Регуляция рН в зрительной клетке и ее роль в механизмах фототрансдукции (ЧАСТЬ І)


Содержание:
Описание
↑ Процессы, вызывающие закисление в зрительной клетке
В фоторецепторах постоянно происходят процессы, которые могли бы вызывать быстрое внутриклеточное закисление. Во внутреннем сегменте клетки это интенсивный метаболизм, приводящий к образованию лахтата и CO2. В наружном сегменте - процессы синтеза и гидролиза цГМФ. Ниже приводятся оценки интенсивности производства Н+ в ходе этих процессов.
Наружный сегмент. Как при гидролизе, так и при синтезе цГМФ выделяется один протон. Темновая скорость гидролиза цГМФ составляет, по оценкам, 2,5-3 мМ в секунду, что предполагает выделение 5-6 мМ Н+/с в темноте. При интенсивном освещении скорость гидролиза цГМФ возрастает примерно в 10 раз, что приводит к выделению -50 мМН /с.
Внутренний сегмент. Большая часть АТФ образуется во внутреннем сегменте в результате анаэробных процессов. Это подтверждается косвенно и тем, что интактная сетчатка достаточно долго активна в бескислородной среде. Здесь мы приведем оценки для двух случаев:
- АТФ продуцируется исключительно за счет окислительного фосфорилирования,
- АТФ производится исключительно вследствие гликолиза.
1. Темновой ток в палочке саламандры составляет 80 рА, что соответствует 80 х 10-17молекул/с. Предполагая, что этот ток поддерживается исключительно Na+- K+- АТФазой и учитывая, что 1 молекула АТФ обеспечивает перенос 3 ионов Nа+, получим 28 х 10-17 молекул АТФ в секунду. При окислительном фосaорилировании 36 АТФ и 6 С02генерируется на одyу молекулу глюкозы и таким образом должно производиться (28/6) х 10-18молекул С02 в секунду. Учитывая, что объем ВС составляет 1,6 пл, при полностью аэробных условиях скорость производства СО, составит 30 мМ/с.
Конечно, следует учесть, что при низких значениях рН С02 будет легко диффундировать, не гидрируясь, из клетки, однако рК карбоновой кислоты составляет 6,1. Следует также учитывать, что скорость диффузии С02 зависит от типа клетки.
2. При анаэробных условиях образуются две молекулы АТФ и две молекулы лактата на молекулу глюкозы. Таким образом, лактат продуцируется с скоростью 28 х 10-17молекул/с, или 170 мМ/г,
В стационарном состоянии ресинтез ГТФ из цГМФ должен осуществляться при поглощении Н+, и это должно было бы создавать баланс. Однако следует учесть, что ферменты, осуществляющие эту реакцию, локализованы не в наружном, а во внутреннем сегменте клетки и, таким образом, эта реутилизация не может предотвратить ацидоз в НСП.
Существование таких мощных источников закисления требует наличия в клетке специальных механизмов поддержания оптимального внутриклеточного рН. Обычно внутриклеточный рН поддерживается в состоянии выше термодинамического равновесия, и такой механизм должен работать против электрохимического градиента. Предотвратить внутриклеточный ацидоз можно, экстапгируя из клетки Н+ или закачивая в клетку НС03. В последнем случае НС03 взаимодействует с протоном, образуя С02, который может диффундировать из клетки Соответственно известны два типа обменников: Nа+/Н+-обменник и НСO3/Cl- обменник.
Nа+/Н+-обмен как способ поддержания рН обнаружен в плазматических мембранах многих клеток. Одной из характерных черт этого процесса является то, что он ингибируется амилоридом и другими производными пиразина. Nа+/Н+-обмен может быть также заингибирован заменой в среде Na+ на К+. Эти два теста и используются обычно для его обнаружения в клетках.
Существуют гри типа НСO3/С1 -обмена в клетках: Na-независимый НСO3/Сl-обмен, сопряженный с Na+ НСО3 /Сl-обмен и электрогенный Na+/HCO3/Cl -транспорт.
В данном разделе изложены результаты наших исследований механизмов регуляции рН в фоторецепторных клетках и их роли в модуляции процессов лототрансдукции.
↑ Регистрация внутриклеточного рН в слое фоторецепторов
Для доказательства присутствия в зрительной клетке указанных выше механизмов мы прямо измеряли внутриклеточный рН с помощью флуоресцентных красителей, пытаясь различными способами ингибировать или активировать возможные механизмы поддержания рН - Na+/H+- и HCO3/Сl-обменники.
Внутриклеточный рН измерялся в фоторецепторных клетках полностью обесцвеченной сетчатки лягушки с помощью внутриклеточного красителя 2,7,-бис(2-карбоксиэтил)-5,6-карбокси-флуоресцеин (БКФ). Флуоресценция в пике эмиссии (525 нм) при возбуждении светом 490 нм линейно зависит от величины рН. Однако эти изменения могут включать также и не зависимые от рН изменения, в частности, утечку красителя из клетки. Эти изменения могут быть учтены при регистрации эмиссии при 440 нм, которая не зависит от рН. Таким образом, измерение отношения I440/I525обусловлено только изменениями внутриклеточного рН.
Флуоресценция измерялась прямо в слое фоторецепторов в интактной сетчатке, что давало определенные преимущества. Во-первых, сигнал флуоресценции, усредненный от MHOI их клеток, был значительно интенсивнее, что позволяло получать значительно лучшее отношение сигнал-шум; во-вторых, фоторецепторы в изолированной сетчатке значительно нативнее. Такое измерение удавалось благодаря морфологическим особенностям сетчатки. Слой фоторецепторов занимает всю ее поверхность на глубину 60 мкм, что дает возможность регистрировать флуоресценцию от поверхности при том, что вклад других слоев значительно ниже. Кроме того, и это наиболее важно, добавляя краситель к сетчатке с рецепторной стороны и отмывая его через несколько минут, удалось найти условия, в которых флуоресцентный краситель проникал только в фоторецепторные клетки. Распределение красителя проверялось в специальных экспериментах. Его концентрация определялась в проксимальных срезах толщиной 20 мкм (т.е. в условиях, когда удавалось разделять наружный и внутренний сегменты) Результаты гель-электрофореза 6 таких срезов показаны на рис. 6.1. Видно, что в срезах наружных сегментов присутствуют только родопсин, трансдуцин и арестин - характерные белки наружного сегмента. Отсутствие в ощутимых количествах других белков свидетельствует о том, что в срезах наружного сегмента отсутствуют примеси внутреннего сегмента.
