. Особый интерес представляют методы прижизненной визуализации функциональной активности коры головного мозга, основанные на аппаратной регистрации уровня кровоснабжения и интенсивности метаболизма коры.
Топографические особенности зрительной коры головного мозга. Первичная зрительная кора (17-е поле по Бродману) составляет примерно 3% всей поверхности коры. Называется она еще корой птичьей шпоры, поскольку занимает верхнюю и нижнюю губы борозды птичьей шпоры. Борозда птичьей шпоры расположена на медиальной поверхности полушария мозга, но ее передний конец распространяется вниз и кзади . При этом она переходит на боковую поверхность полушария (рис. 4.2.44. 4.2.45).

Рис. 4.2.44. Внутренняя и нижняя поверхности полушария головного мозга: 1—лобная доля головного мозга; 2 — колено мозолистого тела; 3 — тело мозолистого тела; 4 — медиальная поверхность зрительного бугра; 5 — теменно-затылочная борозда; 6 - клин; 7 — задний отдел шпорной борозды; 8— язычная извилина; 9—передний отдел шпорной борозды; 10 — перешеек
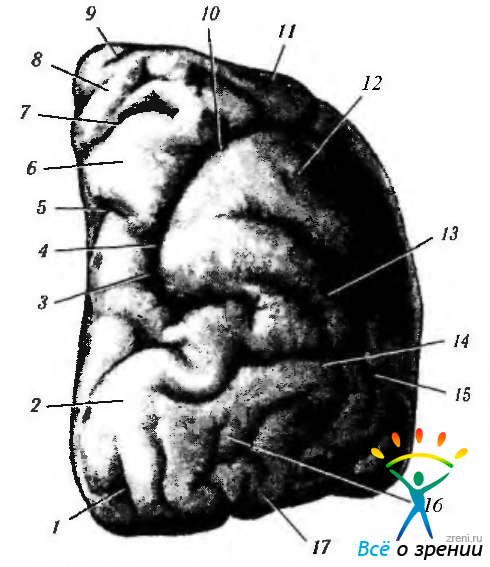
Рис. 4.2.45. Правое полушарие мозга (вид сзади): 1— извилина шпорной борозды; 2— затылочная доля; 3 — поперечная затылочная борозда; 4 — внутритеменная (интрапариетальная) борозда; 5 — теменно-затылочная борозда; 6 — верхняя теменная борозда; 7 — постцентральная верхняя борозда; 8 — поясная борозда; 9 центральная борозда; 10 верхняя теменная борозда; 11 — задняя центральная извилина; 12 — угловая извилина; 13 — надкраевая извилина; 14 — задняя теменная извилина; 15 — латеральная затылочная борозда; 16 — верхняя височная борозда; 17—медиальная височная извилина
Теменно-затылочная борозда присоединяется к борозде птичьей шпоры под небольшим углом немного спереди средней точки, разделяя ее на переднюю и заднюю части. Если теменно-затылочная борозда и борозда птичьей шпоры открыты, они кажутся разделенными вертикальной извилиной, называемой клиновидной (gyrus cuneate).
Полулунная борозда не всегда четко выражена. Она небольшая и часто пересекает борозду птичьей шпоры под прямым углом. Эта борозда отделяет первичную зрительную кору (стриарная) от вторичной коры (перистриарная). Третичная зрительная кора (парастриарная) скрыта в стенке борозды.
Часто можно обнаружить две изогнутые борозды, расположенные выше и ниже полярной борозды. Верхние полярные борозды образуют арку, направляющуюся вверх и к медиальной поверхности затылочной доли от верхней границы лунной борозды. Снизу полярная борозда изгибается книзу и вперед. Эти полярные борозды подходят в виде полумесяца к первичной зрительной коре.
Язычная извилина (gyrus lingvalis) отделяет борозду птичьей шпоры от коллатеральной борозды. Сзади она достигает полюса затылочной доли. Спереди извилина непрерывна с гиппокампальной бороздой (gyry hippocampi), располагающейся сбоку среднего мозга, и переходит в крючок извилины морского коня (uncus gyri hippocampi). Извилина повторно изгибается и формирует задне-боковую границу передней перфорирующей субстанции.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Теменно-затылочная борозда появляется в виде глубокой щели на медиальной поверхности полушария мозга. Она спускается вперед от верхне-медиальной границы мозга, приблизительно на расстоянии 5 см от затылочного полюса. Направляется она к задней части мозолистого тела. Здесь она присоединяется к борозде птичьей шпоры. На выпуклой верхнелатеральной поверхности мозга борозда продолжается на определенном расстоянии в виде боковой (латеральной) части теменно-затылочной борозды.
