Гидродинамика глаза


Содержание:
Описание
В глазном яблоке содержится несколько гидродинамических систем, связанных с циркуляцией водянистой влаги, влаги стекловидного тела, увеальной тканевой жидкости и крови. Циркуляция внутриглазных жидкостей обеспечивает нормальный уровень внутриглазного давления и питание всех тканевых структур глаза.Вместе с тем глаз представляет собой сложную гидростатическую систему, состоящую из полостей и щелей, разделенных эластичными диафрагмами. От гидростатических факторов зависит сферическая форма глазного яблока, правильное положение всех внутриглазных структур, нормальное функционирование оптического аппарата глаза. Гидростатический буферный эффект обусловливает устойчивость тканей глаза к повреждающему действию механических факторов. Нарушения гидростатического равновесия в полостях глаза приводят к существенным изменениям в циркуляции внутриглазных жидкостей и развитию глаукомы. При этом наибольшее значение имеют нарушения в циркуляции водянистой влаги, основные особенности которой рассмотрены ниже.
↑ Водянистая влага
Водянистая влага заполняет переднюю и заднюю камеры глаза и по специальной дренажной системе оттекает в эпи- и интрасклеральные вены. Таким образом, водянистая влага циркулирует преимущественно в переднем сегменте глазного яблока. Она участвует в метаболизме хрусталика, роговой оболочки и трабекулярного аппарата, играет важную роль в поддержании определенного уровня внутриглазного давления. Глаз человека содержит около 250—300 мм3, что составляет примерно 3—4% от общего объема глазного яблока.
Композиция водянистой влаги существенно отличается от состава плазмы крови. Ее молекулярная масса составляет всего 1,005 (плазмы крови — 1,024), в 100 мл водянистой влаги содержится 1,08 г сухого вещества (в 100 мл плазмы крови — более 7 г). Внутриглазная жидкость более кислая, чем плазма крови, в ней повышено содержание хлоридов, аскорбиновой и молочной кислот. Избыток последней, по-видимому, связан с метаболизмом хрусталика. Концентрация аскорбиновой кислоты во влаге в 25 раз выше, чем в плазме крови. Основными катионами являются калий и натрий.
Неэлектролитов, особенно глюкозы и мочевины, во влаге содержится меньше, чем в плазме крови. Недостаток глюкозы можно объяснить утилизацией ее хрусталиком. Водянистая влага содержит лишь небольшое количество белков — не более 0,02%, пропорция альбуминов и глобулинов такая же, как в плазме крови. В камерной влаге обнаружены также в небольшом количестве гиалуроновая кислота, гексозамин, никотиновая кислота, рибофлавин, гистамин, креатин. По данным А. Я. Бунина и А. А. Яковлева (1973), водянистая влага содержит буферную систему, обеспечивающую постоянство pH путем нейтрализации продуктов метаболизма внутриглазных тканей.
Водянистая влага образуется главным образом отростками цилиарного (ресничного) тела. Каждый отросток состоит из стромы, широких тонкостенных капилляров и двух слоев эпителия (пигментного и непигментного). Эпителиальные клетки отделены от стромы и задней камеры наружной и внутренней пограничными мембранами. Поверхности непигментных клеток имеют хорошо развитые оболочки с многочисленными складками и вдавлениями, как это обычно бывает у секреторных клеток.
Основным фактором, обеспечивающим отличие первичной камерной влаги от плазмы крови, является активный транспорт субстанций. Каждое вещество переходит из крови в заднюю камеру глаза с характерной для этого вещества скоростью. Таким образом, влага в целом является интегральной величиной, слагающейся из отдельных обменных процессов [Caprioli J., 1987].
Ресничный эпителий осуществляет не только секрецию, но и реабсорбцию некоторых веществ из водянистой влаги. Реабсорбция осуществляется через специальные складчатые структуры клеточных мембран, которые обращены к задней камере. Доказано, что из влаги кровь активно переходят йод и некоторые органические ионы.
Механизмы активного транспорта ионов через эпителий цилиарного тела изучены недостаточно. Полагают, что ведущую роль в этом играет натриевая помпа, с помощью которой в заднюю камеру поступает около 2/3 ионов натрия. В меньшей степени за счет активного транспорта в камеры глаза поступают ионы хлора, калия, бикарбонаты, а также аминокислоты. Механизм перехода аскорбиновой кислоты в водянистую влагу неясен. При концентрации аскорбата в крови выше 0,2 ммоль/кг механизм секреции насыщается, поэтому повышение концентрации аскорбата в плазме крови выше этого уровня не сопровождается его дальнейшей аккумуляцией в камерной влаге. Активный транспорт некоторых ионов (особенно Na ) ведет к гипертоничности первичной влаги. Это служит причиной поступления в заднюю камеру глаза воды за счет осмоса. Первичная влага непрерывно разбавляется, поэтому концентрация большинства неэлектролитов в ней ниже, чем в плазме.
Таким образом, водянистая влага продуцируется активно. Энергетические затраты на ее образование покрываются за счет метаболических процессов в клетках эпителия цилиарного тела и деятельности сердца, благодаря которой поддерживается достаточный для ультрафильтрации уровень давления в капиллярах цилиарных отростков.
Большое влияние на композицию оказывают процессы диффузии. Липоидорастворимые вещества проходят через гематоофтальмический барьер тем легче, чем выше их растворимость в жирах. Что касается жиронерастворимых субстанций, то они выходят из капилляров через щели в их стенках со скоростью, обратно пропорциональной размеру молекул. Для веществ, имеющих молекулярную массу больше 600, гематоофтальмический барьер практически непроницаем. Исследования с применением радиоактивных изотопов показали, что одни вещества (хлор, тиоционат) входят в глаз путем диффузии, другие (аскорбиновая кислота, бикарбонат, натрий, бром) — посредством активного транспорта [Becker В., 1961].
В заключение отметим, что в образовании водянистой влаги принимает участие (хотя и весьма небольшое) ультрафильтрация жидкости. Средняя скорость продуцирования водянистой влаги равна примерно 2 мм /мин, следовательно, в течение 1 сут через передний отдел глаза протекает около 3 мл жидкости.
↑ Камеры глаза
Водянистая влага сначала поступает в заднюю камеру глаза, которая представляет собой щелевидное пространство сложной конфигурации, расположенное кзади от радужки. Экватор хрусталика делит камеру на переднюю и заднюю части (рис. 3).

Рис. 3. Камеры глаза (схема). 1 — шлеммов канал; 2 — передняя камера; 3 — передний и 4 — задний отделы задней камеры; 5 — стекловидное тело.
В нормальном глазу экватор отделен от цилиарной короны промежутком шириной около 0,5 мм, и этого вполне достаточно для свободной циркуляции жидкости внутри задней камеры. Это расстояние зависит от рефракции глаза, толщины цилиарной короны и размеров хрусталика. Оно больше в миопическом и меньше в гиперметропическом глазу. При некоторых условиях хрусталик как бы ущемляется в кольце цилиарной короны (цилиохрусталиковый блок).
Задняя камера соединяется с передней через зрачок. При плотном прилегании радужки к хрусталику переход жидкости из задней камеры в переднюю затруднен, что приводит к повышению давления в задней камере (относительный зрачковый блок). Передняя камера служит основным резервуаром для водянистой влаги (0,15— 0,25 мм ). Изменения ее объема сглаживают случайные колебания офтальмотонуса.
Особенно важную роль в циркуляции водянистой играет периферическая часть передней камеры, или ее угол (УПК). Анатомически различают следующие структуры УПК: вход (апертура), бухту, переднюю и заднюю стенки, вершину угла и нишу (рис. 4).