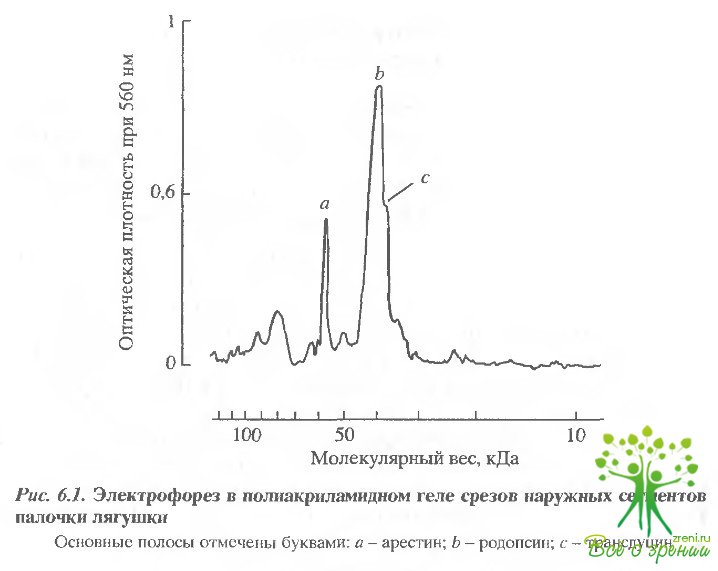
В срезах внутреннего сегмента, напротив, практически нет родопсина и, кроме других белков, присутствует арестин, значительное количество которого, как известно, локализовано именно во внутреннем сегменте. Кроме того, срезы наружных сегментов не содержали сукцинатдегидрогеназы - маркерного белка митохондрий, которых много во внутреннем сегменте, но которые отсутствуют в наружном.
Раствор, содержащий БКФ, каплями наносился на сетчатку с рецепторной стороны и после трехминутной инкубации сетчатка промывалась чистым раствором Рингера. При таких условиях обработки флуоресценция была обнаружена только в наружных сегментах, и поэтому можно предполагать, что изменения флуоресценции отражают по крайней мере изменения рН в фоторецепторном слое сетчатки Кроме того, сетчатка помещалась в измерительный отсек так, что к ФЭУ была обращена ее фоторецепторная сторона. Поскольку интенсивность регистрируемой флуоресценции сильно падает с толщиной препарата, то это создавало дополнительные условия для регистрации флуоресценции именно от фоторецепторного слоя. Стационарное значение внутриклеточного рН в начале эксперимента в растворе Рингера, не содержащем бикарбоната, составило 6,68 ± 0,05 (рН 7.5, среднее по 11 экспериментам). Такое значение несколько ниже того, которое обычно находят в нейронах, ц причины этого будут изложены в обсуждении. На величину внутриклеточного рН существенно влияло присутствие в среде бикарбоната. Так, в 7 случаях сетчатка с самого начала инкубировалась в растворе Рингера, содержащем бикарбонат. Значение рН увеличивалось в этом случае до 6,90 ±0,15.
↑ Nа+/Н+ и НСО3/С1 -обмен в зрительных клетках
Существуют два, хотя и не очень специфичных, но достаточно достоверных способа выявления Nа+/Н+-обмена в клетках.
- Замена экстраклеточного на К+[/sup]или холин должна приводить к внутриклеточному закислению, так как калий не может заменить натрий в процессе обмена.
- Амилорид - блокатор практически всех трансмембранных процессов, включающих участие натрия, также должен вызывать внутриклеточное закисление.
Na+на К+ заменялся в растворе Рингера, не содержащем бикарбоната, чтобы свести к минимуму влияние регуляторов рН, использующих градиент бикарбоната в среде. Типичный эксперимент такого рода представлен на рис. 6.2. Замена в растворе 95 мМ Na+, 3 мМ К+ на 98 мМ К+ приводит к медленному монотонному закислению внутриклеточной среды.

В восьми подобных экспериментах средняя скорость закисления составила 0,024 единицы рН в минуту. После того как внутриклеточный рН уменьшался на 0,08 единицы, К+ заменяли на Na+, и это приводило к медленному восстановлению внутриклеточного рН. В том случае, когда внутриклеточное закисление не превышало 0,3 единицы рН, эффект обычно был полностью обратим. В тех же случаях, когда рН изменялся на 0,6-0,8 единиц, рН внутри клетки уменьшался необратимо.
Добавление 1 мМ амилорида в раствор, не содержащий би-карбоната, всегда приводило к внутриклеточному закислению (рис. 6.3). Однако замена раствора, содержащего амилорид, на обычный раствор Рингера не приводила к восстановлению исходного рН, а лишь приостанавливала процесс внутриклеточного закисления.

Это связано, по-видимому, с тем, что даже при длительной перфузиине удается полностью отмыть амилорид, и значительная часть обменника остается блокированной. В некоторых случаях, когда отмывание амилорида происходило при относительно небольшом изменении внутриклеточного рН (не больше чем на 0,2 ед.), нам удавалось достигнуть практически полного восстановления исходного рН.
↑ Транспорт бикарбоната в фоторецьпторах
Добавление бикарбоната увеличивает рН. В экспериментах, представленных на рис. 6.4Л, сетчатка инкубировалась сначала в растворе Рангера, не содержащем бикарбоната (12 мМ фосфатный буфер). После стабилизации рН 6 мМ фосфата в растворе заменялось на 6 мМ бикарбоната. В шести подобных экспериментах эта процедура приводила к внутриклеточному защелачиванию. Эффект был обратимым, при обратной замене на фосфат первоначальное значение рН восстанавливалось. В 10 подобных экспериментах рН увеличивался на 0,26 ± 0,04 единицы рН.