Цитоархитектоника первичной зрительной коры. Как было указано в начале этой главы, кора головного мозга имеет многослойное строение. Причем строение каждого слоя коры в различных отделах мозга имеет свои особенности. Выражаются они наличием различного количества слоев, размера и формы нейронов, а также их дендритными полями. Учитывая перечисленные признаки, выделено довольно большое количество полей. Как указано выше, наиболее популярной оказалась классификация Бродмана. В соответствии с этой классификацией поля 17, 18 и 19 отнесены к коре, отвечающей за обработку зрительной информации. Строение этих полей отличается от других участков коры, на чем мы остановимся ниже.
Первичная зрительная кора (поле 17) состоит из шести клеточных слоев (рис. 4.2.46— 4.2.49):
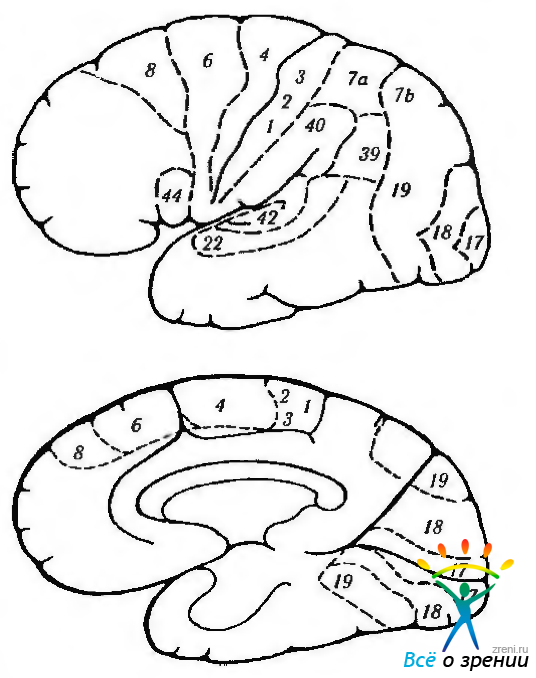
Рис. 4.2.46. Цитоархитектонические поля коры головного мозга по Бродману: а — наружная поверхность левого полушария; б — внутренняя поверхность правого полушария. Поля 17, 18 и 19 относятся к зрительной коре
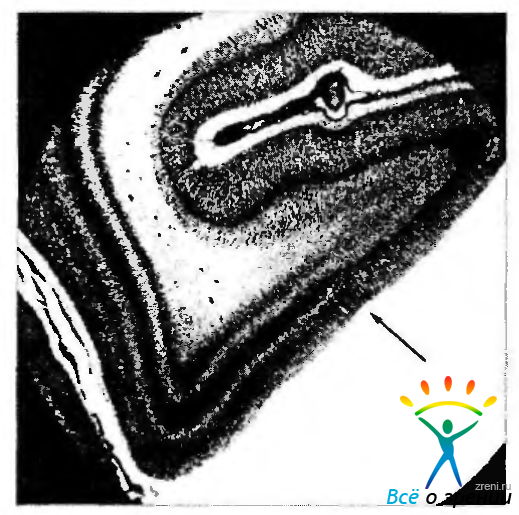
Рис. 4.2.47. Сагиттальный срез зрительного поля новорожденного в области нижнего края борозды птичьей шпоры: отмечается внезапное исчезновение полоски Дженнери (стрелка). Парастриарное поле определяется справа
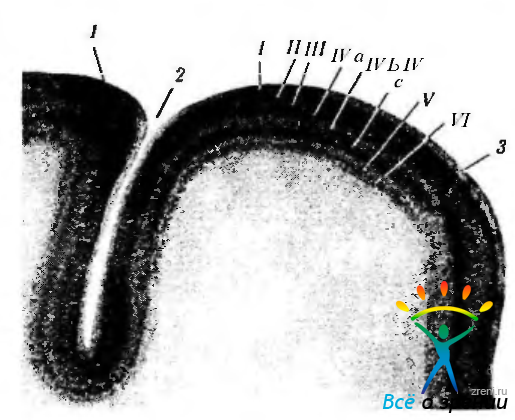
Рис. 4.2.48. Слой зрительной коры (по Vogt, 1902): 1 — стриарная кора; 2 — борозда птичьей шпоры; 3 — парастри-арная кора; 1—плексиформный слой является самым поверхностным слоем. Следующий за ним толстый темный слой состоит из трех частей — слоев II, III и IV а; II — наружный гранулярный слой является самой наружной частью толстого темного слоя; III—слой пирамидных клеток является средней частью этого слоя; II а — внутренний гранулярный слой является внутренней частью толстого темного слоя; IVb—внутренний гранулярный слой (полоска Дженнери) выглядит в виде светлой полоски; IVc — внутренний гранулярный слой; V—слой ганглиозных клеток выглядит светлым; 17 мультиформный слой является внутренним темным слоем. Необходимо обратить внимание на то, что полоска Дженнери соответствует IV Ь слою и что она исчезает на границе парастриарной коры

Рис. 4.2.49. Схематическое изображение особенностей клеточного состава зрительной коры обезьяны (no Clark, 1942) (объяснение в тексте)
- I слой. Молекулярный слой (по Бродману) является наиболее поверхностным слоем коры мозга и не отличается по строению от других участков коры мозга. Состоит он из небольших гранулярных и горизонтальных клеток.