Рис. 4. Угол передней камеры. 1 — трабекула; 2 — шлеммов канал; 3 — ресничная мышца; 4 — склеральная шпора. Ув. 140.
Вход в угол расположен там, где оканчивается десцеметова оболочка. Задней границей входа служит радужка, которая образует здесь последнюю к периферии складку стромы, получившую название «складка Фукса». К периферии от входа расположена бухта УПК. Передней стенкой бухты служат трабекулярная диафрагма и склеральная шпора, задней — корень радужки. Корень — наиболее тонкая часть радужки, так как содержит только один слой стромы. Вершина УПК занята основанием цилиарного тела, которое имеет небольшую выемку — нишу УПК (angle recess). В нише и рядом с нею часто расположены остатки эмбриональной увеальной ткани в виде тонких или широких тяжей, идущих от корня радужки к склеральной шпоре или дальше к трабекуле (гребенчатая связка).
↑ Дренажная система глаза
Дренажная система глаза расположена в наружной стенке УПК. Она состоит из трабекулярной диафрагмы, склерального синуса и коллекторных канальцев. К дренажной зоне глаза относят также склеральную шпору, цилиарную (ресничную) мышцу и вены-реципиенты.
↑ Трабекулярный аппарат
Трабекулярный аппарат имеет несколько названий: «трабекула (или трабекулы)», «трабекулярная диафрагма», «трабекулярная сеть», «решетчатая связка». Он представляет собой кольцевидную перекладину, переброшенную между передним и задним краями внутренней склеральной бороздки. Эта бороздка образуется за счет истончения склеры около ее окончания у роговицы. На разрезе (см. рис. 4) трабекула имеет треугольную форму. Верхушка ее прикреплена к переднему краю склеральной бороздки, основание связано со склеральной шпорой и частично с продольными волокнами цилиарной мышцы. Передний край бороздки, образованный плотным пучком круговых коллагеновых волокон, получил название «переднее пограничное кольцо Швальбе». Задний край — склеральная. шпора — представляет собой выступ склеры (напоминающий на разрезе шпору), который прикрывает изнутри часть склеральной бороздки. Трабекулярная диафрагма отделяет от передней камеры щелевидное пространство, которое называют венозной пазухой склеры, шлеммовым каналом или склеральным синусом. Синус связан тонкими сосудами (выпускники, или коллекторные канальцы) с эпи- и интрасклеральными венами (вены-реципиенты).
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Трабекулярная диафрагма состоит из трех основных частей:
- увеальной трабекулы,
- корнеосклеральной трабекулы
- и юкстаканаликулярной ткани.
Наружный слой трабекулярного аппарата, прилежащий к шлеммову каналу, значительно отличается от других трабекулярных слоев. Его толщина варьирует от 5 до 20 мкм, увеличиваясь с возрастом. При описании этого слоя используют различные термины: «внутренняя стенка шлеммова канала», «пористая ткань», «эндотелиальная ткань (или сеть)», «юкстаканаликулярная соединительная ткань» (рис. 5).
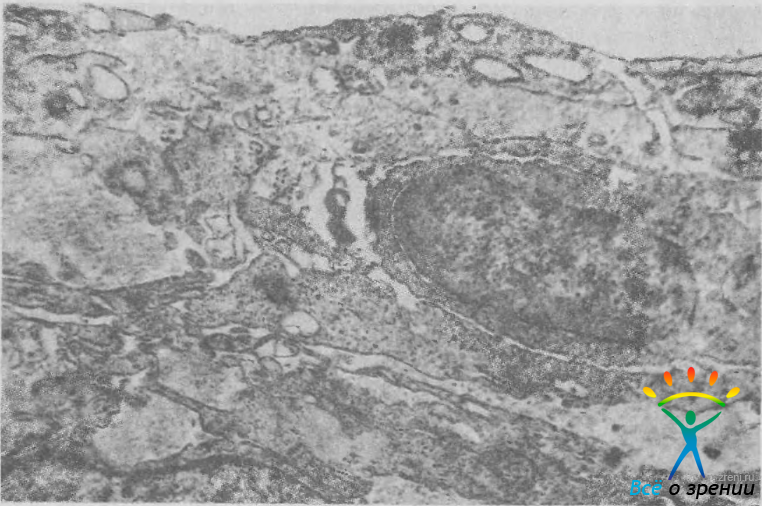
Рис. 5. Электронограмма юкстаканаликулярной ткани. Под эпителием внутренней стенки шлеммова канала расположена рыхлая волокнистая ткань, содержащая гистиоциты, коллагеновые и эластические волокна, экстрацеллюлярный матрикс. Ув. 26 000.
Юкстаканаликулярная ткань состоит из 2—5 слоев фиброцитов, свободно и без определенного порядка лежащих в рыхлой волокнистой ткани. Клетки похожи на эндотелий трабекулярных пластин. Они имеют звездчатую форму, их длинные, тонкие отростки, соприкасаясь друг с другом и с эндотелием шлеммова канала, образуют своеобразную сеть. Экстрацеллюлярный матрикс является продуктом эндотелиальных клеток, он состоит из эластических и коллагеновых фибрилл и гомогенной основной субстанции. Установлено, что в состав этой субстанции входят чувствительные к гиалуронидазе кислые мукополисахариды. В юкстаканаликулярной ткани много нервных волокон такого же характера, как и в трабекулярных пластинах.
↑ Шлеммов канал
Шлеммов канал, или склеральный синус, представляет собой циркулярную щель, расположенную в задненаружной части внутренней склеральной бороздки (см. рис. 4). От передней камеры глаза он отделен трабекулярным аппаратом, кнаружи от канала расположен толстый слой склеры и эписклеры, содержащий поверхностно и глубоко расположенные венозные сплетения и артериальные веточки, участвующие в формировании краевой петлистой сети вокруг роговицы. На гистологических срезах средняя ширина просвета синуса составляет 300—500 мкм, высота — около 25 мкм. Внутренняя стенка синуса неровная и местами образует довольно глубокие карманы. Просвет канала чаще одиночный, но может быть двойным и даже множественным. В некоторых глазах он разделен перегородками на отдельные отсеки (рис. 6).
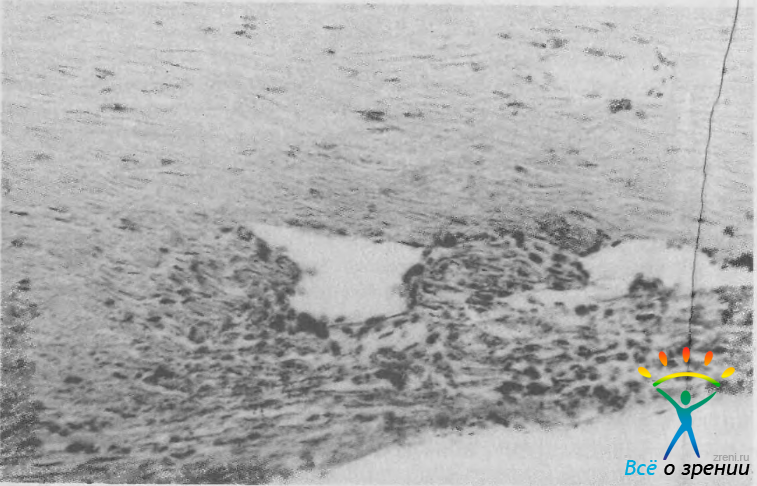
Рис. 6. Дренажная система глаза. В просвете шлеммова канала видна массивная перегородка. Ув. 220.