Следует отметить, что увеличение концентрации НССГ при постоянном значении внеклеточного рН должно обязательно сопровождаться увеличением концентрации С02как снаружи, так и внутри клетки, поскольку С02 свободно диффундирует через мембраны. Гидрирование внутри клетки должно вызывать освобождение протона и таким образом уменьшать величину рН. Тем не менее мы наблюдаем подщелачивание, и это является сильным аргументом в пользу того, что в клетке присутствует механизм транспорта бикарбоната. Может показаться странным (см. рис. 6.4,А), что даже в самой начальной фазе не наблюдается закисления. Это обстоятельство, однако, объясняется медленной скоростью гидрирования С02в отсутствие карбоангидразы.

Известно, что НСО3/Сl-обмен ингибируется производными стильбена, такими как ДИДС и СИТС. В наших экспериментах добавление 0,2 мМ ДИДС к раствору, содержащему бикарбонат, приводило к стабильному монотонному закислению (см. рис. 6.4,б). Однако в этом случае, как и с амилоридом, эффект не был полностью обратим. Отмывание ДИДС приводило лишь к предотвращению индуцированного им закисления, но не приводило к восстановлению исходного рН.
Уменьшение концентрации С1~ приводит к росту внутриклеточного рН. В следующей серии экспериментов мы изучали, участвует ли НСО3/Сl-обмен в транспорте бикарбоната. Если это так, то снижение внутриклеточной концентрации С1- должно приводить к ускорению транспорта HCO3 и таким образом к внутриклеточному защелачиванию Замена в среде 95 мМ NaCl на глюконат натрия действительно приводила к существенному снижению внутриклеточного рН (рис. 6.5,А). Эффект был полностью обратимым при обратной замене в растворе глюконата натрия на хлорид натрия.
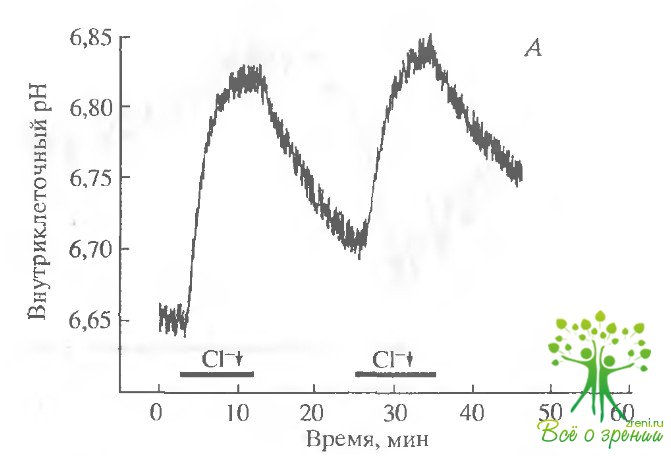
На одной и той же сетчатке эта процедура может быть повторена несколько раз. Более того, внутриклеточное закисление аддитивно. Так, в том случае, когда сначала только половина хлорида замещалась глюконатом, а затем хлорид замещался полностью, повторное закисление было практически таким же, как и первое.
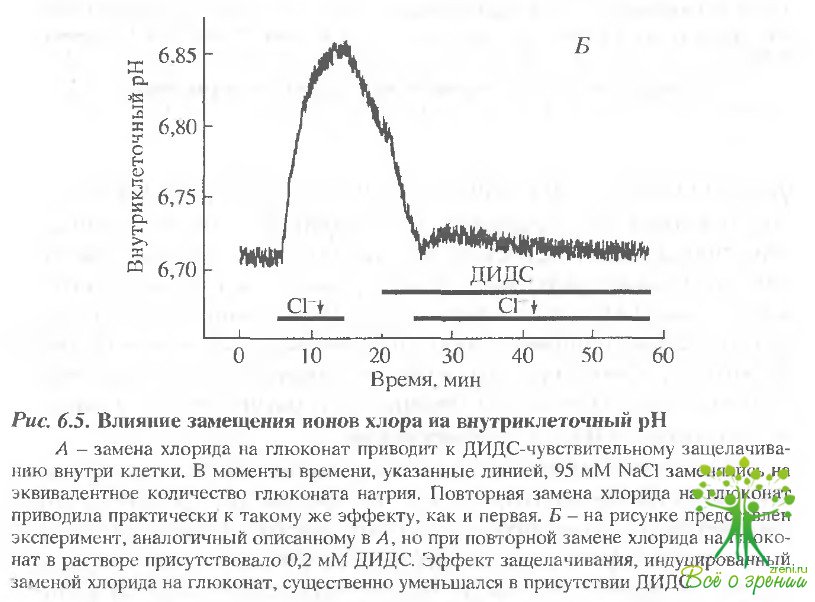
Защелачивание, индуцированное заменой хлора, чувствительно к ДИДС. Если описанный ранее эффект удаления хлора обусловлен НСО3/Сl -обменом, то он должен быть чувстителен к ДИДС. Результаты влияния ДИДС представлены на рис. 6.5,Б. Эксперимент проводился следующим образом. Сначала, как описано выше, 95 мМ Сl~ замещалось глюконатом, что приводило к внутриклеточному защелачиванию. Затем, в момент достижения максимального эффекта, глюконат вновь замещался на хлорид. Последующее добавление ДИДС приводило к сильному ускорению закисления, индуцированного увеличением хлорида в среде. Затем, когда величина рН достигала своего первоначального значения, хлорид натрия вновь замещался на глюконат. Эффект защелачивания в этом случае был значительно меньше.
В фоторецепторкой клетке существует зависимый НСО3/Сl -обменник.Чтобы определить тип обменника, работающего в фоторецепторной мембране, описанные выше эксперименты проводились в безнатриевой среде. Сначала NaСl в среде инкубации замещался на КСl, а затем КСl замещался на глюконат. Если бы эти манипуляции приводили к внутриклеточному
защелачиванию, это означало бы, что НСО3 /Сl-обмен не зависит от натрия. Эксперименты, поставленные в соответствии с таким протоколом, представлены на рис. 6.6. Чтобы избежать сильного закислення, вызванного удалением из среды натрия, хлорид замещался на глюконат уже через 3,5 мин после удаления натрия. Защелачивание, индуцир данное такой заменой, было достаточно сильным.
