Нуклеотидрегулируемые каналы сенсорных клеток (ЧАСТЬ ІІІ)


Содержание:
Описание
↑ Влияние реагентов, модифицирующих SH-группы, на фототок в зрительной клетке и цГМФ-регулируемые ионные каналы
Как отмечалось выше, сульфгидрильные группы родопсина функционально важны для активации трансдуцина и, следовательно, непосредственно участвуют в цикле фототрансдукции. Если это так, то их модификация должна приводить к изменениям фототока, регистрируемого в сетчатке или изолированной клетке. В настоящем разделе нами изложены эксперименты, подтверждающие это предположение. Однако оказалось, что при изменении фототока, вызванного модификацией SH-групп, обнаруживается компонент, который никак не может быть объяснен ингибированием или активацией ферментов, участвующих в усилении сигнала. Попытка объяснить этот феномен привела нас к исследованию ионных каналов в плазматической мембране клетки. В соответствующем разделе показано, что эти каналы также содержат функционально важные SH-группы, модификация которых может приводить к активации каналов и в отсутствие цГМФ.
Для исследования действия модификации SH-групп на ток изолированной палочки саламандры использовался N-этилмалеимид (НЭМ), иодацетамид (ИАА) и р-гидроксимеркурийбензоат (ПГМБ), которые добавляли в раствор Рингера, омывающий наружный сегмент палочки. Ток изолированной палочки саламандры регистрировался методом всасывающего электрода.
На рис. 4.5, А представлено изменение темнового тока и ответа палочки на свет, вызванное добавлением в проток 0,5 мМ НЭМ.
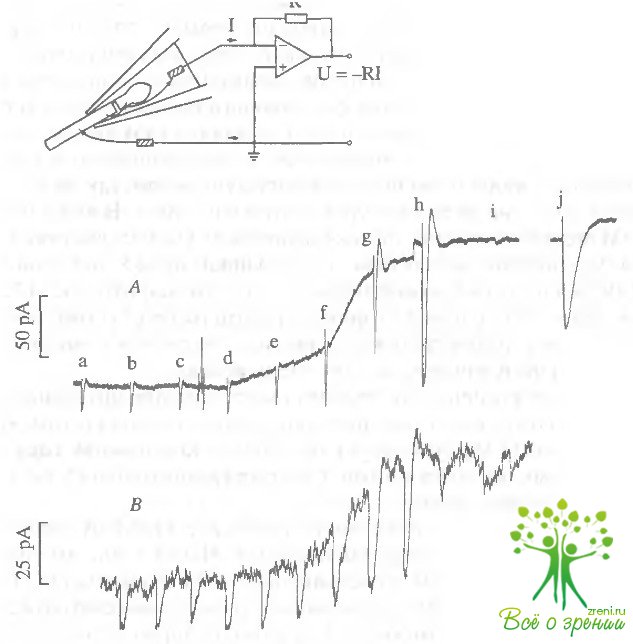
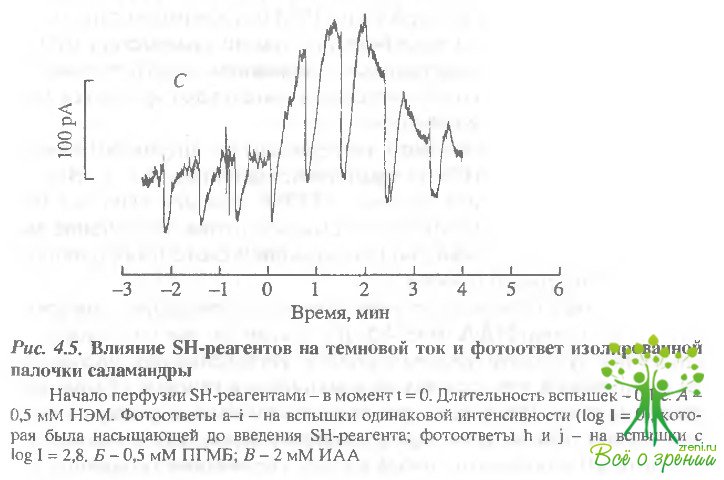
Под действием НЭМ темновой ток палочки начинает возрастать и выходит на новый уровень. Стрелками указаны моменты включения коротких вспышек света. Интенсивность вспышек a-i была одинаковой (log I = 0). Эта интенсивность была насыщающей для фотоответа палочки до воздействия НЭМ (ответы a-d), т.е. вспышки a-d вызывали полное перекрытие тока палочки, и их пики показывают уровень нулевого тока, а их амплитуда - величину темнового тока палочки, омываемой обычным раствором Рингера без добавления фармакологических агентов. В первые несколько минут после начала действия НЭМ амплитуда ответов, вызываемых вспышками этой же интенсивности (f, g), увеличивалась, однако, несмотря на большую амплитуду, пик фотоответа g уже не достигает уровня нулевого тока. В присутствии НЭМ полного перекрытия тока палочки не удалось достичь даже при увеличении интенсивности вспышки на 2,8 log единицы (вспышка h), хотя пиковое значение тока этого фотоответа близко к нулю. Это, с одной стороны, свидетельствует о том, что относительная чувствительность палочки (часть тока, выключаемая одной фотоизомеризацией) уменьшилась.
С другой стороны, то, что свет очень большой интенсивности не перекрывает всего тока палочки, свидетельствует о том, что в присутствии НЭМ существует остаточная компонента тока, которая не выключается светом. Она составляет около 15 пА в нескольких экспериментах.
При дальнейшей инкубации палочки с НЭМ чувствительность продолжала быстро уменьшаться. Через 5 мин амплитуда фотоответа, вызываемого вспышкой с интенсивностью log 1 = О, значительно уменьшилась (фотоответ i). Величина светочувствительного тока (см. фотоответ j на насыщающую вспышку света с интенсивностью log I = 2,8) несколько уменьшается по сравнению с первыми минутами действия НЭМ (см. фотоответ h), но остается примерно в три раза больше, чем до воздействия НЭМ. Однако в связи со значительным снижением чувствительности для выключения всего светочувствительного тока требуется свет гораздо большей интенсивности.
Результаты аналогичного эксперимента с другим SH-связывающим реагентом, ПГМБ, представлены на рис. 4,5, Б. В этих экспериментах, как и в опытах с НЭМ, при действии 0,5 мМ ПГМБ наблюдалось увеличение темнового тока, увеличение амплитуды ответа на насыщающую вспышку света и постепенное снижение чувствительности.