Эндотелий внутренней стенки шлеммова канала представлен очень тонкими, но длинными (40—70 мкм) и довольно широкими (10—15 мкм) клетками. Толщина клетки в периферических отделах около 1 мкм, в центре она значительно толще за счет крупного ядра округлой формы. Клетки образуют сплошной слой, но их концы не накладываются друг на друга (рис. 7),
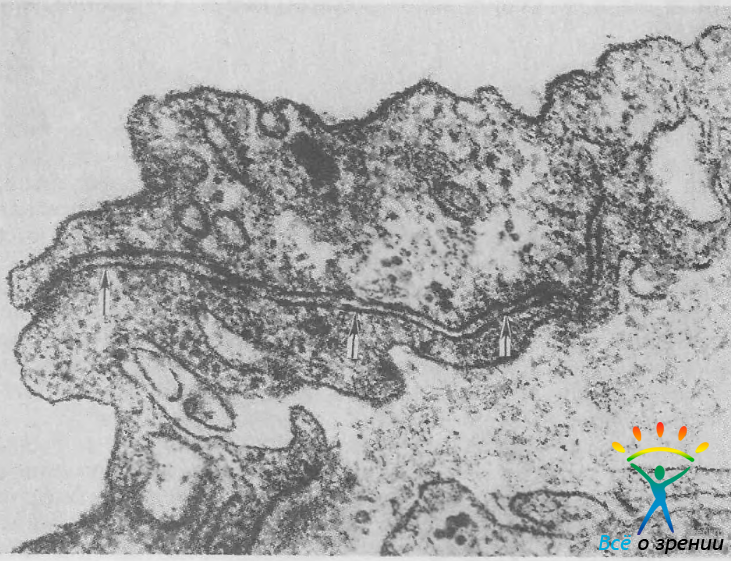
Рис. 7. Эндотелий внутренней стенки шлеммова канала. Две соседние клетки эндотелия разделены узким щелевидным пространством (стрелки). Ув. 42 000.
поэтому возможность фильтрации жидкости между клетками не исключается. С помощью электронной микроскопии в клетках обнаружены гигантские вакуоли, расположенные преимущественно в околоядерной зоне (рис. 8).
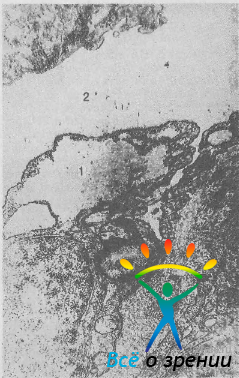
Рис. 8. Гигантская вакуоль (1), расположенная в клетке эндотелия внутренней стенки шлеммова канала (2). Ув. 30 000.
Одна клетка может содержать несколько вакуолей, имеющих овальную форму, максимальный диаметр которых варьирует от 5 до 20 мкм [Tripathi R., 1972]. По данным Н. Inomata и соавт. (1972), на 1 мм длины шлеммова канала приходится 1600 эндотелиальных ядер и 3200 вакуолей. Все вакуоли открыты в сторону трабекулярной ткани, но только часть из них имеет поры, ведущие в шлеммов канал. Величина отверстий, связывающих вакуоли с юкстаканаликулярной тканью, составляет 1—3,5 мкм, со шлеммовым каналом — 0,2—1,8 мкм.
Эндотелиальные клетки внутренней стенки синуса не имеют выраженной базальной мембраны. Они лежат на очень тонком неравномерном слое волокон (преимущественно эластических), связанных с основной субстанцией. Короткие эндоплазматические отростки клеток проникают в глубь этого слоя, в результате чего увеличивается прочность их соединения с юкстаканаликулярной тканью.
Эндотелий наружной стенки синуса отличается тем, что не имеет крупных вакуолей, ядра клеток плоские и эндотелиальный слой лежит на хорошо сформированной базальной мембране.
↑ Коллекторные канальцы, венозные сплетения
Кнаружи от шлеммова канала, в склере, расположена густая сеть сосудов — интрасклеральное венозное сплетение, другое сплетение расположено в поверхностных слоях склеры. Шлеммов канал связан с обоими сплетениями так называемыми коллекторными канальцами, или выпускниками. По данным Ю. Е. Батманова (1968), количество канальцев варьирует от 37 до 49, диаметр — от 20 до 45 мк. Большинство выпускников начинается в заднем отделе синуса. Можно выделить четыре типа коллекторных канальцев:
- узкие короткие коллекторы, связывающие синус с интрасклеральным венозным сплетением;
- одиночные крупные сосуды, которые выходят на поверхность склеры и впадают в эписклеральные вены (рис. 9);
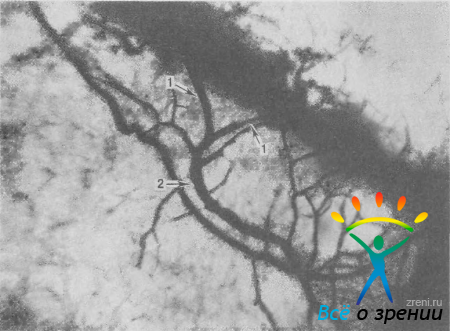 Рис. 9. Дренажная система глаза, перфузированная раствором туши. Два выпускника шлеммова канала (1) соединяются и образуют водяную вену (2). Ув. 45.
Рис. 9. Дренажная система глаза, перфузированная раствором туши. Два выпускника шлеммова канала (1) соединяются и образуют водяную вену (2). Ув. 45. - короткие каналы, которые отходят от синуса, идут параллельно ему и вновь впадают в шлеммов канал;
- отдельные сосуды, связывающие шлеммов канал с венозной сетью цилиарного тела.
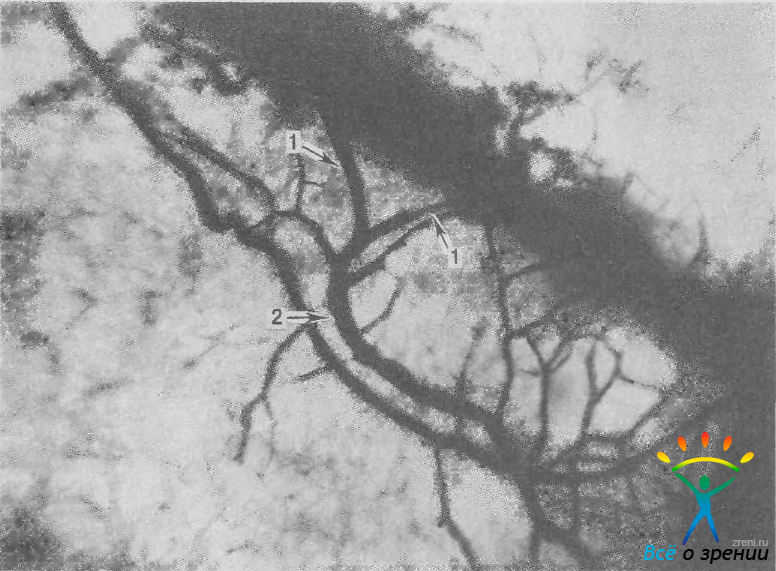
Коллекторные канальцы 2-го типа хорошо видны при биомикроскопии. Они впервые были описаны К. Ascher (1942) и получили название «водяные вены». Эти вены содержат чистую или с примесью крови жидкость. Они появляются в области лимба и идут назад, впадая под острым углом в вены-реципиенты, несущие кровь. Водянистая влага и кровь в этих венах смешиваются не сразу: на некотором протяжении в них можно видеть слой бесцветной жидкости и слой (иногда два слоя по краям) крови. Такие вены получили название «ламинарные». Устья крупных коллекторных канальцев со стороны синуса прикрыты несплошной перегородкой, которая, по-видимому, до некоторой степени предохраняет их от блокады внутренней стенкой шлеммова канала при повышении внутриглазного давления. Выходное отверстие крупных коллекторов имеет овальную форму и диаметр 40—80 мкм.