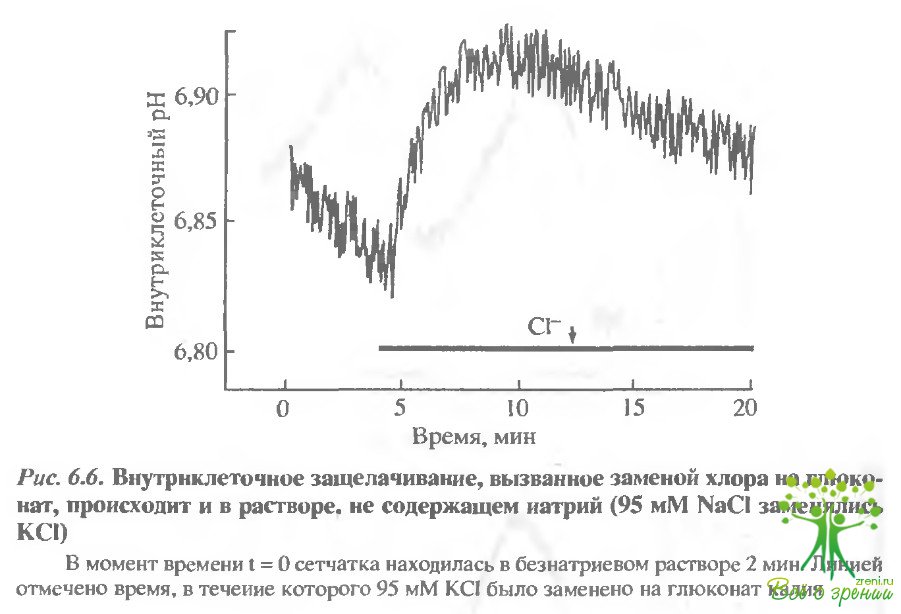
Это позволяет предполагать, что значительная часть транспорта бикарбоната осуществляется натрий-независимым НСО3/Сl- обменником.
Выше отмечалось, что как в наружном, так и во внутреннем сегмента: фотореиепторной клетки происходят процессы, которые могли бы приводить к сильному ацидозу. Очевидно, что рН в фоторецепторе должен поддерживаться в достаточно узком диапазоне значений, так как фоточуствительность клетки зависит от рН. Механизм поддержания рН, использующий натриевый градиент, который на плазматической мембране достаточно высок, представляется оптимальным путем регуляции рН. Менее очевидна роль и натрий-независимого НСО3/Сl -обмена. Если хлорид находится в состоянии термодинамического равновесия, то такой обменник будет стремиться сдвинуть рН к состоянию равновесия. Поскольку натрий-зависимые обменные механизмы поддерживают рН при более низких значениях, то эти два класса обменников будут работать в прот иеоположные стороны. Такой механизм мог бы более эффективно регулировать гомеостаз рН, так как будет приводить клетку к стационарному значению рН быстрее как в случае внутриклеточного закисления, так и в случае внутриклеточного защелачивания.
Однако возможны и другие причины. Хлорид может и не находиться в зрительных клетках в состоянии термодинамического равновесия. Есть данные, что ЕCl в палочках положителен даже при темновых значениях потенциала. In vivo внутриклеточный рН может оказаться достаточно высоким, и Nа+/Н+-обмен не сможет обеспечить его регуляцию, так как имеет низкий рК. В этом случае рН может снижаться за счет экстракции из клетки НС03.
↑ Изменения фоточувствительного тока в палочке при изменении внутриклеточного рН
В предыдущем разделе было показано, что в фоторецепторной клетке содержатся несколько типов ионных обменников, которые в совокупности могут поддерживать внутриклеточный рН в достаточно узком диапазоне значений, обеспечивая оптимальную работу зрительной клетки. Однако сам по себе факт, что в клетке локализованы те или иные обменники, мало говорит о роли этих структур в физиологии фоторецептора. Кроме того, как указывалось выше, фот акцепторная клетка состоит из двух пространственно и функционально разделенных частей. Как источники внутриклеточного закисления, так и процессы, протекающие в этих частях клетки, существенно различаются.
В настоящем разделе мы опишем возможную физиологическую роль описанных выше механизмов регуляции рН отдельно в наружном и внутреннем сегментах фоторецепторной клерки. Используя метод всасывающего электрода, ток можно одинаково эффективно регистрировать, засасывая в пипетку внешний или внутренний сегмент. При этом воздействию перфузата остается доступной только свободная часть клетки, что и позволяет исследовать отдельно внутренний и наружный сегменты.
Результаты, которые приведены в данной главе, свидетельствуют о том, что рН по-разному воздействует на наружный и внутренний сегмент фоторецепторов. Ввиду этою эффекты, наблюдаемые при перфузии внутреннего (ВС) и наружного (НС) сегментов, описаны отдельно. Существенно то, что как темновой ток, так и фототок уменьшаются, когда ионы Н+входят в наружный сегмент фоторецептора и, напротив, увеличиваются, когда Н+удаляется.
Таким образом, когда в экспериментах рН различными способами изменялся, можно было полагать, что, если фототок не изменяется, то и внутриклеточный рН во внешнем сегменте не изменился. Такой подход позволяет использовать фоторецептор в качестве своеобразного рН-электрода и таким образом изучить локализацию обменников в фоторецепторной клетке.
Внутренний сегмент. Эффект амилорида. Чтобы исследовать роль Na+/Н+-обмена, мы использовали амилорид (2 мМ). Как показали измерения внутриклеточного рН, амилорид должен вызывать внутриклеточное закисление и соответственно уменьшение фотоответа. Наружный сегмент фоторецептора засасывался в пипетку, и таким образом был защищен от воздействия амилорида. Действие амилорида всегда приводило к подавлению фототока. Во всех случаях, когда амилорид воздействовал на клетки с большим начальным фототоком (порядка 25 рА), в течение 10 мин его добавление приводило к полной и необратимой потере фоточувствительности. Такой же результат был получен и в растворе Рингера, содержащем бикарбонат (6 мМ) (рис. 6.7).