- II слой. Наружный гранулярный слой складывается из небольших пирамидных нейронов. Обнаруживается в нем и определенное количество нейронов, отличающихся короткими аксонами. Аксоны некоторых клеток поднимаются в поверхностный слой, а другие распространяются в горизонтальной плоскости.
- III слой. Поверхностный слой пирамидных клеток. Состоит он из пирамидных клеток, которые в более глубоких слоях взаимодействуют с гранулярными клетками.
- IV слой. Этот слой состоит из подслоя, складывающегося из крупных звездчатых клеток с овальными или треугольной формы клеточными телами. Эти нейроны ориентированы беспорядочно. Их дендриты распространяются кнаружи в различных направлениях, а аксоны достигают маленьких звездчатых нейронов или распространяются в белом веществе. В глубоких участках этого слоя выявляются солитарные клетки Мейнерта и «звездчатые» пирамидные клетки, которые располагаются вблизи гранулярных клеток, малых пирамидных клеток и звездчатых клеток, отличающихся короткими аксонами. Глубокая часть IV слоя выполнена, главным образом, плотно расположенными гранулярными клетками. Многие клетки обладают короткими аксонами, распределяющимися в различных направлениях.
- V слой. Этот слой, называемый гранулярной пластинкой, состоит из плотно упакованных пирамидных нейронов различного размера, включая так называемые гигантские пирамидные клетки Мейнерта. Располагаются они в наиболее глубоких участках слоя.
- VI слой. Мультиформная пластинка по Бродману наиболее интенсивно окрашена в своей наружной части и содержит звездчатые пирамидные нейроны (слой VIа). Внутренний слой менее интенсивно окрашен и содержит среднего размера нейроны, обладающие веретеновидной и треутольной формой клеточного тела (слой VIb).
За прошедшие годы с момента описания Бродманом цитоархитектоники зрительной коры произошли некоторые уточнения цитологических особенностей нейронов. При этом определены их функциональные особенности. Мы позволим себе кратко привести эти сведения, полученные Van Essen, Deyoe. Вышеприведенные авторы дифференцируют следующие типы нейронов первичной зрительной коры:
- Шипоподобные звездчатые клетки. Располагаются они в слоях 4С а и 4С b первичной зрительной коры. Относятся к локальным экс-цитатным нейронам.
- Гладкие звездчатые клетки. Располагаются они в слоях 4С а и 4С Ь. Относятся к локальным тормозным нейронам.
- Простые пирамидные клетки. Обнаруживаются в слоях 4В. 2 и 3. Относятся к эксцитатным нейронам.
- Комплексные пирамидные клетки. Обнаруживаются в слоях 2, 3, 5 и 6. Относятся к эксцитатным нейронам.
Миелоархитектоника первичной зрительной коры довольно существенно отличается от других участков коры мозга. Четвертый слой содержит белую полоску, описанную German. Видна эта полоска невооруженным глазом. В связи с этим четвертый слой подразделяют на три подслоя — А. В и С. Зрительные афференты оканчиваются преимущественно в IV слое (рис. 4.2.50).