Несколько отличная от этой картина наблюдалась при добавлении в раствор ИАА (рис. 4.5, В). В этом случае не наблюдали снижения чувствительности палочки. Интенсивность насыщающей вспышки в этих опытах не изменялась в течение 12 мин перфузии палочки ИАА даже при повышении концентрации реагента до 2 мМ. Кроме того, при добавлении ИАА, как и в опытах с другими SH-реагентами, наблюдалось увеличение темнового тока, однако это было кратковременной реакцией на добавление реагента, и уровень темнового тока возвращался к первоначальному через несколько минут.
Таким образом, все исследованные SH-реагенты вызывали
- увеличение темнового тока палочки,
- увеличение амплитуды фотоответа на насыщающую вспышку света на величину, коррелирующую с увеличением темнового тока, однако во всех случаях меньшую, чем увеличение темнового тока,
- возникновение (во всех случаях) нечувствительной к свету компоненты темнового тока, которая сохранялась даже после насыщающей вспышки света. Кроме того, два из исследованных реагентов, НЭМ и ГТГМБ, вызывали снижение чувствительности палочки. Из этих результатов следует, что SH-реагенты влияют, по крайней мере, на два параметра фсторецепторной клетки: во-первых, повышают проводимость каналов в плазматической мембране и, во-вторых, ингибируют цикл фототрансдукции в палочке.
Под действием НЭМ фотоответ интактных палочек через несколько минут замедляется, а кривая ответ-интенсивность становится круче. Аналогичные изменения вызывают ингибиторы ФДЭ. Отсюда возникло предположение, что вызванное SH-реагентами изменение чувствительности палочки связано с ингибирующим действием SH-реагентов на ФДЭ. При этом следует заметить, что ИАА не оказывал заметного влияния на чувствительность палочки, поэтому интересно сравнить его влияние на активность ФДЭ с действием НЭМ.
Мы оценивали фосфсдиэстеразную активность в суспензии бычьих НСП, контролируя изменения рН, вызванные гидролизом цГМФ. На рис. 4.6, а и рис. 4.6, б представлены изменения фосфодиэстеразной активности в суспензии НСП, вызванные добавками ОД мМ НЭМ и 0,1 мМ ПГМБ соответственно.
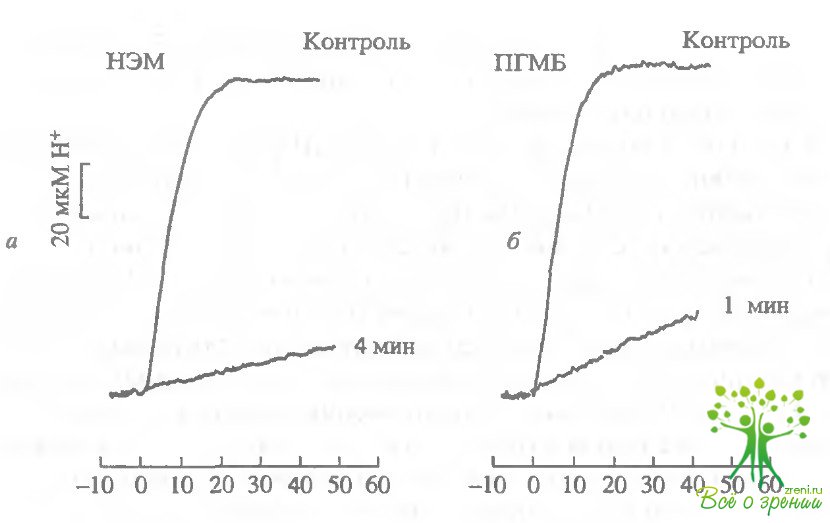
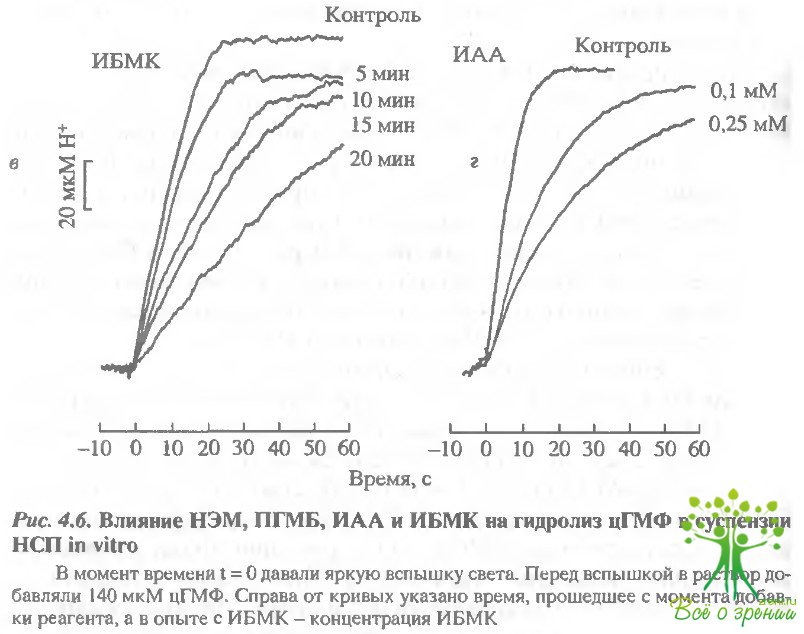
НЭМ и ПГМБ быстро и эффективно блокируют фосфодиэстеразную активность в суспензии НСП. Для сравнения были проведены аналогичные эксперименты с широко применяемым известным ингибитором ФДЭ изобутилметилксантином (ИБМК) в концентрации 0,1 и 0,25 мМ. Такие концентрации ИБМК вызывали существенные изменения в электрофизиологии палочек. Результаты такого эксперимента представле ны на рис. 4.6, г. НЭМ и ПГМБ блокируют фосфодиэстеразную активность в суспензии НСП эффективнее, чем ИБМК. ИАА в концентрации 0,2 мМ также оказывал ингибирующее действие на фосфодиэстеразную активность суспензии НСП, однако менее эффективное, чем НЭМ и ПГМБ (см. рис. 4.6, в).
Из описанных выше экспериментов с очевидностью следует, что если плазматическая мембрана проницаема для НЭМ даже в небольшой степени, этою будет вполне достаточно для блокирования цикла фоготрансцукции и снижения чувствительности палочки, поскольку НЭМ является эффективным ингибитором?