Эписклеральные и интрасклеральные венозные сплетения связаны между собой анастомозами. Количество таких анастомозов 25— 30, диаметр 30—47 мкм.
↑ Цилиарная мышца
Цилиарная мышца тесно связана с дренажной системой глаза. В мышце различают четыре типа мышечных волокон:
- меридиональные (мышца Брюкке),
- радиальные, или косые (мышца Иванова),
- циркулярные (мышца Мюллера)
- и иридальные волокна (мышца Калазанса).
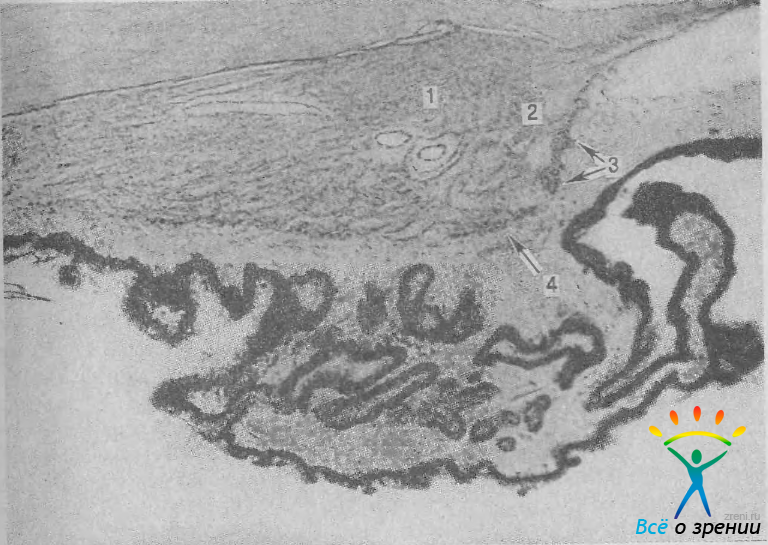
Рис. 10. Мышцы ресничного тела. 1 — меридиональная; 2 — радиальная; 3 — иридальная; 4 — циркулярная. Ув. 35.
Радиальная мышца имеет менее правильное и более рыхлое строение. Ее волокна свободно лежат в строме цилиарного тела, расходясь веером от угла передней камеры к цилиарным отросткам. Часть радиальных волокон начинается от увеальной трабекулы.
Циркулярная мышца состоит из отдельных пучков волокон, расположенных в передневнутреннем отделе цилиарного тела. Существование этой мышцы в настоящее время подвергается сомнению, Ее можно рассматривать как часть радиальной мышцы, волокна которой расположены не только радиально, но и частично циркулярно.
Иридальная мышца расположена у места соединения радужки и цилиарного тела. Она представлена тонким пучком мышечных волокон, идущих к корню радужки. Все части цилиарной мышцы имеют двойную — парасимпатическую и симпатическую — иннервацию.
Сокращение продольных волокон цилиарной мышцы приводит к растяжению трабекулярной мембраны и расширению шлеммова канала. Радиальные волокна оказывают аналогичное, но, по-видимому, более слабое воздействие на дренажную систему глаза.
↑ Варианты строения дренажной системы глаза
Иридокорнеальный угол у взрослого человека имеет выраженные индивидуальные особенности строения [Нестеров А. П., Батманов Ю. Е., 1971]. Мы классифицируем угол не только как общепринято, по ширине входа в него, но и по форме его вершины и конфигурации бухты. Вершина угла может быть острой, средней и тупой. Острая вершина наблюдается при переднем расположении корня радужки (рис. 11).
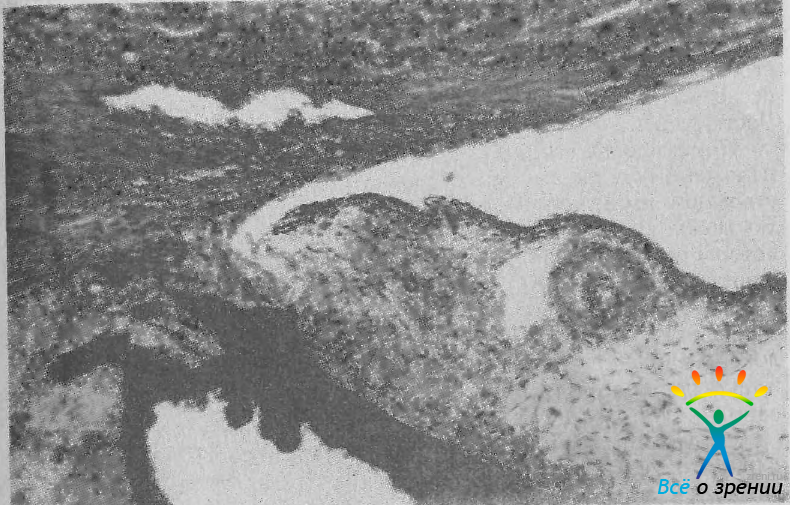
Рис. 11. УПК с острой вершиной и задним положением шлеммова канала. Ув. 90.
В таких глазах полоса ресничного тела, разделяющая радужку и корнеосклеральную сторону угла, очень узкая. Тупая вершина угла отмечается при заднем соединении корня радужки с цилиарным телом (рис. 12).
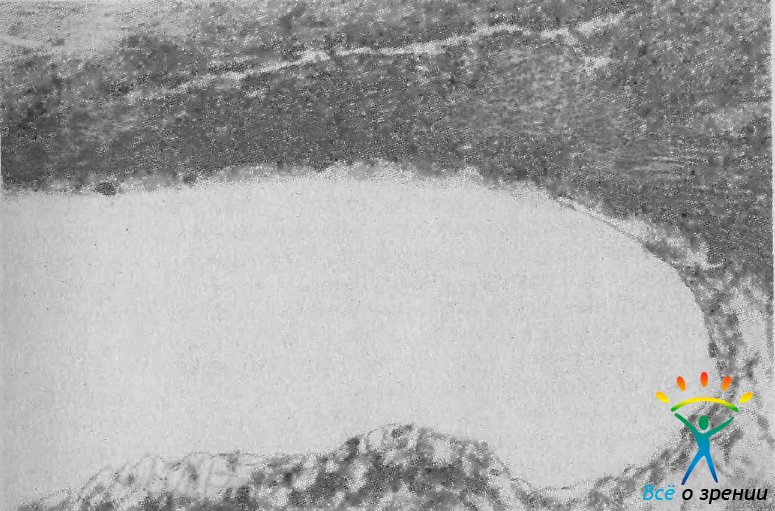
Рис. 12. Тупая вершина УПК и среднее положение шлеммова канала. Ув. 200.
При этом передняя поверхность последнею имеет вид широкой полосы. Средняя вершина угла занимает промежуточное положение между острой и тупой.
Конфигурация бухты угла на разрезе может быть ровной и колбовидной. При ровной конфигурации передняя поверхность радужки постепенно переходит в цилиарное тело (см. рис. 12). Колбовидная конфигурация наблюдается в тех случаях, когда корень радужки образует довольно длинный тонкий перешеек.
При острой вершине угла корень радужки смещен кпереди. Это облегчает образование всех разновидностей закрытоугольной глаукомы, особенно так называемой глаукомы с плоской радужкой. При колбовидной конфигурации бухты угла та часть корня радужки, которая прилежит к цилиарному телу, особенно тонкая. В случае повышения давления в задней камере эта часть резко выпячивается кпереди. В некоторых глазах задняя стенка бухты угла частично образована цилиарным телом. Его передняя часть при этом отходит от склеры, поворачивается внутрь глаза и располагается в одной плоскости с радужкой (рис. 13).