В тех же клетках, где первоначальная величина фотоответа была мала, его уменьшение происходило за существенно больший промежуток времени (обычно более 1 ч).
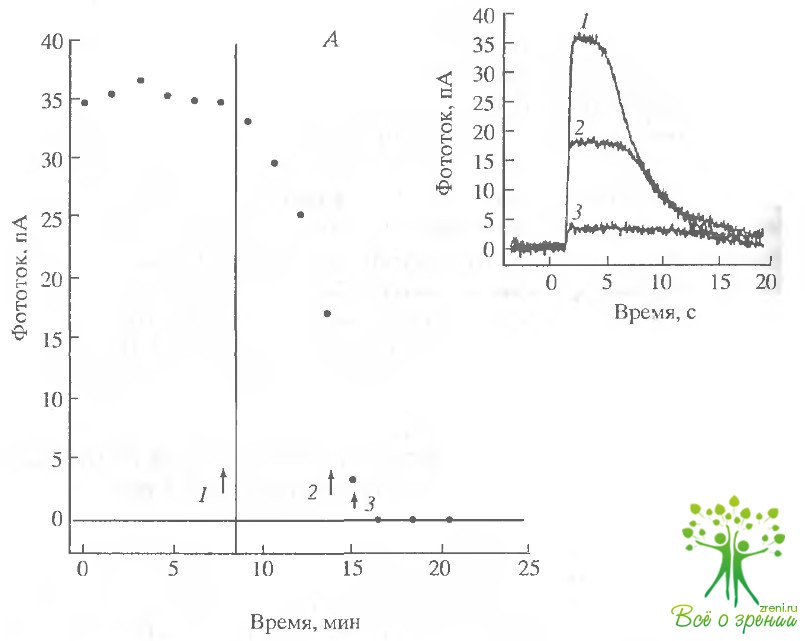
Это может объясняться тем, что в клетках с небольшим значением фототока метаболизм происходит менее интенсивно, и это приводит к более медленному внутриклеточному закислению при ингибировании обменника. Аналогичная закономерность наблюдалась и при действии ДИДС (см. ниже).
Амилорид - не очень специфичное соединение, и наблюдаемый эффект мог бы быть обусловлен действием как на Nа+/Н+-обменник, так и на Na+/Са2+-обменник. Чтобы дифференцировать эффекты, мы провели аналогичные эксперименты в бескальциевой среде, однако не обнаружили существенных различий.
Сохранение темпового тока после потери фоточувствителъности. Наиболее интересным результатом при исследовании внутреннего сегмента было го, что даже после полного исчезновения фотоответов темновой ток частично или полностью сохранялся. Это может служить ключом к пониманию механизма действия рН, так как при закислении наружного сегмента происходит подавление и темнового тока. ЭТОТ вывод был сделан из следующих экспериментов. Когда фототок полностью пропадал, фоторецептор перемещали вдоль пипетки, засасывая его или, напротив, выталкивая. Если темновой ток отсутствует, то такое перемещение не должно сказываться на величине измеряемого тока. Однако в эксперименте величина измеренного таким образом темнового тока практически не изменялась после воздействия амилорида (рис. 6.8).
Подобные манипуляции обычно приводят к изменению давления внутри пипетки, которое могло бы влиять на результаты. Однако это не может объяснить полученный эффект.

Во-первых, давление в пипетке изменяется кратковременно и через 5 с становится равным атмосферному. Во-вторых, если клетка с самого начала не обладала световой чувствительностью. описанные манипуляции не приводили к каким-либо изменениям. Сохранение фототока имело место только в тех случаях, когда амилорид приводил к исчезновению фоточувствительности достаточно быстро.
На рис. 6.8,Б приведены результаты другого типа экспериментов, когда темновой ток и насыщающий фотооверегистрировались одновременно. Это возможно, если базовая линия достаточно стабильна (см. рис. 6.8,Б). В эксперименте, результаты которого представлены на рисунке, амилорид практически мгновенно приводил к падению фототока, однако через 3 мин изменяются как величина фототока, так и нулевая линия, что указывает на го, что значительная часть тока не чувствительна к свету.
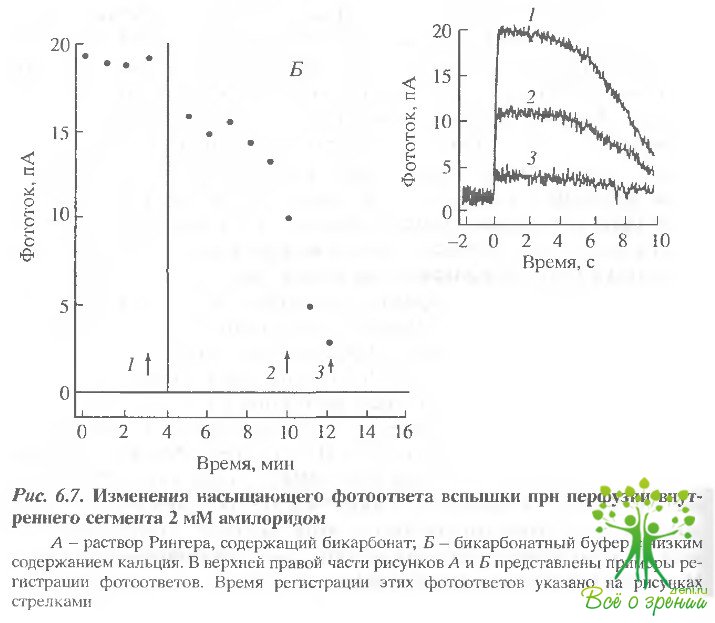
Эффект ДИДС. Для исследования роли НСО3 /Cl -обмена в раствор Рингера, содержащий бикарбонат, добавляли ДИДС. Полученные результаты аналогичны описанным ранее с амилоридом, однако ДИДС действовал обычно медленнее. В клетках с большим первоначальным фототоком значительный эффект при добавлении ДИДС наблюдался только через 10-30 мин после начала перфузии (рис. 6.9). Обратимость эффекта зависела от того, насколько долго ДИДС воздействовал на клетку. В том случае, когда ДИДС начинали отмывать до того как фототок существенно падал, удавалось достигнуть практически полного его восстановления.