ФДЭ. С другой стороны, хотя ингибирующее действие ИАА меньше, чем действие НЭМ, при высокой проницаемости мембраны для ИАА его было бы достаточно для изменения чувствительности палочки.

Однако ИАА не оказывая влияния на чувствительность палочки. Такое несоответствие может быть связано с различием проникающей способности этих реагентов. Кроме того, проницаемость плазматической мембраны для использованных нами реагентов важно знать также и для выяснения локализации функциональных SН-групп. Поэтому мы провели серию экспериментов по исследованию проницаемость плазматической мембраны НСП для НЭМ и ИАА. Изолированные сетчатки лягушки инкубировали с 10 мМ НЭМ или с 10 мМ ИАА а затем оценивали проницаемость мембраны палочки для этих реагентов по степени модификации SН-групп родопсина. Результаты этих экспериментов представлены в таблице.
При инкубации с НЭМ количество немодпфицированных SН-групп через 20 мин падает до 85% от контрольного, а через 120 мин - до 70% Иная картина наблюдается при инкубации сетчаток с ИАА. Через 20 мин инкубации с ИАА количество интактных SН-групп в молекулах родопсина не отличается от контрольного, и только через 120 мин модифицируется около 20% SН-групп
Таким образом, даже при концентрации 10 мМ. в 5 раз превышающей максимальную концентрацию ИАА, применявшуюся в электрофизиологических экспериментах, ИАА проникает в клетку очень медленно. Это означает, что в электрофизиологической части этой работы мы наблюдали действие ИАА на палочку только снаружи, на каналы плазматической мембраны, а не на ферменты цикла трансдукции. Кроме того, это свидетельствует о том, что канальный белок содержит SН-группы не только с цитоплазматической, но и с наружной стороны мембраны и модификация как тех, так и других SH-групп приводит к повышению проводимости каналов.
Естественным объяснением того факта, что при действии SН-модификагоров появляется компонента в фотоответе, которая не может быть подавлена даже при насыщающем ответе, состоит в том, что модификация SН-групп приводит к изменению проводимости цГМФ-регулируемых ионных каналов.
Для исследования действия SН-реагентов на проводимость плазматической мембраны фоторецепторов был применен метод локальной фиксации потенциала на фрагменте мембраны (пэтч- кламп). Мы регистрировали ток фрагмента плазматической мембраны палочки, перфузируемого с внутриклеточной стороны раствором Рингера, содержащим НЭМ или ИАА в концентрациях 0,5 или 2 мМ. На рис. 4.7 представлены вольт-амперные характеристики фрагмента мембраны в растворе Рингера (1) и после добавления с внутриклеточной стороны 0,5 мМ (2) и 2 мМ (3) НЭМ. В присутствии 0,5 мМ НЭМ проводимость мембраны увеличилась почти в два раза по сравнению с контролем. При увеличении концентрации НЭМ проводимость мембраны еще несколько возросла.
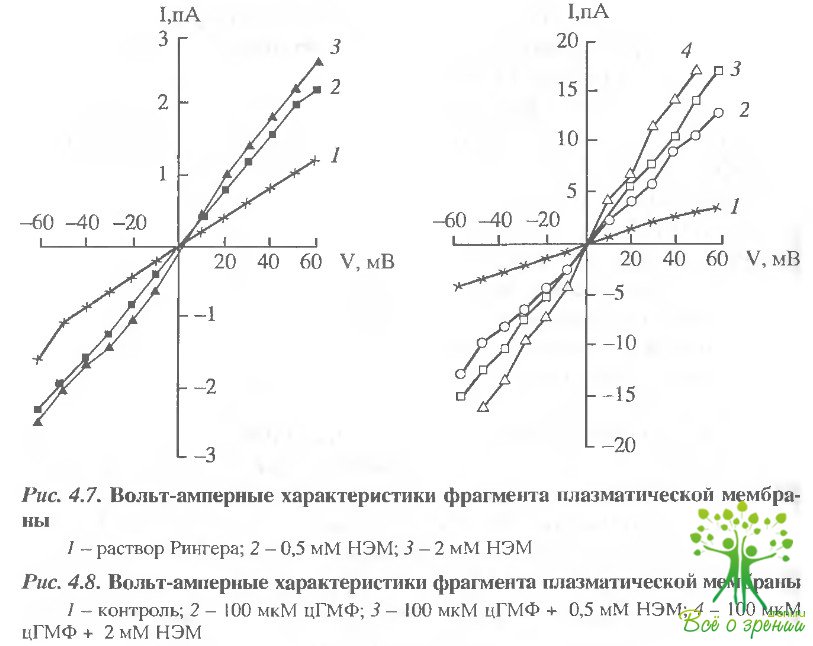
В следующей серии экспериментов (рис. 4.8) мы добавляли в среду инкубации с внутриклеточной стороны 100 мкМ цГМФ, Известно, что при такой концентрации цГМФ более 80% цГМФ-регулируемых каналов в мембране находятся в открытом состоянии. Добавка 100 мкМ цГМФ вызывала увеличение проводимости фрагмента мембраны примерно в 4 раза по сравнению с контролем. Добавка 0,5 и 2 мМ НЭМ на фоне 100 мкМ цГМФ вызывала дальнейшее увеличение проводимости фрагмента мембраны в 1,3 и 1,6 раз соответственно. Абсолютное увеличение проводимости, вызванное добавкой 2 мМ НЭМ, в / присутствии цГМФ было в 5 раз больше, чем без цГМФ, когда все каналы закрыты (130 и 25 пСм соответственно).
Аналогичный результат вызывала аппликация с внутриклеточной стороны ИАА (рис. 4.9). Добавка 2 мМ ИАА повышала проводимость фрагмента мембраны примерно в 2 раза.