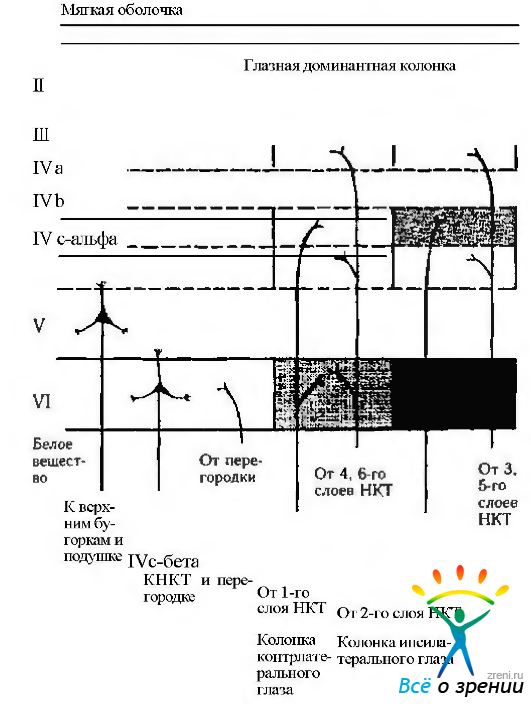
Рис. 4.2.50. Особенности связей зрительной коры (по Каnclell, 1983): проекции от магноцеллюлярного слоя наружного коленчатого тела завершаются в слоях IVc-альфа (и в VI слое), в то время как проекции парвоцеллюлярных слоев завершаются в слое IVc-бета (и распространяются на слои IVa и VI). Вход от каждого глаза формирует доминантные колонки. Колонки ипсилатерального и контрлатерального глаза (вертикально ориентированные колонки 500 мкм шириной и несколько миллиметров длиной) прилежат друг к другу, формируя регулярную структуру. Зрительная кора проецируется на наружное коленчатое тело, перегородку (claustnim), верхние бугорки четверохолмия, подушку зрительного бугра и другие структуры. Проекции на верхние бугорки и подушку исходят из пирамидных клеток V слоя, а на наружное коленчатое тело и перегородку — от пирамидных клеток VI слоя
Аксоны, идущие он нейронов наружного коленчатого тела, первоначально проникают в IV слой, а затем распространяются на V и VI слои. При этом большая часть терминалов афферентных волокон формируется в IV слое.
Электронномикроскопические исследования позволили выявить большое разнообразие типов синаптических образований в этих слоях первичной зрительной коры (сферические, шайбоподобные и др.). Часть синапсов — эксцитатные, а часть — тормозные. Завершаются аксоны нейронов наружного коленчатого тела в IV слое на телах звездчатых нейронов, а часть на телах пирамидных.
Подводя итоги обсуждения особенностей архитектоники первичной зрительной коры, необходимо выделить основные структурные отличия этого поля:
- Зрительная кора тоньше (1,5 мм), чем кора других участков мозга. Это связано с тем. что нейропиль выражен менее четко. При этом количество нейронов больше.
- Внутренний зернистый слой (IV) толще, а нейроны, его формирующие, лежат довольно плотно. Наружный и особенно внутренний зернистые слои состоят из большого количества плотно расположенных мелких клеток.
- Основное шестислойное строение коры усложняется наличием дополнительной полоски в слое IV. В результате этого данный слой подразделяют на подслои — IVA, IVB и IVC.
- Зрительная полоска (IVB) содержит большое количество горизонтально расположенных звездчатых нейронов.
- Ганглиозный слой (V) содержит пирамидные нейроны Мейнерта размером 3 мкм. Эти клетки проецируются на верхние бугорки четверохолмия и, возможно, на ядра глазодвигательных нервов.
- Морфологические особенности дендритных полей нейронов зрительной коры свидетельствуют о том, что они принимают участие в интегративных зрительных процессах.
Проекция сетчатки на первичную зрительную кору. Проекция сетчатки на зрительную кору головного мозга была изучена благодаря сопоставлению клинических проявлений повреждения различных участков коры, обнаруживаемых при аутопсии. Подобные исследования проведены как у человека, так и у экспериментальных животных.
Впервые наиболее полные данные относительно особенностей проекции сетчатки на зрительную кору были получены Holmes, Lister при анализе выпадения полей зрения при повреждении коры головного мозга у солдат, раненных во время первой мировой войны. Схожие исследования были проведены Minkovski во время русско-японской войны. Holmes описал проекцию монокулярных, бинокулярных и макулярных волокон. В соответствии с полученными этими и другими исследователями сведениями каждая область сетчатки представлена в определенной области зрительной коры. При этом сохраняется основной принцип проекции — «точка к точке». Все зрительные сигналы, исходящие от корреспондирующих половин сетчаток, передаются одному коленчатому ядру, а затем первичной зрительной коре той же стороны. Так, нервные волокна, идущие от темпоральной половины сетчатки правого глаза, проходят через правый зрительный тракт. От назальной половины левого глаза волокна перекрещиваются в зрительном перекресте. Затем они присоединяются к неперекрещенным волокнам правого глаза и достигают правого наружного коленчатого тела. Далее они направляются к первичной зрительной коре правого полушария.