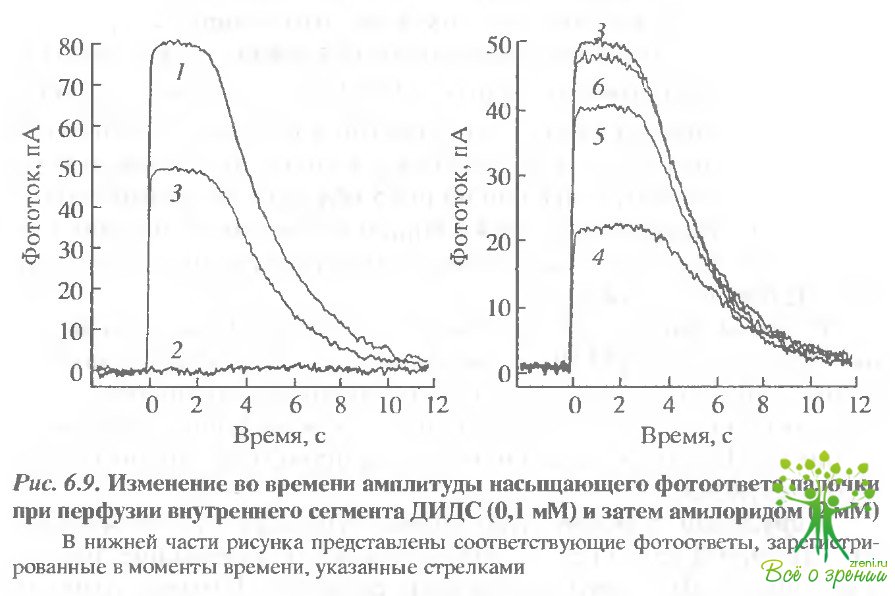
В тех же случаях, когда под действием ингибитора фототок уменьшался почти до нуля, эффект становился необратимым. Как и в случае амилорида, мы обнаружили, что изменения внутриклеточного рН, обусловленные ДИДС, вызывают изменения фототока, однако темновой ток клетки сохраняется. В экспериментах, представленных на рис. 6.9, амилорид добавлялся фазу после отмывания ДИДС. В этом случае его эффект отличался от описанного ранее. Величина фототока сначала уменьшалась примерно на 50%, а затем в течение 10 мин восстанавливалась до уровня несколько меньшего, чем исходный. Это наблюдение также подтверждает высказанное ранее предположение о том, что эффект внутриклеточного закисления зависит от физиологического состояния клетки.
Изменения внеклеточного рН около внутреннего сегмента в диапазоне 7,1-7,8 не влияли нвеличину фототока. Этот факт может объясняться по крайней мере двумя причинами. Либо регуляция внутриклеточного рН во внутреннем сегменте настолько эффективна, что относительно небольшие изменения внеклеточного рН на нее не влияют, либо на фототрансдукцию оказывают влияние только значительные изменения внутриклеточного рН.
В предыдущей главе мы показали, что замена хлорида на глюконат приводит к внутриклеточному защелачиванию клетки, так как локализованный в клетке НСО3/Сl-обменник увеличивает концентрацию бикарбоната внутри клетки из-за возникновения градиента хлора. Следуя той же логике в экспериментах на внутреннем сегменте, мы заменили 95 мМ NаСl на эквимолярное количество глюконата натрия. Однако такая замена не приводила к значительному и постоянному увеличению фототока, как это следовало бы ожидать.
В целом, можно заключить, что незначительные изменения внутриклеточного рН ВС, вызываемые как изменениями внеклеточного рН, так и заменой хлора на глюконат, по-видимому, носят локальный характер и не приводят к изменению внутрикле-точного рН в наружном сегменте, где и происходят процессы фототрансдукции.
Наружный сегмент. Переходные процессы при изменении рН. Либман показал, что снижение экстраклеточного рН вокруг наружного сегмента фоторецепторной клетки уменьшает темновой ток в течение 1 с. Хотя эти результаты и были интерпретированы в рамках предположения об изменении внутриклеточного рН, их нельзя считать окончательными. В этих экспериментах раствор омывал клетку в относительно небольшие промежутки времени (не более нескольких десятков секунд). В силу этой причины темновой ток полностью подавлялся только при рН 3,5.
На рис. 6.10 представлены результаты эксперимента, в котором внеклеточный рН изменялся сначала от 7,5 до 7,2, а затем - до 7,8.
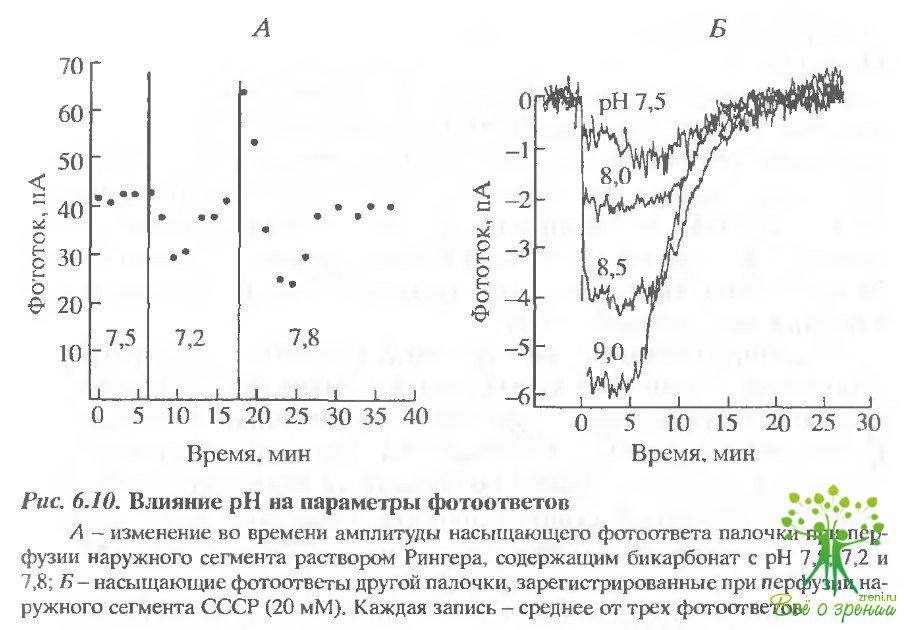
Тот факт, что фототок всегда восстанавливался до первоначального значения, говорит о том, что в клетке присутствует механизм с обратной связью. Однако даже в этом случае можно создать ситуацию, когда мощность механизма, регулирующего рН. недостаточна для его поддержания на постоянном уровне. На том же рисунке представлены результаты экспериментов, в которых в перфузат добавлялся ионофор СССР (20 мМ), который соз7ает большую протонную проводимость. В этом случае даже в стационарном состоянии удается наблюдать увеличение насыщающего фотоответа при увеличении внеклеточного рН.