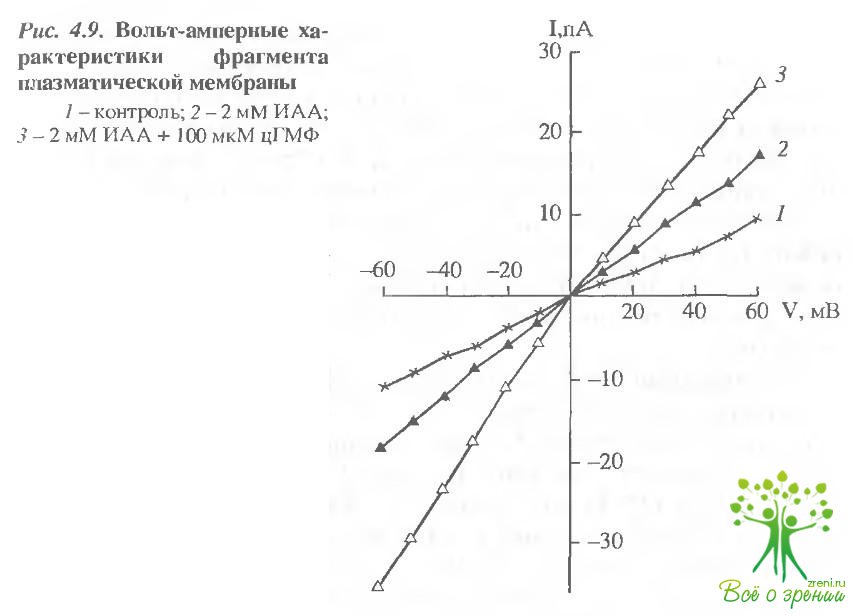
Добавка цГМФ на фоне 2 мМ ИАА также вызывала увеличение проводимости мембраны. Таким образом, SН-связывающие реагенты вызывают увеличение проводимости плазматической мембраны палочки, однако при этом сохраняется цГМФ-регулируемая проводимость мембраны. По абсолютной величине изменение проводимости, вызнанное SН-реагентами в плазматической мембране, значительно превышает изменения проводимости мембраны диска, вызванные модификацией SН-групп родопсина. Следовательно, значительные изменения проводимости плазматической мембраны не могут быть обусловлены модификацией БН групп содержащегося в ней родопсина. По-видимому, канальный белок содержит SН-группы, модификация которых приводит к изменению состояния проводимости капала.
В следующей серии опытов для исследования действия модификации SН-групп с наружной стороны плазматической мембраны SН-реагенты добавляли в раствор, омывающий наружный сегмент изолированной палочки саламандры, внутренний сегмент которой затянут во всасывающую пипетку, и регистрирова ли вызванные этим воздействием изменения тока палочки.?
Поскольку в экспериментах по измерению тока оценивалось действие SН-реагентов на интегральную проводимость фрагмента мембраны, а не на свойства одиночных каналов, при интерпретации результатов следует рассмотреть все возможные мишени для SН-реагентов в плазматической мембране. Ими могли бы быть канальный белок, Nа+-Са2+-обменник и родопсин.
В опытах на дисках мы показали, что модификация 8Н-групп родопсина оказывает незначительное действие на проводимость мембраны и вряд ли может вносить существенный вклад в заметное повышение проводимости фрагмента плазматической мембраны.
По-видимому, наблюдаемый нами эффект не связан также и с модификацией SН-групп Na+-Са2+-обменника. Известно, что длительное воздействие SН-сьязывающего реагента дитиотреитола подавляет активность Na+-Са2+-обменника. Однако известно также, что ингибирование Nа+-Са2+-обмена приводит к полной потере световой чувствительности палочки, а в опытах с ИАА, который действует только с наружной стороны мембраны, снижения чувствительности не наблюдалось. Кроме того, если бы имело место ингибирование Ка+-Са2+-обменника, наблюдалось бы снижение проводимости, поскольку обменник электрогенный, в то время как в наших экспериментах наблюдалось повышение проводимости. Наконец, в опытах на фрагментах мембраны среда внутри пипетки и в кювете была одинаковой, и на мембране не было каких-либо ионных градиентов, в то время как Nа+ -Са2+-обменник функционирует в условиях градиента Са2+ на плазматической мембране. Поэтому скорее всего в экспериментах с мембранными фрагментами Nа+-Са2+-обменник вообще не вносил вклада в интегральную проводимость фрагмента мембраны.
Таким образом, есть основания полагать, что проводимость мембраны меняется в результате модификации SН-групп канального белка. Увеличение проводимости мембраны наблюдается как в присутствии насыщающей концентрации цГМФ ("темповое" состояние мембраны), так и без цГМФ (имитация насыщающего света, когда все каналы закрыты). Следовательно, модификация SН-групп приводит к появлению не зависящей от света компоненты проводимости канала.
Влияние N0 на цГМФ-зиоасимый ток. Недавно было обнаружено, что N0 может прямо влиять на активность нуклеотидрогулируемых каналов в других рецепторных тканях. Так, было найдено, что оксид азота активирует каналы обонятельных нейронов и слуховых волосковых клеток. Обнаруженная нами в наружных сегментах фоторецепторов сравнительно высокая активность NO-синтазы позволяла предположить, что если NO подобным образом прямо активирует цГМФ-зависимые каналы фоторецепторов, тогда оксид азота мог бы являться еще одним звеном в механизме модуляции процесса фототрансдукции. Такое предположение выглядит тем более естественным, что ранее нами была обнаружена в отсутствие пГМФ прямая активация пуклеотцдрегулируемых каналов в результате модификации сульфгидрильных групп, эти эксперименты описаны в предыдущем разделе.
Для того чтобы понять, существует ли прямое влияние оксида азота на работу цГМФ-акгивируемых каналов фоторепепторных клеток, исследовалось изменение интегрального тока в ответ на добавление доноров NO в перфузионный раствор. На рис. 4.10 приведены типичные результаты регистрации интегральных токов методом "пэтч-кламп". Для регистрации использовались наружные сегменты фоторецепторных палочек.

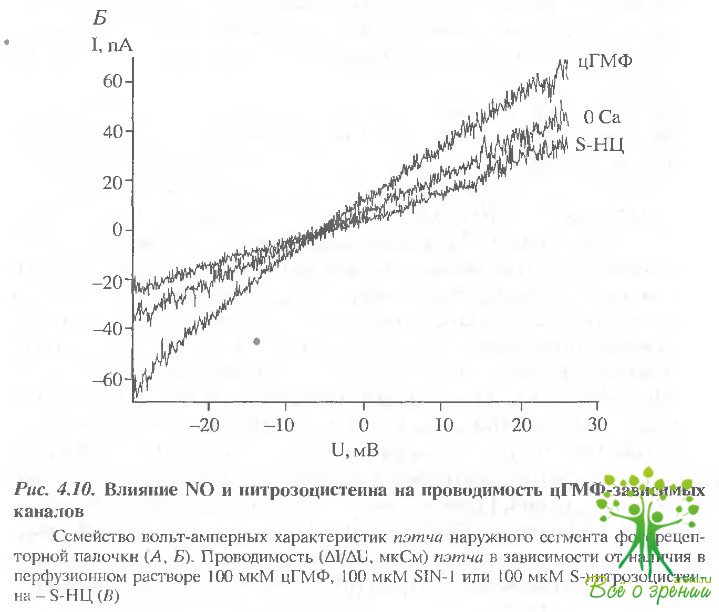
Регистрировалась вольт-амперная характеристика пэтча, содержащего цГМФ-активируемые каналы, в интервале от -30 до +40 мВ.