Верхние и нижние квадранты сетчатки представлены ниже и выше шпорной борозды, соответственно. Периферия сетчатки проецируется на передние отделы, а макула ближе к затылочному полюсу. На наиболее передние части зрительной коры проецируются на самые периферические части назального поля сетчатки, что соответствует монокулярному «височному полумесяцу».
Макула по площади представлена в зрительной коре значительно больше, чем в сетчатой оболочке (рис. 4.2.51, 4.2.52).

Рис. 4.2.51. Представительство полей зрения в стриарной коре (по Miller, 1952): a —правое поле зрения; б — стриарная кора (вид сзади); в — стриарная кора (вид с медиальной поверхности мозга)
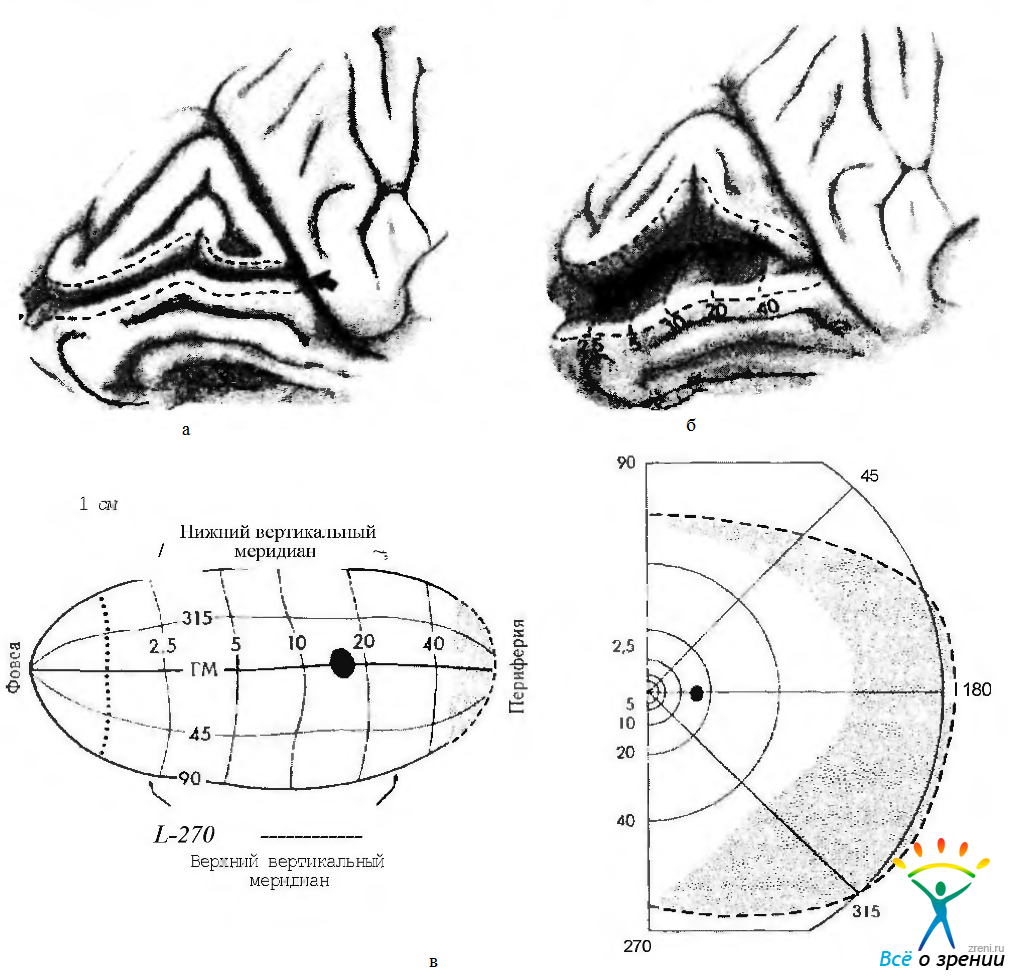
Рис. 4.2.52. Проекция зрительного поля на зрительную кору (по Horton, Hovt, 1991): а — левая затылочная доля. Локализация стриарной коры в пределах борозды птичьей шпоры. Граница (пунктирная линия) между стриарной корой (VI) и экстрастриарной корой (V2) содержит представительство вертикального меридиана; б— вид стриарной коры после открытия губ борозды птичьей шпоры. Пунктирные линии отмечают координаты зрительного поля. Представительство горизонтального меридиана локализуется приблизительно вдоль основания щели птичьей борозды. Вертикальные пунктирные линии отмечают изоэксцентричные контуры от 2,5 до 40 градусов. Стриарная кора как бы обернута вокруг полюса затылочной доли и распространяется на протяжении 1 см. на латеральную поверхность, где и локализуется проекция макулы; в — плоское схематическое изображение проекции правого полуполя на левую зрительную кору. Ряд точек указывает приблизительную локализацию щели
Благодаря этому иногда довольно обширные повреждения затылочной коры не сопровождаются нарушением центрального зрения.