Влияние амилорида на наружный сегмент фоторецептора. Амилорид. воздействуя на наружный сегмент зрительной клетки (рис. 6.11), существенно подавляет насыщающий фототок. Эффект слабо обратим.
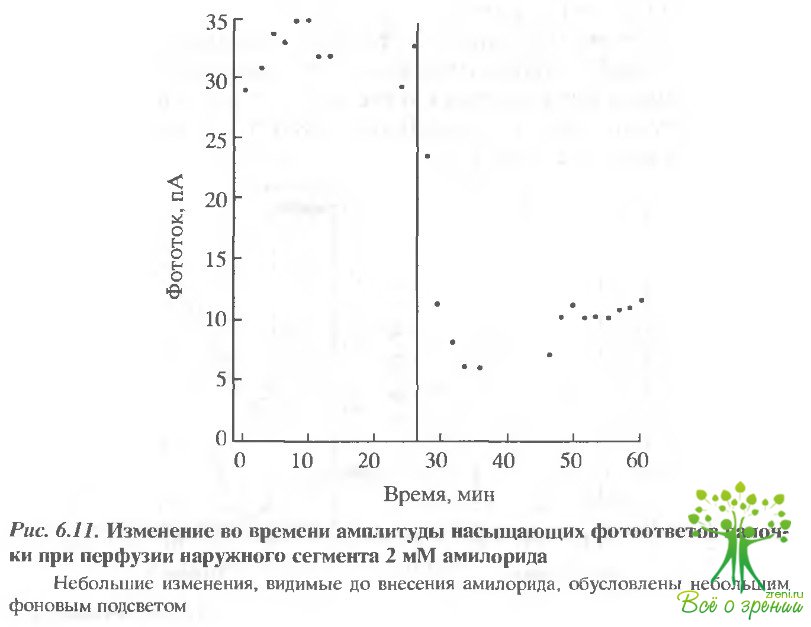
Максимальная величина восстановления фототока после отмывания амилорида не превышала 50%. Воздействие амилорида существенно увеличивалось в тех случаях, когда эксперимент проводился на фоне подсветки, которая увеличивает постоянный гидролиз цГМФ и, следовательно, увеличивает скорость образования Н+. Это подтверждает тот факт, что значительная часть эффекта обусловлена именно воздействием амилорида на Nа+/Н+-обмен.
Следует отметить, однако, что действие амилорида обычно обратимо. Амилорид и его производные не очень специфичны, и в зрительной клетке могут воздействовать как на цГМФ-регулируемые каналы, так и на Nа+/Са2+-обменник.
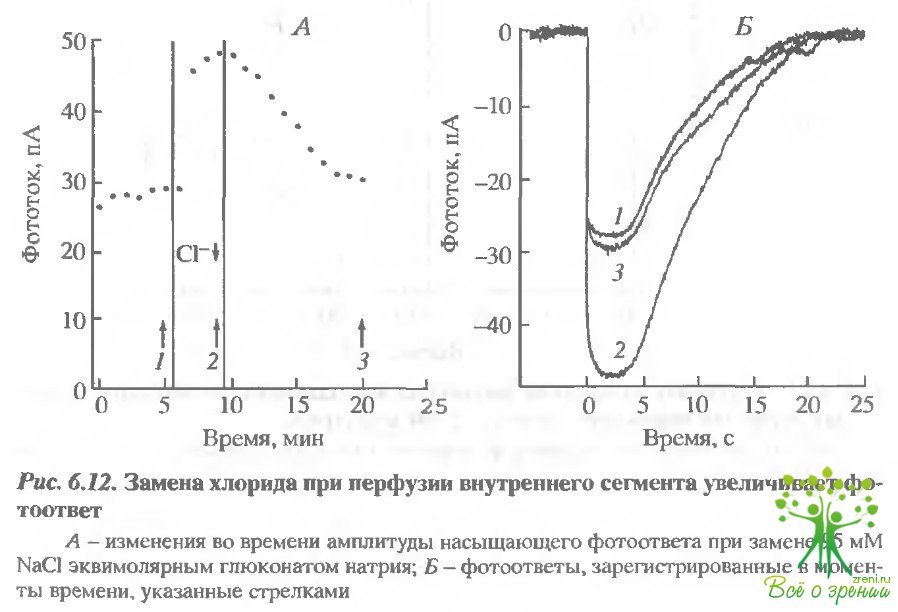
Транспорт бикарбоната е наружном сегменте. В том случае, когда в пипетку засасывался внутренний сегмент, а наружный омывался раствором, содержащим ДИДС (рис. 6.12), амплитуда фототока всегда возрастала (в среднем на 18% по семи клеткам). Если предполагать что Nа+/Н+-обменник поддерживает рН в состоянии термодинамического равновесия, то НСО3/Сl-обменник удаляет из клетки бикарбонат и, следовательно, должен закислять клетку.
Эта гипотеза была подтверждена экспериментами двух типов.