На рис. 4.10, А, Б представлены вольт-амперные характеристики одного пэтча. На рис. 4.10, В изображена соответствующая диаграмма, отражающая изменение проводимости в ходе одного эксперимента в зависимости от добавления в перфузирующий раствор цГМФ, S1N-1 или S-нитрозоцистеина. Как и должно быть, добавление цГМФ в насыщающей концентрации вызывает обратимое увеличение проводимости почти в два раза. Однако добавление в перфузирующий раствор доноров NO - SIN-1 и S-ни грозоцистеина в отсутствие цГМФ не вызывало ощутимого изменения проводимости, как это наблюдалось в случае слуховых волосковых клеток и обонятельных нейронов.
В состав канального белка зрительных клеток входят несколько цистеиновых остатков, причем многие из них являются ключевыми для механизма работы канала. Например, в ?-субъединице из фоторецепторной палочки цыпленка есть семь эндогенных цистеинов. Один из них - С-481. вызывающий потенциирование ЦН-канала, модифицируется предпочтительнее в открытом состоянии, другой - С-505, вызывающий ингибирование ЦН-ханала, модифицируется предпочтительнее, когда с каналом не связаны лиганды. Существенное повышение на механизм открывания ЦН-каналов оказывает взаимодействие между N- и С-концевыми участками субъединиц, причем взаимодействие, видимо, происходит между двумя цистеиновыми остатками, в ?-субъединице палочкового канала это С-35 и С-481.

Модификация цистеинов ЦН-каналов из фоторецепторных клеток приводит к их активации и в отсутствие цГМФ. Более того, было установлено, что интегральный ток зрительной клетки, возникающий вследствие активации цистеинов ЦН-каналов, был на 20% выше, чем цГМФ-активируемый ток. В 1996 г. аналогичный эффект был обнаружен в обонятельных ЦН-каналах. Было установлено, что активация канала может вызываться не только модификацией SH-групп, например, малеимидом, но и действием оксида азота. Так, при изучении ЦН-каналов обонятельных рецепторов крысы выяснили, что реагенты, высвобождающие NO, могут прямо активировать канал в отсутствие циклических нуклеотидов, вызывая S-нитролизирование свободных SH-групп цистеи- новых остатков. Для нахождения мишени для NO в ? -субъединицах ЦН-канала обонятельного нейрона крысы были последовательно произведены замены четырех разных цистеиновых остатков, расположенных во внутриклеточной части канала (С-460, С-484, С-520, С-552), на серин. В результате было выяснено, что только замена C-460S лишает канал чувствительности к NO. Как уже было сказано выше, С 460 расположен в Ц-связывающей области ЦН-каиала, которая играет существенную роль в механизме открывания канала. Можно полагать, что обнаруженный нами ранее эффект обусловлен действием на гомологичные цистеины в структуре канала из фоторецепторной клетки.
↑ Действие реагентов, модифицирующих SH-группы, на цАМФ-регулируемые ионные каналы из слуховых волосковых клеток
Итак, модификация SH-групп приводит к активации нуклеотидрегулируемых каналов как зрительных, так и обонятельных клеток, а оксид азота может активировать только каналы из обонятельных клеток. В этой связи интересно выяснить, как ведут себя цАМФ-регулируемые ионные каналы из слуховых волосковых клеток.
Типичная запись активности одиночных ионных каналов из волосковых клеток морской свинки представлена на рис. 4.11. В отсутствие цАМФ в симметричном растворе, не содержащем двухвалентных катионов, не наблюдается никакой активности ионных каналов.
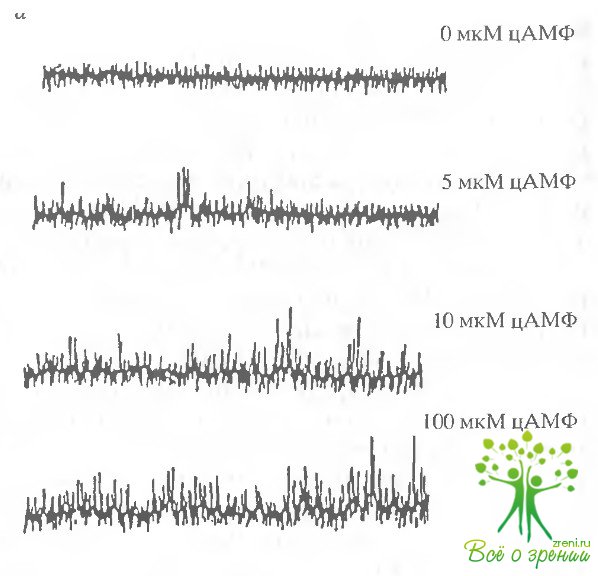
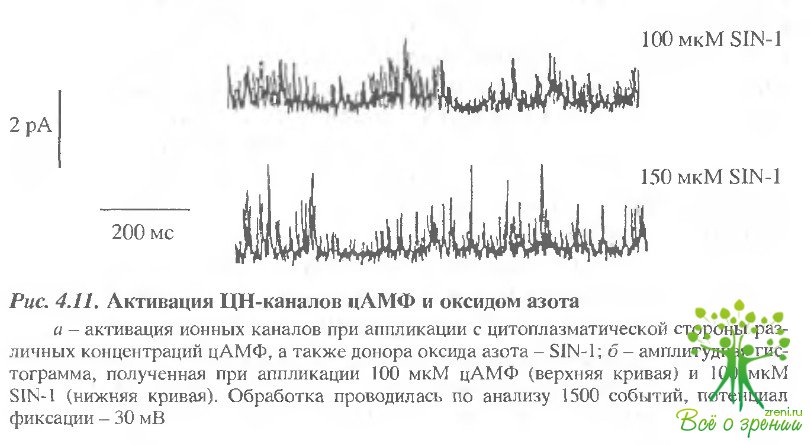
Добавление с внешней стороны пэтча 50 мкМ цАМФ приводило к появлению "выбросов", характерных для активности каналов. Такой эффект в данном исследовании наблюдался в 12 из 43 пэтчей. Эффект был полностью обратим, и отмывание цАМФ приводило к исчезновению характерных "выбросов".