Волокна, обеспечивающие моно- и бинокулярное зрение на разных уровнях зрительного пути, разделены. Как указано выше, монокулярное поле у человека представлено крайней периферией темпорального поля. При этом вовлекается наиболее назальная часть сетчатки. Волокна формируют «назальный полумесяц», расположенный медиальней перекрещивающихся волокон, а затем представлены маленьким пучком в вентральной части зрительного тракта и наружного коленчатого тела.
В зрительной лучистости верхние волокна находятся в верхнем квадранте, а нижние располагаются в нижнем квадранте. Монокулярное поле проецируется на переднюю часть нижней губы шпорной борозды
Связи первичной зрительной коры. Связи первичной зрительной коры достаточно многочисленны. Основными из них являются следующие:
- Поле 17 связано со зрительной корой второго полушария посредством волокон, про ходящих в мозолистом теле (более характерна подобная связь для полей 18 и 19).
- Зрительные поля связаны со зрительными полями лобной доли. Особенно это характерно для ассоциативной коры (поле 19).
- Поле 17 связано с верхними бугорками четверохолмия (через 19 поле).
- Поле 17 связано с ядрами глазодвигательного нерва и другими ядрами благодаря нисходящим волокнам, направляющимся к зри тельной лучистости (особенно от области 19).
Перечисленные связи являются анатомической основой зрительно-тактильной, зрительно-слуховой и других ассоциативных функций, включая движение глаза. Функциональное значение этих связей будет описано несколько ниже, а также приведено в разделе «Головной мозг».
Физиология зрительной коры. Перед тем как описать структурно-функциональную организацию зрительной коры, необходимо привести хотя бы элементарные сведения относительно ее физиологии. Это необходимо не только потому, что электрофизиологические исследования явились основой дальнейшего углубленного изучения структуры коры с целью объяснения связи между функцией и структурой, но и потому, что без четкого представления функций зрительной коры исключительно сложно понять особенности ее нейронных связей.
Как указывалось выше, раньше предполагали, что основной функцией зрительного тракта, наружных коленчатых тел и зрительной лучистости является выполнение релейной функции. Предполагалось, что сигналы от сетчатки передаются коре головного мозга без какой-либо существенной обработки. К настоящему времени получены новые сведения, существенно расширяющие наши знания о функциях перечисленных структур и, в первую очередь, наружного коленчатого тела. Уже само несоответствие между количеством фоторецепторов и ганглиозных клеток указывает на наличие конвергенции информации на уровне сетчатки. Коллатеральные синаптические связи между нейронами сетчатки обеспечивают боковое торможение, усиливающее контрастность между стимулируемыми и нестимулируемыми точками. Именно они создают циркулярные рецептивные поля ON/OFF-типа, являющиеся фундаментальными структурно-функциональными единицами обработки зрительной информации.
Функция рецептивных полей изменяется по мере продвижения к коре головного мозга. Довольно существенная обработка закодированной информации происходит в наружном коленчатом теле, которое отличается сложностью межнейронных связей. Как указано выше, в наружном коленчатом теле также существует концентрическая организация рецептивных полей. Эта концентрическая организация напоминает рецептивные поля сетчатой оболочки. При этом рецептивные поля наружного коленчатого тела реагируют на линейные «зрительные стимулы», а также на направление движения объекта.
И, наконец, окончательная обработка зрительной информации происходит в коре головного мозга, как в первичной зрительной коре, так и в многочисленных ассоциативных полях.
Hubei, Wiesel первыми описали свойства рецептивных полей зрительной коры. Авторы после подведения микроэлектрода к нейрону IV слоя зрительной коры освещали сетчатую оболочку различными по форме, размеру, направленности и скорости движения «зрительными стимулами». В результате эксперимента авторы получали возможность регистрировать активность отдельного нейрона зрительной коры при возбуждении сетчатки различными стимулами. Анализируя сигналы нейрона Hubei и Wiesel обнаружили новые типы рецептивных полей зрительной коры, с которыми они никогда не сталкивались при регистрации потенциала действия нейронов сетчатки и наружного коленчатого тела. Классифицировали они эти поля следующим образом.