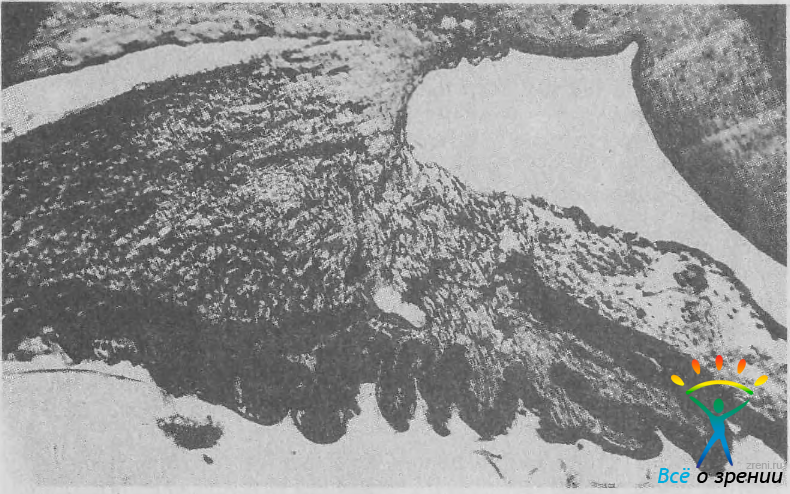
Рис. 13. УПК, задняя стенка которого сформирована короной ресничного тела. Ув. 35.
В таких случаях при выполнении антиглаукоматозных операций с иридэктомией можно повредить цилиарное тело, вызвав сильное кровотечение.
Можно выделить три варианта расположения заднего края шлеммова канала относительно вершины угла передней камеры: переднее, среднее и заднее. При переднем расположении (41% наблюдений) часть бухты угла находится позади синуса (рис. 14).
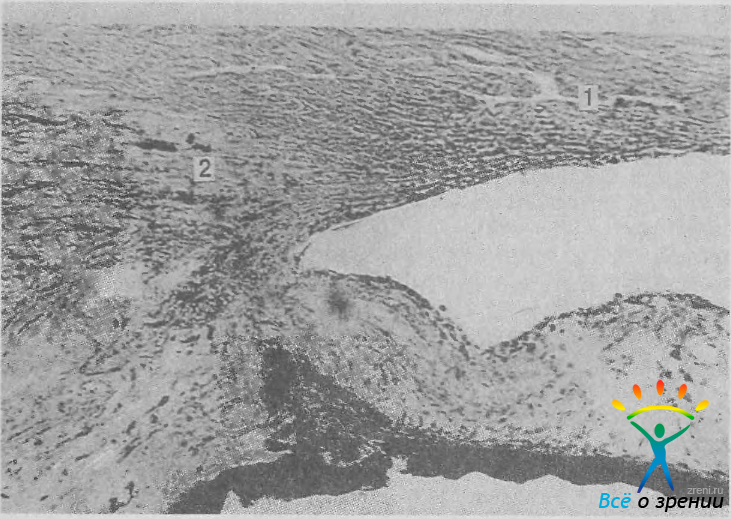
Рис. 14. Переднее положение шлеммова канала (1). Меридиональная мышца (2) начинается в склере на значительном расстоянии от канала. Ув. 86.
Среднее расположение (40% наблюдений) характеризуется тем, что задний край синуса совпадает с вершиной угла (см. рис. 12). Оно по существу является вариантом переднего расположения, так как весь шлеммов канал граничит с передней камерой. При заднем расположении канала (19% наблюдений) часть его (иногда до 1/2 ширины) выходит за пределы бухты угла в область, пограничную с цилиарным телом (см. рис. 11).
Угол наклона просвета шлеммова канала к передней камере, точнее к внутренней поверхности трабекулы, варьирует от 0 до 35°, чаще всего он равен 10—15°.
Степень развития склеральной шпоры индивидуально широко варьирует. Она может закрыть почти половину просвета шлеммова канала (см. рис. 4), однако в некоторых глазах шпора короткая или совсем отсутствует (см. рис. 14).
↑ Гониоскопическая анатомия иридокорнеального угла
Индивидуальные особенности строения УПК могут быть изучены в клинических условиях с помощью гониоскопии. Основные структуры УПК представлены на рис. 15.
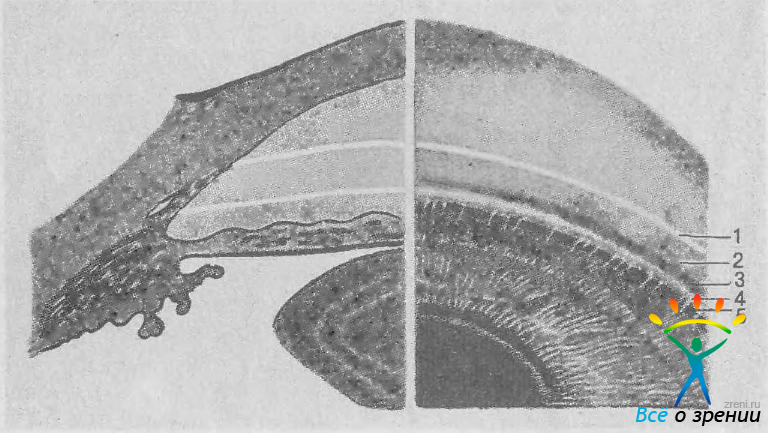
Рис. 15. Структуры УПК. 1 — переднее пограничное кольцо Швальбе; 2 — трабекула; 3 — шлеммов канал; 4 — склеральная шпора; 5 — ресничное тело.
В типичных случаях кольцо Швальбе видно как слегка проминирующая сероватая непрозрачная линия на границе между роговицей и склерой. При осмотре со щелью на этой линии сходятся два луча световой вилки от передней и задней поверхностей роговицы. Кзади от кольца Швальбе имеется незначительное углубление — инцизура, в которой нередко видны осевшие там гранулы пигмента, особенно заметные в нижнем сегменте. У некоторых людей кольцо Швальбе проминирует кзади весьма значительно и смещено кпереди (задний эмбриотоксон). В таких случаях его можно видеть при биомикроскопии без гониоскопа.
Трабекулярная мембрана натянута между кольцом Швальбе спереди и склеральной шпорой сзади. При гониоскопии она выявляется как шероховатая полоска сероватого цвета. У детей трабекула полупрозрачная, с возрастом ее прозрачность уменьшается и трабекулярная ткань выглядит более плотной. К возрастным изменениям относится также отложение в трабекулярном переплете гранул пигмента, а иногда и эксфолиативных чешуек. В большинстве случаев пигментируется только задняя половина трабекулярного кольца. Значительно реже пигмент откладывается и в недеятельной части трабекулы и даже в склеральной шпоре. Ширина видимой при гониоскопии части трабекулярной полосы зависит от угла зрения: чем уже УПК, тем под более острым углом видны его структуры и тем уже они кажутся наблюдателю.
Склеральный синус отделен от передней камеры задней половиной трабекулярной полосы. Самая задняя часть синуса часто заходит за склеральную шпору. При гониоскопии синус виден только в тех случаях, когда он заполняется кровью, и только в тех глазах, в которых пигментация трабекулы отсутствует или выражена слабо. В здоровых глазах синус заполняется кровью значительно легче, чем в глаукоматозных.