Замена хлора на глюконат приводит к увеличению фотоответа. При удалении хлора направление работы НСО3/С1-обменника должно измениться на противоположное, и это должно привести к защелачиванию внутриклеточной среды. Действительно, в том случае, когда 95 мМ NaС1 замещались на глюконат натрия (см. рис. 6.12), амплитуда фотоответа возрастала в среднем на 26% (14-53% на 14 клетках). Аналогичный эффект был получен и в том случае, когда вместо глюконата натрия использовался сульфонат натрия, Если эффект замены хлора обусловлен НСО3/С1-обменом, то он должен быть чувствителен к ДИДС. Эти эксперименты, однако, довольно трудно интерпретировать, так как ДИДС сам по себе влияет на фотоответ. Типичные результаты такого рода экспериментов представлены на рис. 6.13. В этих экспериментах изменения фототока, индуцируемые заменой хлорида на глюконат, составляли 23% в отсутствие ДИДС и 13% - в присутствии ДИДС. Трудность интерпретации состоит в том, что в присутствии ДИДС фотоответ и так значительно увеличен, так что его дальнейший рост, индуцированный заменой хлорида, может быть затруднен по этой причине. ДИДС существенно увеличивает фототок в присутствии моненсина.


Если бикарбонат удаляется из клетки при посредстве НСО3/Сl-обменника, то эффект должен возрастать при возрастании скорости удаления Н+ из клетки. Чтобы достичь этого, мы добавили в среду искусственный Na+/Н+-обменник моненсин (0,1 мМ). Добавление моненсина сначала временно увеличило амплитуду фотоогвета (рис. 6.14), так как интенсификация Nа+/Н+ привела к внутриклеточному защелачиванию. Последующее добавление ДИДС приводило к увеличению фотоответа на менее чем на 80%. Эффект, однако, как и в других случаях, был переходным и плохо обратимым.
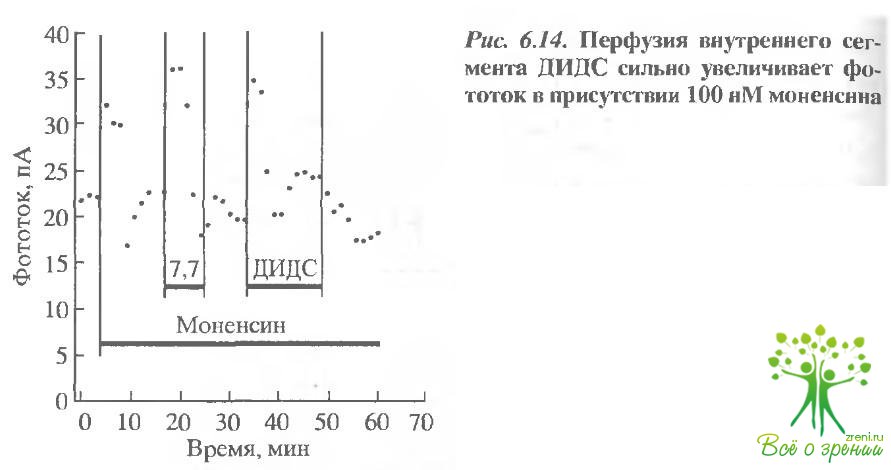
В четырех подобных экспериментах эффект ДИДС составлял в среднем 57% (в аналогичных условиях возрастание фототока при изменении рН от 7,5 до 7,7 составляло 46%). Эта величина существенно больше увеличения фототока, индуцируемого ДИДС в отсутствие моненсина, и таким образом эксперименты подтверждают наши предположения о роли НСО3/Сl-обменника в поддержании рН.
↑ Фототрансдукция в палочках сетчатки лягушки модулируется бикарбонатом
Влияние ацетазоламида и ДИДС. В данном разделе рассмотрен вопрос о том, существуют ли в сетчатке позвоночных механизмы. способные модулировать процессы трансдукции путем регуляции внутриклеточного pH и таким образом регулировать световую чувствительность зрительных клеток. Известно, что в органах и тканях регуляция pH тесно связана с метаболизмом С02Особенно это важно в нервной системе, где капилляры часто пространственно удалены от нервных тканей, и роль клеток, транспортирующих СО2, выполняют астроциты. Как известно, сетчатка эффективно снабжается кровью, поэтому механизмы метаболизма С02 и H могут существенно влиять на работу ее элементов, в частности фоторецепторных клеток.
Мюллеровские клетки обладают несколькими чертами, которые позволяют предположить их важную роль в метаболизме С02. Это специализированные астроциты, которые пронизывают сетчатку от дистальной до апикальной части. Они занимают существенную ее часть по объему и находятся в контакте с клетками трех ядерных слоев. В различных группах позвоночных большая часть карбоангидразы (КА) сетчатки локализована именно в Мюллеровских клетках, тогда как фермент отсутствует в большинстве нейронов, включая палочки. Это предполагает участие в транспорте С02, где эффективность зависит от быстрого КА-катализируемого гидрирования.
Задачей описанных выше экспериментов было выяснить важность такого транспорта и показать, что он должен быть блокирован или несколько замедлен при ингибировании активности КА. Для образования двуокиси углерода необходимо снабжение клетки ионами водорода, тогда как С02 входит в клетку беспрепятственно и после гидратации в основном диссоциирует на Н+ и НСО3 . В противовес этому многие нейроны и глиальные клетки имеют НСО3-зависимый обменный механизм, регулирующий внутриклеточный pH. Подкисление палочек уменьшает их темновой ток и содержание цГМФ, a увеличение С02-НС03, как было заявлено, имеет совершенно противоположный эффект. Поскольку последние работы включали относительно большие изменения буферной емкости, необходимо прежде всего было выяснить, действительно ли С02 и НСО; как таковые увеличивают темновой ток и концентрацию цГМФ. Это оказалось действительно так, и позволило предположить, что существует НСО зависимый механизм регуляции рН. Такие анионные обменники обычно ингибируются ДИДС, который мы использовали как фармакологический инструмент. Результаты свидетельствуют, что, во-первых, в палочках действительно существует НСО3 -регулируемый, ДИДС-зависимый механизм поддержания внутриклеточного рН и, во-вторых, действительно происходит существенное, кат авизируемое КА, удаление С02-НСО -Н+ из сетчатки. Подобно другим астроцитам, в этом случае, по-видимому, включается выброс протона Мюллеровскими клетками со стороны, обращенной к склере, что приводит к подкислению в дистальной части палочек.
Статья из книги: Молекулярные механизмы зрительной рецепции | Каламкаров Г.Р., Островский М.А..

Комментариев 0