«Простые клетки». «Простые клетки» обнаруживаются в 17-м поле. Рецептивные поля «простых клеток» при освещении сетчатки световым пятном небольшого размера напоминают рецептивные поля наружного коленчатого тела (концентрические поля ON/OFF). Принципиально они отличаются между собой пространственным расположением «эксцитатных» и «тормозных» доменов. «Простые клетки» не формируют концентрические структуры, свойственные рецептивным полям сетчатки и наружного коленчатого тела. Организованы они в параллельные, рядом расположенные подполя. Наиболее часто узкие длинные «эксцитатные» или «тормозные» области зажаты между двумя симметричными подобластями противоположного типа. Некоторые подполя имеют различный размер. При медленном (2—4 градуса с) изменении ориентации световой щели, падающей на сетчатую оболочку, можно получить максимальный ответ таких рецептивных полей при совпадении ориентации щели с осью рецептивного поля. Ориентационная избирательность «простых клеток», таким образом, является их основной особенностью. Благодаря разнообразию строения этих рецептивных полей в зрительной коре фактически представлены все возможные изменения пространственной ориентации зрительного сигнала.
Рецептивные поля «простых клеток» также отвечают на перемещающиеся изображения, представленные в виде пластинки, разреза или грани. Рецептивные поля «простых клеток» обычно формируют потенциалы действия в тот момент, когда перемещающаяся световая щель входит в ON-область. Наиболее интенсивный ответ наблюдается тогда, когда объект пересекает границу, проходящую между ON- и OFF-центрами. Клетки с симметричным расположением подполей формируют потенциал действия при движении светового стимула в любом направлении. Клетки с асимметричными подполями часто дают неодинаковые ответы при движении светового стимула во встречных направлениях. Оптимальная скорость движения светового стимула также регистрируется «простыми клетками».
«Комплексные клетки». «Комплексные клетки» обнаружены в полях 17, 18 и 19. В рецептивных полях комплексных клеток невозможно выявить «эксцитатные» и «тормозные» подобласти при использовании неподвижных световых стимулов и стимулов в виде пятна. При движении объекта электрофизиологически можно выявить ON- и OFF-ответы, но сила ответа зависит от направления движения светового стимула (щель) относительно рецептивного поля. Максимальный ответ возникает только при движении светового стимула (щель) поперек рецептивного поля. Необходимо отметить и то, что если максимальный ответ рецептивных полей «простых клеток» возникает в момент пересечения светового стимула внутренней границы рецептивного поля (граница между антагонистическими подобластями), то рецептивные поля комплексных клеток возбуждаются при перемещении объекта в любом месте рецептивного поля (рис. 4.2.53, а).

Рис. 4.2.53. Рецептивные поля комплексной (а) и энд-стопт (б) клеток первичной зрительной коры обезьяны и особенности их ответа (по Hubei, Weisel, 1968): пунктирная линия отмечает границы рецептивного поля. Ориентация «зрительного стимула» показана слева, а особенности реакции нейрона — справа (объяснение в тексте)
Выявлено несколько типов комплексных рецептивных полей. Одним из них являются так называемые энд-стопт (end-stopped) клетки. Отличаются эти рецептивные поля тем, что они возбуждаются только в том случае, когда возбуждающий сигнал не превышает размер рецептивного поля (рис. 4.2.53, б). Максимальный ответ возникает тогда, когда световая щель или пластинка равна длине рецептивного поля. При превышении размера световым сигналом рецептивного поля ответ не увеличивается. Подобные рецептивные поля напоминают один из вариантов рецептивного поля «комплексной клетки», свойством которой является то, что при превышении световым сигналом длины рецептивного поля ответ этого поля снижается.
«Гиперкомплексные клетки». Функции «простых», «комплексных» и «гиперкомплексных» рецептивных полей различны. «Простые клетки» реагируют на сконцентрированный или рассеянный свет, а «комплексные» — анализируют контрастность, наличие границы между объектами с различной степенью освещения.
Чувствительны они и при определении направленности освещения и скорости движения объекта.
Hubei и Wiesel выявили, что поле 18 содержит 90—95% «комплексных клеток», а в 19-м поле 58% клеток можно отнести к «гиперкомплексным клеткам». «Гиперкомплексные клетки» в меньшем количестве обнаружены и в полях 17 и 18. Авторы отнесли выявленные клетки к клеткам третьего порядка обработки зрительной информации. Их особенностью является то, что «гиперкомплексные клетки» отвечают на зрительные стимулы, отличающиеся сложным строением. Сложность стимула характеризуется его различной формой, контрастностью, скоростью и направлением его движения и др. Авторами выявлено 6 вариантов организации «гиперкомплексной клетки». Более подробно об особенностях различных типов «гиперкомплексной клетки» можно прочитать в ряде работ, посвященных электрофизиологии зрительного восприятия.