В случае, когда к тому же препарату добавлялись вещества, продуцирующие оксид азота (100 мкМ SIN-1, или 150 мкМ S-нитрозоцистеин), это приводило к активации каналов, аналогичной той, что наблюдалась при добавлении циклических нуклеотидов. В том случае, когда пэтч не содержал ионных каналов, добавление S-нитрозоцистеина и SIN-1 было неэффективным и напротив, в большинстве случаев, когда пэтч содержал ионные каналы, активируемые цАМФ, они также могли активироваться и донорами оксида азота.
Как SIN-I, так и S-нитрозоцистеин могут продуцировать оксид азота только в течение непродолжительного времени Мы измеряли содержание оксида азота в растворах этих соединений через 10 ч после их приготовления, и концентоация оксида азота в них падала в 10 раз, а после суток хранения концентрация оксида азота становилась практически неизмеримой. Когда раствор SIN-T или S-читрозоцистеин после суток хранения добавлялся к пэтчу, содержащему ЦН-каналы, то это не приводило к их активации. Такой контрольный эксперимент свидетельствует о том, что каналы активирует именно оксид азота, а не производные его доноров, образующиеся при их разложении.
В отличие от действия цАМФ, эффект оксида азота был слабо обратим. Если отмывание цАМФ приводило практически мгновенно к исчезновению выбросов, характерных для каналов, то отмывание SIN-1 и S-нитрозоцистеина приводило к их медленному исчезновению; полностью эффект оксида азота исчезал только через 20 мин после полного отмывания реагентов.
В том случае, когда SIN-1 и S-нитрозоцистеин добавлялись на фоне цАМФ, это не приводило к каким-либо заметным изменениям в активности каналов, однако в этом случае после отмывания реагентов эффект был плохо обратим.
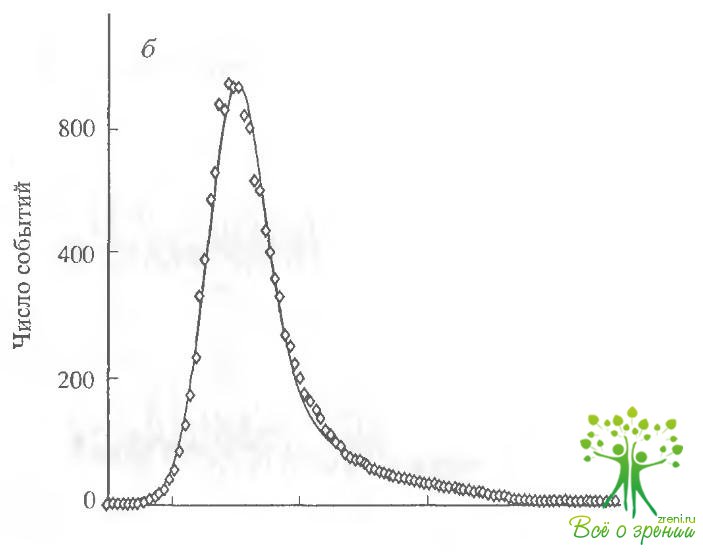
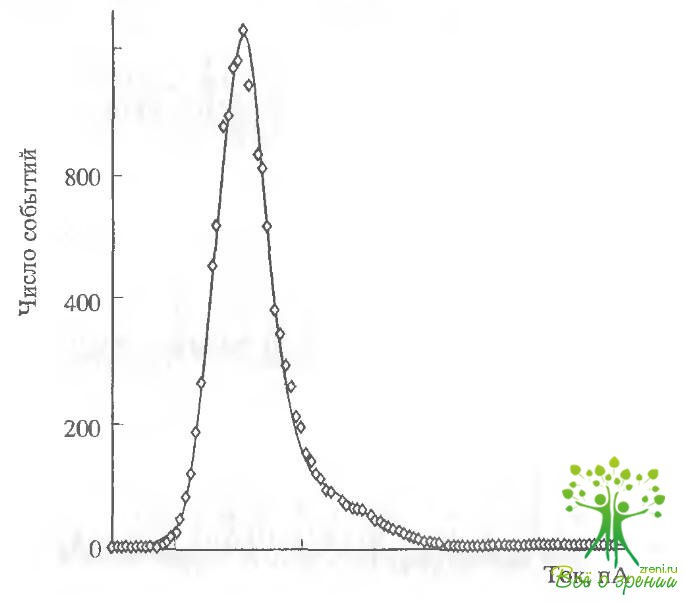
На рис. 4.11, б представлены амплитудные гистограммы, полученные при обработке результатов регистрации тока через мембранный фрагмент при действии на него цАМФ и SIN-1. Гистограммы хорошо аппроксимируются кривыми Гаусса, что позволяет определить ток одиночного канала. Величины токов оказались достаточно близкими для каналов, активированных цГМФ и SIN-1. Так, в присутствии 50 мкМ цАМФ единичная проводимость составляла 1,5 пА, а в присутствии 100 мкМ SIN-1 - 1,7 пА для гистограмм, показанных на рисунке. Величины, усредненные по шести аналогичным опытам, составили 1,62 ± 0,21 и 1,74 ± 0,15 пА для цАМФ и SIN-1 соответственно (хотя величины одиночной проводимости каналов, активированных разными способами и близки, величина проводимости одиночного канала, активированного SIN-1, всегда была несколько выше, чем при активации цАМФ).
Для более точного описания характеристик каналов мы определяли среднее время нахождения канала в открытом и закрытом состояниях для обоих активирующих агонистов. В обоих случаях кривые хорошо аппроксимировались одной экспонентой и давали следующие величины. Постоянная времени открытого состояния для цАМФ составляла примерно 0,45 с, а для SIN-1 -0,55 с.
Для постоянной времени закрытого состояния были получены величины 0,83 и 1,03 с для цАМФ и SIN-1 соответственно (результаты получены при обработке непрерывной записи активности каналов в течение 40 мин).
Реагенты, модифицирующие SH-группы, активируют ЦН- какалы аналогично оксиду азота.Ранее нами было установлено, что реагенты, модифицирующие сульфгидрильные группы (иодацетамид, парахлормеркурийбензойная кислота), будучи добавленными к пэтчу, содержащему ЦН-регулируемые каналы, могли активировать ток и в отсутствие цГМФ. Аналогичный эффект был обнаружен нами и при исследовании цАМФ-регулируемых каналов в волосковых клетках.