Расположенная кзади от трабекулы склеральная шпора имеет вид узкой беловатой полоски. Ее трудно идентифицировать в глазах с обильной пигментацией или развитой увеальной структурой в вершине УПК.
В вершине УПК в виде полосы разной ширины расположено ресничное тело, точнее его передняя поверхность. Цвет этой полосы варьирует от светло-серого до темно-коричневого в зависимости от цвета глаз. Ширина полосы ресничного тела определяется местом прикрепления к нему радужки: чем дальше кзади радужка соединяется с ресничным телом, тем шире видимая при гониоскопии полоса. При заднем прикреплении радужки вершина угла тупая (см. рис. 12), при переднем — острая (см. рис. 11). При чрезмерно переднем прикреплении радужки цилиарное тело не видно при гониоскопии и корень радужки начинается на уровне склеральной шпоры или даже трабекулы.
Строма радужки образует складки, из которых самая периферическая, часто называемая складкой Фукса, расположена напротив кольца Швальбе. Расстояние между этими структурами определяет ширину входа (апертуру) в бухту УПК. Между складкой Фукса и цилиарным телом расположен корень радужки. Это ее самая тонкая часть, которая может смещаться кпереди, вызывая сужение УПК, или кзади, приводя к его расширению, в зависимости от соотношения давлений в передней и задней камерах глаза. Нередко от стромы корня радужки отходят отростки в виде тонких нитей, тяжей или нешироких листков. В одних случаях они, огибая вершину УПК, переходят на склеральную шпору и образуют увеальную трабекулу, в других — пересекают бухту угла, прикрепляясь к его передней стенке: к склеральной шпоре, трабекуле или даже к кольцу Швальбе (отростки радужки, или гребенчатая связка). Следует отметить, что у новорожденных увеальная ткань в УПК значительно выражена, но с возрастом атрофируется, и у взрослых при гониоскопии ее обнаруживают редко. Отростки радужки не следует путать с гониосинехиями, которые выглядят более грубыми и отличаются беспорядочностью расположения.
В корне радужки и увеальной ткани в вершине УПК иногда видны тонкие сосуды, расположенные радиально или циркулярно. В таких случаях обычно обнаруживают гипоплазию или атрофию стромы радужки.
В клинической практике важное значение придают конфигурации, ширине и пигментации УПК. На конфигурацию бухты УПК существенное влияние оказывает положение корня радужки между передней и задней камерами глаза. Корень может быть плоским, выпяченным кпереди или запавшим кзади. В первом случае давление в переднем и заднем отделах глаза одинаковое или почти одинаковое, во втором — выше давление в заднем отделе, в третьем — в передней камере глаза. Выпячивание кпереди всей радужки указывает на состояние относительного зрачкового блока с повышением давления в задней камере глаза. Выпячивание только корня радужки свидетельствует об его атрофии или гипоплазии. На фоне общего бомбажа корня радужки можно видеть очаговые выпячивания ткани, напоминающие кочки. Эти выпячивания связаны с мелкоочаговой атрофией стромы радужки. Причина западения корня радужки, которое наблюдается в некоторых глазах, не вполне ясна. Можно думать или о более высоком давлении в переднем отделе глаза по сравнению с задним, или о некоторых анатомических особенностях, создающих впечатление западения корня радужки.
Ширина УПК зависит от расстояния между кольцом Швальбе и радужкой, ее конфигурации и места прикрепления радужки к ресничному телу. Приведенная ниже классификация ширины У ПК составлена с учетом видимых при гониоскопии зон угла и ориентировочной его оценки в градусах (табл. 1).

Таблица 1. Гониоскопическая классификация ширины УПК
При широком УПК можно видеть все его структуры, при закрытом — только кольцо Швальбе и иногда переднюю часть трабекулы. Правильно оценить ширину УПК при гониоскопии можно только в том случае, если больной смотрит прямо перед собой. Изменяя положение глаза или наклон гониоскопа, можно увидеть все структуры даже при узком УПК.
Ширину УПК можно ориентировочно оценить и без гониоскопа. Узкий луч света от щелевой лампы направляют на радужку через периферическую часть роговицы как можно ближе к лимбу. Сопоставляют толщину среза роговицы и ширину входа в УПК, т. е. определяют расстояние между задней поверхностью роговицы и радужкой. При широком УПК это расстояние примерно равно толщине среза роговицы, среднешироком — 1/2 толщины среза, узком — 1/4 толщины роговицы и щелевидном — менее 1/4 толщины роговичного среза. Этот способ [Herrick W., 1969] позволяет оценить ширину УПК только в носовом и височном сегментах. Следует иметь в виду, что вверху УПК несколько уже, а внизу — шире, чем в боковых отделах глаза.
Наиболее простой тест для оценки ширины УПК предложен М. В. Вургафтом и соавт. (1973). Он основан на феномене полного внутреннего отражения света роговицей. Источник света (настольная лампа, фонарик и т. п.) помещают с наружной стороны от исследуемого глаза: сначала на уровне роговицы, а затем медленно смещают кзади. В определенный момент, когда лучи света попадают на внутреннюю поверхность роговицы под критическим углом, с носовой стороны глаза в зоне склерального лимба появляется яркое световое пятно. Широкое пятно — диаметром 1,5—2 мм — соответствует широкому, а диаметром 0,5—1 мм — узкому УПК. Нерезкое свечение лимба, появляющееся только при повороте глаза кнутри, характерно для щелевидного УПК. При закрытом иридокорнеальном угле свечение лимба вызвать не удается.
Узкий и особенно щелевидный УПК предрасположен к блокаде его корнем радужки при возникновении зрачкового блока или расширении зрачка. Закрытый угол свидетельствует об уже существующей блокаде. Для того чтсбы дифференцировать функциональный блок угла от органического, на роговицу надавливают гониоскопом без гаптической части [Forbes М., 1966]. При этом жидкость из центрального отдела передней камеры смещается к периферии, и при функциональной блокаде угол открывается. Обнаружение узких или широких спаек в УПК свидетельствует о его частичной органической блокаде.
Трабекула и прилегающие к ней структуры нередко приобретают темную окраску вследствие оседания в них пигментных гранул, поступающих в водянистую влагу при распаде пигментного эпителия радужки и ресничного тела. Степень пигментации принято оценивать в баллах от 0 до 4. Отсутствие пигмента в трабекуле обозначают цифрой 0, слабую пигментацию ее задней части — 1, интенсивную пигментацию той же части — 2, интенсивную пигментацию всей трабекулярной зоны — 3 и всех структур передней стенки УПК — 4. В здоровых глазах пигментация трабекул появляется только в среднем или пожилом возрасте и выраженность ее по приведенной выше шкале оценивается в 1—2 балла. Более интенсивная пигментация структур УПК свидетельствует с патологии.
↑ Отток водянистой влаги из глаза
Различают основной и дополнительный (увеосклеральный) пути оттока. Согласно некоторым расчетам, по основному пути оттекает примерно 85—95% водянистой влаги, по увеосклеральному — 5— 15% [BillA., Phillips С., 1971]. Основной отток проходит через трабекулярную систему, шлеммов канал и его выпускники.
Трабекулярный аппарат представляет собой многослойный, самоочищающийся фильтр, обеспечивающий одностороннее движение жидкости и мелких частиц из передней камеры в склеральный синус. Сопротивление движению жидкости в трабекулярной системе в здоровых глазах в основном обусловливают индивидуальный уровень ВГД и его относительное постоянство.