Важно отметить, что «комплексные» и «гиперкомплексные» клетки получают информацию от обоих глаз и, таким образом, обеспечивают бинокулярное зрение.
Иерархия рецептивных полей. Рецептивные поля в функциональном отношении существенно отличаются на различных уровнях зрительного пути.
Возникает вопрос, каким образом нейроны наружного коленчатого тела генерируют рецептивные поля корковых клеток? «Простые клетки» сконцентрированы в IV слое первичной зрительной коры, т. е. в том же самом слое, который принимает большую часть проекции от наружного коленчатого тела. «Простые клетки» также обнаруживаются в VI слое. Аксоны «простых клеток» разветвляются и формируют многочисленные синапсы на клетках других слоев коры.
«Комплексные клетки» обычны для всех слоев коры, кроме уровня IV. Из этого вытекает логический вывод — «простые клетки» IV и VI слоев проецируются на «комплексные клетки», расположенные в наружной части IV слоя.
На основании этого Hubei и Wiesel предположили, что простые клеточные рецептивные поля созданы из рецептивных полей наружного коленчатого тела. Например, рецептивное поле «простой клетки» может быть сгенерировано эксцитатным входом от ряда коленчатых клеток с центром, выстроенным в линию, как показано на рис. 4.2.54, а.
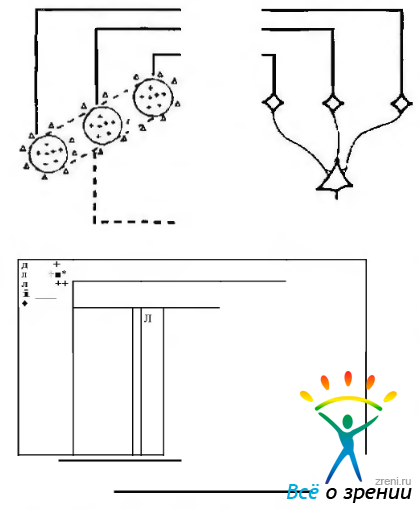
Рис. 4.2.54. Схематические изображения, иллюстрирующие гипотезу Hubei. Weisel (1962) относительно организации простых (а) и комплексных (б) рецептивных полей: а — большое количество нейронов наружного коленчатого тела, четыре из которых представлено справа, обладает рецептивным полем с ON-центром, расположенным вдоль плоскости сетчатки. Все они проецируются на «простую клетку» коры и синапсы становятся эксцитатными. Рецептивное поле кортикальной клетки, в результате этого будет обладать продолговатым рецептивным полем с ON-центром, показанным слева; б - определенное количество клеток с простым рецептивным полем проецируется на одну «комплексную клетку». В этом случае «простая клетка» обладает продолговатым рецептивным полем, содержащим эксцитатные и тормозные зоны. Граница поля ограничена пунктирной линией. Последовательная активация вертикального края движущимся стимулом поперек этого прямоугольника, независимо от его положения, будет активировать «простые клетки» и вызывать эксцитатный ответ более высоко расположенного нейрона
В то же время они считают, что рецептивные поля «комплексных клеток» построены из рецептивных полей «простых клеток» (рис. 4.2.54, б). Таким образом, нейроны с «концентрическими», «сложными» и «сверхсложными» рецептивными полями образуют последовательно усложняющиеся этапы анализа сигналов, включая цветовые, в коре головного мозга. Это находит свое отражение и в характере распределения нейронов с цветовыми рецептивными полями по слоям коры. Если клетки с «концентрическими» и «простыми» рецептивными полями обнаруживаются в III и IV слоях коры, тесно связанных с входами наружного коленчатого тела, то клетки со «сложными» и «сверхсложными» рецептивными полями проецируются на верхние (II и верхняя часть III слоя) и нижние (V и VI) слои коры. Из выше приведенных данных видно, что сейчас свойства рецептивных полей различного типа довольно хорошо определены. В то же время их роль в зрительном восприятии пока не полностью ясна. Тем не менее не ясно, благодаря каким нейронам и на каком уровне происходит формирование «зрительного восприятия» как высшей формы обработки зрительной информации.
Продолжение в следующей статье: Функциональная анатомия зрительной системы ? Часть 5
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0