На рис. 4.12 показаны результаты такого исследования Добавление 1 мМ иодацетамида приводило к активации каналов аналогично цАМФ или оксиду азота.
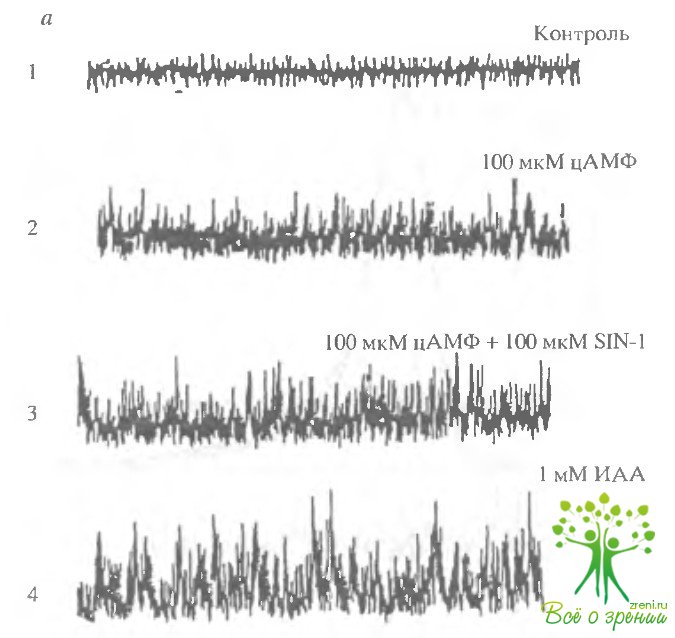
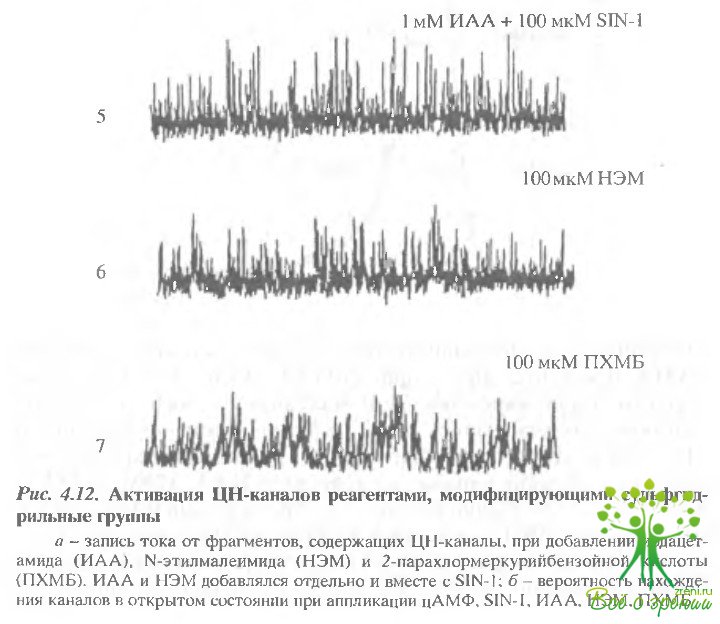
Аналогичное действие оказывал также N-этилмалеймид и парахлормекурийбензойная кислота (ПХМБ).
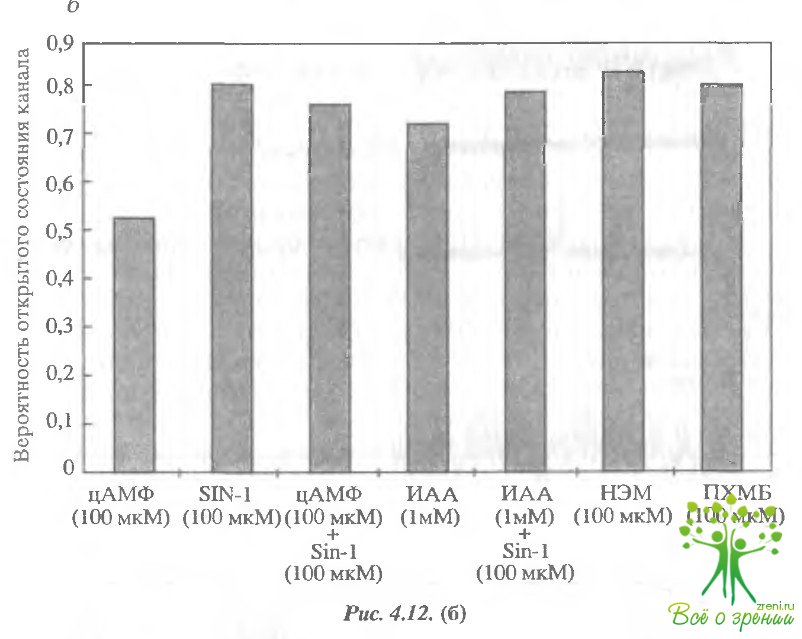
Действие всех реагентов, модифицирующих SH-группы более напоминало действие доноров оксида азота, чем цАМФ.
Действительно:
- эффект ИАА, НЭМ и ПХМБ был необратим, и состояние каналов не восстанавливалось после их отмывания (как отмечалось ранее, действие S-нитрозоцистеина и SIN-1 также было слабообратчмым);
- постоянная времени открытого состояния канала под действием ИАА, НЭМ и ПХМБ возрастала существенно больше, чем при действии цАМФ;
- добавление как SIN-1, так и ИАА на фоне цАМФ приводило к дополнительному возрастанию постоянной времени открытого состояния примерно на 30%. Эти факты позволяют предполагать, что механизм действия оксида азота и реагентов, модифицирующих сулофгидрильные группы, одинаков, а именно: добавление N0 приводит к S-нитрозилированию SH-групп цистеиновых остатков.
NO и цАМФ действуют на разные связывающие центры канала.Для того чтобы выяснить, действуют ли цАМФ и NO на один и тот же сайт в молекуле канала, мы провели эксперименты с аналогом цАМФ - Rp-цАМФ. Это соединение является ингибитором протеинкиназы A и, как показано для ЦН-активируемых каналов из обонятельных нейронов, связывается с каналом, не активируя его. Таким образом, если бы оксид азота действовал на тот же центр, что и цАМФ, после действия Rp-цАМФ его эффект не проявлялся бы.
Результаты такого эксперимента представлены на рис. 4.13. Эксперимент проводился обычно по следующему протоколу. Вначале регистрировался ток через мембранный фрагмент при добавлении 100 мкМ цАМФ, чтобы убедиться, что фрагмент содержит хотя бы один ЦН-канал. Затем цАМФ отмывался и добавлялось 500 мкМ Rp-цАМФ. Это не приводило к активации ЦН-каналов.