В трабекулярном аппарате выделяют четыре анатомических слоя. Первый из них, увеальная трабекула, можно сравнить с решетом, которое не препятствует движению жидкости. Корнеосклеральная трабекула имеет более сложное строение. Она состоит из нескольких «этажей» — узких щелей, разделенных прослойками волокнистой ткани и отростками эндотелиальных клеток на многочисленные отсеки. Отверстия в трабекулярных пластинах не совпадают друг с другом. Движение жидкости осуществляется в двух направлениях: в поперечном, через отверстия в пластинах, и продольном, по межтрабекулярным щелям. Учитывая особенности архитектоники трабекулярной сети и сложный характер движения в ней жидкости, можно предположить, что часть сопротивления оттоку водянистой влаги локализуется в корнеосклеральной трабекуле.
В юкстаканаликулярной ткани нет явных, оформленных путей оттока. Все же, по данным J. Rohen (1986), влага через этот слой движется по определенным маршрутам, отграниченным менее проницаемыми участками ткани, содержащей гликозаминогликаны. Полагают, что основная часть сопротивления оттоку в нормальных глазах локализуется в юкстаканаликулярном слое трабекулярной диафрагмы.
Четвертый функциональный слой трабекулярной диафрагмы представлен непрерывным слоем эндотелия. Отток сквозь этот слой происходит в основном через динамические поры или гигантские вакуоли. В связи с их значительным количеством и размерами сопротивление оттоку здесь небольшое; по данным A. Bill (1978), не более 10% от его общей величины.
Трабекулярные пластины связаны с продольными волокнами ресничкой мышцы и через увеальную трабекулу с корнем радужки. В нормальных условиях тонус ресничной мышцы непрерывно изменяется. Это сопровождается колебаниями в натяжении трабекулярных пластин. В результате этого трабекулярные щели попеременно расширяются и спадаются, что способствует движению жидкости внутри трабекулярной системы, ее постоянному перемешиванию и обновлению. Аналогичное, но более слабое влияние на трабекулярные структуры оказывают колебания тонуса зрачковых мышц. Колебательные движения зрачка препятствуют застою влаги в криптах радужки и облегчают отток из нее венозной крови.
Непрерывные колебания тонуса трабекулярных пластин играют важную роль в сохранении их эластичности и упругости. Можно предположить, что прекращение колебательных движений трабекулярного аппарата приводит к огрублению волокнистых структур, перерождению эластических волокон и в конечном счете к ухудшению оттока водянистой влаги из глаза.
Движение жидкости через трабекулы выполняет еще одну важную функцию: промывание, очистку трабекулярного фильтра. В трабекулярную сеть поступают продукты распада клеток и пигментные частицы, которые удаляются с током водянистой влага. Трабекулярный аппарат отделен от склерального синуса тонким слоем ткани (юкстаканаликулярная ткань), содержащим волокнистые структуры и фиброциты. Последние непрерывно продуцируют, с одной стороны, мукополисахариды, а с другой — ферменты, деполимеризующие их. После деполимеризации остатки мукополисахаридов вымываются водянистой влагой в просвет склерального синуса.
Промывная функция водянистой влаги хорошо изучена в экспериментах. Ее эффективность пропорциональна минутному объему жидкости, фильтрующейся через трабекулу, и, следовательно, зависит от интенсивности секреторной функции ресничного тела.
Установлено, что мелкие частицы, размером до 2—3 мкм, задерживаются в трабекулярной сети частично, а более крупные — полностью. Интересно, что нормальные эритроциты, диаметр которых 7—8 мкм, проходят через трабекулярный фильтр довольно свободно [Bill А., 1968]. Это связано с эластичностью эритроцитов и их способностью проходить через поры диаметром 2—2,5 мкм. Вместе с тем измененные и потерявшие эластичность эритроциты задерживаются трабекулярным фильтром.
Очищение трабекулярного фильтра от крупных частиц происходит путем фагоцитоза. Фагоцитарная активность характерна для клеток трабекулярного эндотелия. Состояние гипоксии, которое возникает при нарушении оттока водянистой влаги через трабекулу в условиях понижения ее продукции, приводит к уменьшению активности фагоцитарного механизма очистки трабекулярного фильтра.
Способность трабекулярного фильтра к самоочищению уменьшается в пожилом возрасте из-за снижения скорости продуцирования водянистой влаги и дистрофических изменений в трабекулярной ткани. Следует иметь в виду, что трабекулы не имеют кровеносных сосудов и получают питание из водянистой влаги, поэтому даже частичное нарушение ее циркуляции отражается на состоянии трабекулярной диафрагмы.
Клапанная функция трабекулярной системы, пропускающей жидкость и частицы только в направлении из глаза в склеральный синус, связана прежде всего с динамическим характером пор в эндотелии синуса. Если давление в синусе выше, чем в передней камере, то гигантские вакуоли не формируются и внутриклеточные поры закрываются. Одновременно наружные слои трабекулы смещаются кнутри. При этом сдавливаются юкстаканаликулярная ткань и межтрабекулярные щели. Синус нередко заполняется кровью, но ни плазма, ни эритроциты не проходят в глаз, если не поврежден эндотелий внутренней стенки синуса.
Склеральный синус в живом глазу представляет собой очень узкую щель, движение жидкости по которой связано со значительной затратой энергии. Вследствие этого водянистая влага, поступающая в синус через трабекулу, течет по его просвету только до ближайшего коллекторного канала. При повышении ВГД просвет синуса суживается и сопротивление оттока по нему увеличивается. В связи с большим количеством коллекторных канальцев сопротивление оттоку в них невелико и более стабильно, чем в трабекулярном аппарате и синусе.
↑ Отток водянистой влаги и закон Пуазейля
Дренажный аппарат глаза можно рассматривать как систему, состоящую из канальцев и пор. Ламинарное движение жидкости в такой системе подчиняется закону Пуазейля. В соответствии с этим законом объемная скорость движения жидкости прямо пропорциональна разности давлений в начальном и конечном пунктах движения. Закон Пуазейля положен в основу многих исследований по гидродинамике глаза. На этом законе основаны, в частности, все тонографические расчеты. Между тем в настоящее время накопилось много данных, свидетельствующих о том, что с повышением внутриглазного давления минутный объем водянистой влаги увеличивается в значительно меньшей мере, чем это следует из закона Пуазейля. Этот феномен можно объяснить деформацией просветов шлеммова канала и трабекулярных щелей при повышении офтальмотонуса. Результаты исследований на изолированных глазах человека с перфузией шлеммова канала тушью показали, что ширина его просвета прогрессивно уменьшается при увеличении внутриглазного давления [Нестеров А. П., Батманов Ю. Е., 1978]. При этом синус сдавливается сначала только в переднем отделе, а затем происходит очаговое, пятнистое сдавление просвета канала и в Других частях канала. При повышении офтальмотонуса до 70 мм рт. ст. открытой остается узкая полоска синуса в самом заднем его отделе, защищенном от сдавления склеральной шпорой.
При кратковременном повышении внутриглазного давления трабекулярный аппарат, смещаясь кнаружи в просвет синуса, растягивается и его проницаемость увеличивается. Однако результаты проведенных нами исследований показали, что если высокий уровень офтальмотонуса поддерживать в течение нескольких часов, то возникает прогрессирующее сдавление трабекулярных щелей: сначала в зоне, прилежащей к шлеммову каналу, а затем и в остальных отделах корнеосклеральной трабекулы.
↑ Увеосклеральный отток
Кроме фильтрации жидкости по дренажной системе глаза, у обезьян и человека отчасти сохранился и более древний путь оттока — через передний отдел сосудистого тракта (рис. 16).
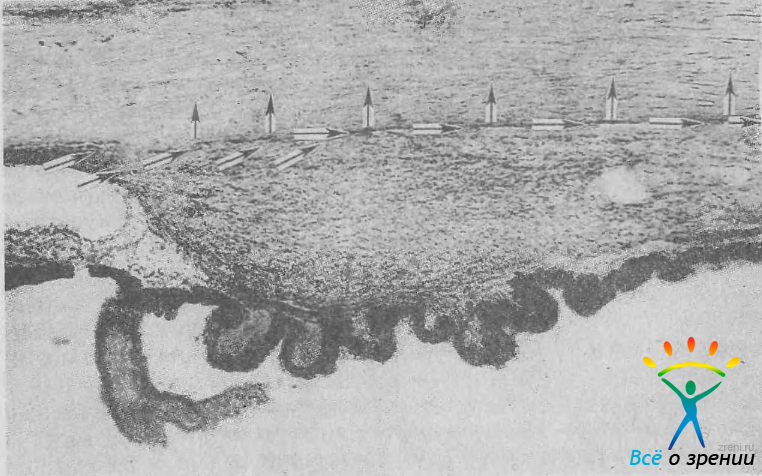
Рис. 16. УПК и ресничное тело. Стрелками показан увеосклеральный путь оттока водянистой влаги. Ув. 36.
Увеальный (или увеосклеральный) отток осуществляется из угла передней камеры через передний отдел ресничного тела вдоль волокон мышцы Брюкке в супрахориоидальное пространство [Bill А., 1966]. Из последнего жидкость оттекает по эмиссариям и прямо через склеру или всасывается в венозное отделы капилляров сосудистой оболочки [Pederson J. et al., 1977].
Проведенные в нашей лаборатории исследования [Черкасова И. Н., Нестеров А. П., 1976] показали следующее. Увеальный отток функционирует при условии, что давление в передней камере превышает давление в супрахориоидальном пространстве не менее чем на 2 мм рт. ст. В супрахориоидальном пространстве отмечается значительное сопротивление движению жидкости, особенно в меридиональном направлении. Склера проницаема для жидкости. Отток через нее подчиняется закону Пуазейля, т. е. пропорционален величине фильтрующего давления. При давлении 20 мм рт.ст. через 1 см2 склеры фильтруется в среднем 0,07 мм3 жидкости в 1 мин. При истончении склеры отток через нее пропорционально увеличивается. Таким образом, каждый отдел увеосклерального пути оттока (увеальный, супрахориоидальный и склеральный) оказывает сопротивление оттоку водянистой влаги. Повышение офтальмотонуса не сопровождается усилением увеального оттока, так как на ту же величину повышается и давление в супрахориоидальном пространстве, которое к тому же суживается. Миотики уменьшают увеосклеральный отток, а циклоплегические препараты увеличивают его. По данным A. Bill и С. Phillips (1971), у человека по увеосклеральному пути оттекает от 4 до 27% водянистой влаги.
Индивидуальные различия в интенсивности увеосклерального оттока, по-видимому, весьма значительны. Они зависят от индивидуальных анатомических особенностей и возраста. Van der Zippen (1970) обнаружил у детей открытые пространства вокруг пучков ресничной мышцы. С возрастом эти пространства заполняются соединительной тканью. При сокращении ресничной мышцы свободные пространства сдавливаются, а при ее расслаблении расширяются.
По нашим наблюдениям, увеосклеральный отток не функционирует при остром приступе глаукомы и злокачественной глаукоме. Это объясняется блокадой УПК корнем радужки и резким повышением давления в заднем отделе глаза.
Увеосклеральный отток играет, по-видимому, некоторую роль в развитии цилиохориоидальной отслойки. Как известно, увеальная тканевая жидкость содержит значительное количество белка из-за высокой проницаемости капилляров ресничного тела и хориоидеи [Bill А., 1987]. Коллоидно-осмотическое давление плазмы крови равно примерно 25 мм рт.ст., увеальной жидкости — 16 мм рт.ст., а величина этого показателя для водянистой влаги близка к нулю. Вместе с тем разность гидростатического давления в передней камере и супрахориоидее не превышает 2 мм рт.ст. Следовательно, главной движущей силой оттока водянистой влаги из передней камеры в супрахориоидею служит разность не гидростатического, а коллоидно-осмотического давления. Коллоидно-осмотическое давление плазмы крови также является причиной всасывания увеальной жидкости в венозные отделы сосудистой сети ресничного тела и хориоидеи. Гипотония глаза, чем бы она не была вызвана, приводит к расширению увеальных капилляров и повышению их проницаемости. Концентрация белка, а следовательно, и коллоидно-осмотическое Давление плазмы крови и увеальной жидкости становятся примерно Равными. В результате этого усиливается всасывание водянистой влаги из передней камеры в супрахориоидею, а ультрафильтрация Увеальной жидкости в сосудистую сеть прекращается. Ретенция Увеальной тканевой жидкости приводит к отслойке цилиарного тела хориоидеи, прекращению секреции водянистой влаги.
↑ Регуляция продукции и оттока водянистой влаги
Скорость образования водянистой влаги регулируется как пассивными, так и активными механизмами. При повышении ВГД суживаются увеальные сосуды, уменьшается кровоток и давление фильтрации в капиллярах цилиарного тела. Снижение ВГД приводит к противоположным эффектам. Изменения увеального кровотока при колебаниях ВГД в определенной мере полезны, так как способствуют поддержанию стабильного ВГД.
Есть основания думать, что на активную регуляцию продукции водянистой влаги влияет гипоталамус. Как функциональные, так и органические гипоталамические нарушения часто ассоциируются с повышенной амплитудой суточных колебаний ВГД и гиперсекрецией внутриглазной жидкости [Бунин А. Я., 1971].
Пассивная и активная регуляция оттока жидкости из глаза отчасти рассмотрена выше. Основное значение в механизмах регуляции оттока имеет ресничная мышца. По нашему мнению, определенную роль играет также радужка. Корень радужки связан с передней поверхностью ресничного тела и увеальной трабекулой. При сужении зрачка корень радужки, а вместе с ним и трабекула натягиваются, трабекулярная диафрагма отходит кнутри, а трабекулярные щели и шлеммов канал расширяются. Аналогичный эффект дает сокращение дилататора зрачка. Волокна этой мышцы не только расширяют зрачок, но и натягивают корень радужки. Эффект натяжения корня радужки и трабекулы особенно выражен в тех случаях, когда зрачок ригиден или фиксирован миотиками. Это позволяет объяснить положительное действие на отток водянистой влаги ?-адреноагонистов и особенно комбинации их (например, адреналина) с миотиками.
Изменение глубины передней камеры также оказывает регулирующее влияние на отток водянистой влаги. Как показали перфузионные опыты, углубление камеры приводит к немедленному усилению оттока, а ее обмеление — к его задержке [Grant М., 1963]. Мы пришли к такому же выводу, изучая на нормальных и глаукоматозных глазах изменения оттока под влиянием передней, боковой и задней компрессии глазного яблока [Нестеров А. П. и др., 1974]. При передней компрессии через роговицу радужка и хрусталик отдавливались кзади и отток влаги увеличивался в среднем в 1,5 раза по сравнению с его величиной при боковой компрессии той же силы. Задняя компрессия приводила к смещению иридохрусталиковой диафрагмы кпереди, и показатель оттока при этом снижался в 1,2—1,5 раза. Влияние изменений положения иридохрусталиковой диафрагмы на отток можно объяснить только механическим действием натяжения корня радужки и цинновых связок на трабекулярный аппарат глаза. Поскольку при усилении продукции влаги передняя камера углубляется, этот феномен способствует поддержанию стабильного ВГД.
----
Статья из книги: Глаукома. Нестеров А.П.

Комментариев 0