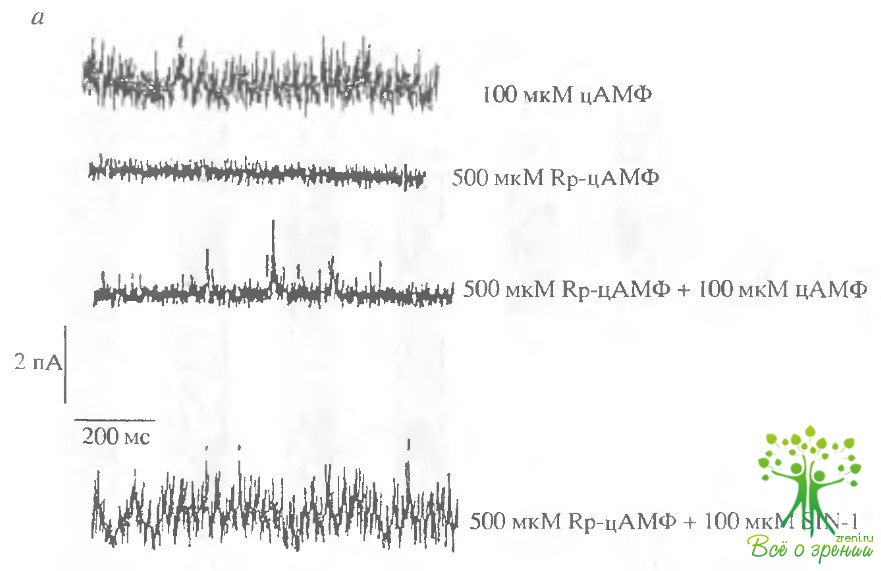
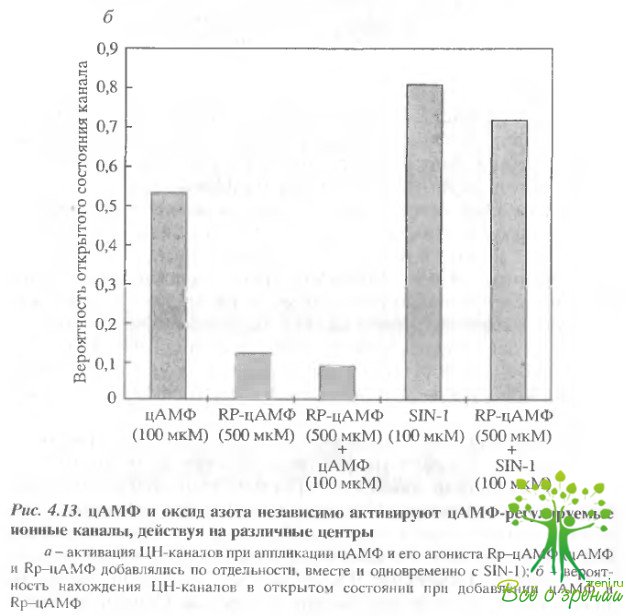
Добавление цАМФ на фоне Rp-цАМФ также не приводило к регистрации активных каналов. Однако добавление на фоне Rp-цАМФ 100 мкМ SIN-1 приводило к регистрации каналов.
В описанных экспериментах показано, что ЦП-каналы из волосковых клеток органа Корти могут быть активированы не только цАМФ, но и веществами, продуцирующими оксид азота. Такая активация не связана, как это наблюдается в большинстве случаев, с активацией циклазы и увеличением концентрации циклических нуклеотидов. Эффект наблюдается на мембранном фрагменте клетки и в отсутствие АТФ.
Оксид азота активирует тот же самый канал, что и цАМФ. Во-первых, действие доноров NO наблюдалось только в тех пэтчах, в которых присутствовали цАГЛФ-регулируемые ионные каналы. Во-вторых, основные кинетические характеристики каналов были аналогичными. Существенное отличие наблюдалось только в вероятности открытого состояния канала. Эта величина при активации оксидом азота была обычно на 30% выше, чем при активации цАМФ. Это подтверждено и при исследовании интегрального тока. Вольт-амперные характеристики, полученные на фрагментах, содержащих несколько каналов, демонстрируют большую величину тока при насыщающих концентрациях оксида азота, чем при насыщающих концентрациях цАМФ.
Эффект оксида азота обусловлен прямым его действием на ионный канал, причем наиболее вероятным механизмом такого действия является модификация оксидом азота сульфгидрильных групп в ионном канале. Во-первых, нами показано, что реагенты, модифицирующие SH-группы, сами по себе в отсутствие оксида азота или цАМФ способны активировать канал. Во-вторых, действие всех SH-реагентов (ИАА, НЭМ, ПХМБ) было аналогично действию оксида азота и несколько отличалось от действия цАМФ. SH-реагенты, аналогично оксиду азота, в насыщающих концентрациях вызывали больший интегральный ток через мембранный фрагмент, чем насыщающая концентрация цАМФ.
То, что оксид азота может действовать прямо через модификацию сульфгидрильных групп, не является удивительным. Взаимодействие предельно окисленного оксида азота (NOT) является весьма вероятной химической реакцией (см. обзор Lipton, 1993). Предполагается также, что именно нитрозотиолы служат тем стабильным соединением, которое и позволяет оксиду азота быть таким важным физиологическим соединением. Сам по себе оксид азот является крайне нестабильным соединением и маловероятно, что он может быть эффективным медиатором сам по себе. То, что модификация сульфгидрильных групп может активировать канал в отсутствие циклических нуклеотидов, было ранее продемонстрировано нами на примере каналов фоторецепторной клетки, а также показано для ЦН-каналов из обонятельных нейронов.
Интересным фактом в нашем случае является то, что NО и цАМФ действуют на различные связывающие центры в молекуле ионного канала Действительно, оксид азота вызывал активацию канала и в том случае, когда цАМФ-сайт был блокирован Rp-цАМФ. Все использованные нами SH-реагенты не проникают через мембрану и взаимодействовали только с цитогшазматической стороной пэтча Таким образом, можно предположить, что оба связывающих центра ионных каналов, которые могут активировать канал, локализованы с цитоплазматической стороны.
Статья из книги: Молекулярные механизмы зрительной рецепции | Каламкаров Г.Р., Островский М.А..

Комментариев 0