Глаз | Часть 2

Содержание:
Описание

Здесь мы продолжим обсуждать тему строения и функций глаз, начатых в предыдущей теме.
↑ Размеры рецептивных полей
Попробуем теперь соотнести события, развертывающиеся на сетчатке, с повседневным зрительным восприятием внешнего мира. У разных ганглиозных клеток рецептивные поля различны по величине. Особенно заметным и регулярным образом изменяются размеры центров рецептивных полей: они минимальны в центральной ямке сетчатки, где острота зрения, или способность различать небольшие объекты, выше всего; чем дальше от центральной ямки, тем поля становятся все крупнее, а острота зрения соответственно понижается.
Величину рецептивного поля можно измерять двумя способами. Проще всего указывать его размеры непосредственно на сетчатке. Недостаток такого способа в том, что с обыденной точки зрения он не будет достаточно осмысленным. Вместо этого можно измерять величину проекции рецептивного поля на внешний мир, например его диаметр на экране, перед которым находится животное; при этом, однако, нужно указывать, как далеко от глаз расположен экран. Во избежание этих трудностей лучше, пожалуй, выражать величину рецептивного поля углом, под которым само животное будет видеть проекцию поля на экран, как показано на рис. 29. Мы рассчитываем этот угол в радианах путем простого деления диаметра поля на расстояние до экрана, но я буду пользоваться градусами путем пересчета по формуле: (радианы х л)/180. Один миллиметр на сетчатке человека примерно соответствует углу 3,5 градуса. При расстоянии до экрана 135 см одному градусу соответствуют на экране 2,5 см. Луна и Солнце, видимые с Земли, имеют почти одинаковые размеры в полградуса.
У обезьян наименьшие из измеренных до сих пор центров рецептивных полей имеют диаметр около 2 угловых минут, или 10 мкм (0,01 миллиметра) на сетчатке. Эти ганглиозные клетки расположены, по-видимому, вне центральной ямки, но поблизости от нее. В самой ямке диаметр колбочек и расстояния между их центрами составляют около 2,5 мкм; это хорошо согласуется с данными об остроте нашего зрения — мы способны различить две точки при видимом расстоянии между ними всего лишь 0,5 угловых минут. Кружок диаметром 2,5 мкм на сетчатке (угол 0,5 минуты) соответствует 25-центовой монетке, видимой с расстояния около 150 метров.
На дальней периферии сетчатки центры рецептивных полей состоят из тысяч рецепторов и могут иметь диаметр в 1 градус и более. Таким образом, по мере удаления от центральной ямки отмечается явно не случайное хорошо согласованное изменение трех величин: падает острота зрения, возрастает число рецепторов, участвующих в отдельных прямых путях (к биполярам и далее к ганглиозным клеткам), и увеличивается диаметр центров рецептивных полей. Это помогает нам понять значение прямых и непрямых путей от рецепторов к ганглиозным нейронам, так как дает веские основания для вывода, что центр рецептивного поля определяется прямым путем, а антагонистическая периферия — непрямым путем и что острота зрения лимитируется прямыми путями. Для получения дополнительных данных в пользу такого вывода нужны были сведения об активности других клеток сетчатки; об этом пойдет речь в следующих разделах.

↑ Фоторецепторы
Прошло немало лет, прежде чем был достигнут существенный прогресс в физиологии рецепторов, биполяров, горизонтальных и амакриновых клеток. Тому было множество причин: пульсация сосудов постоянно мешала попыткам удерживать микроэлектрод в одиночной клетке или рядом с ней; рецепторы, биполяры и горизонтальные клетки не генерируют импульсов, поэтому регистрация намного меньших градуальных потенциалов требует применения внутриклеточных методик; трудно с уверенностью сказать, в клетке какого типа (или рядом с какой клеткой) находится электрод.
Некоторые из этих затруднений можно преодолеть надлежащим выбором животного; например, сетчатки холоднокровных позвоночных способны выживать, будучи извлечены из глаза и погружены в солевой раствор, насыщенный кислородом, и при этом отсутствие кровообращения исключает пульсацию артерий; у протея (род крупных саламандр) очень большие клетки, их активность легко регистрировать; рыбы, лягушки, черепахи, кролики и кошки — все эти животные имеют свои преимущества при исследованиях того или иного типа, поэтому при изучении физиологии сетчатки использовались разные виды. Трудность при работе с таким большим числом видов состоит в том, что детали организации сетчатки могут заметно различаться у разных животных. Кроме того, наши представления о сетчатке приматов, реакции которой трудно регистрировать, до недавнего времени в значительной мере основывались на результатах, полученных на других видах. Однако по мере преодоления технических трудностей ускоряется и прогресс исследований на приматах.
В последние годы изучение реакции палочек и колбочек на свет очень сильно продвинулось вперед, и появилось ощущение, что мы начинаем понимать, как они работают.
Палочки и колбочки различаются во многих отношениях. Наиболее важно различие в их относительной чувствительности: палочки чувствительны к очень слабому свету, колбочки требуют намного более яркого освещения. Я уже описывал различия в их распределении по сетчатке, наиболее заметное из них — отсутствие палочек в центральной ямке. Они различны и по форме: палочки длинные и тонкие, а колбочки короткие и конусообразные. Как палочки, так и колбочки содержат светочувствительные пигменты. Во всех палочках пигмент один и тот же; колбочки делятся на три типа, каждый из них со своим особым зрительным пигментом. Эти четыре пигмента чувствительны к различным длинам световых волн, и в случае колбочек эти различия составляют основу цветового зрения.
Под воздействием света в рецепторах происходит процесс, называемый выцветанием. В этом процессе молекула зрительного пигмента поглощает фотон — единичный квант видимого света — и при этом химически превращается в другое соединение, хуже поглощающее свет или, быть может, чувствительное к другим длинам волн. Практически у всех животных, от насекомых до человека, и даже у некоторых бактерий этот рецепторный пигмент состоит из белка, к которому присоединена небольшая молекула, близкая к витамину А; она и представляет собой химически трансформируемую светом часть. Благодаря главным образом работам Джорджа Уолда из Гарварда, проведенным в 50-х годах, нам теперь многое известно о химии выцветания и последующего восстановления зрительных пигментов.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]

Большинство обычных сенсорных рецепторов — химических, температурных или механических — деполяризуется в ответ на соответствующий стимул, т. е. они реагируют на возбуждающий стимул так же, как обычные нейроны; деполяризация ведет к высвобождению медиатора из аксонных окончаний (часто, как и в случае зрительных рецепторов, это не приводит к возникновению импульсов, вероятно из-за очень малой длины аксона). У беспозвоночных, от усоногих раков до насекомых, световые рецепторы ведут себя таким же образом, и до 1964 года предполагалось, что аналогичный механизм — деполяризация под влиянием света — действует также в палочках и колбочках позвоночных.
В 1964 году японскому нейрофизиологу Цунео Томита, работавшему в университете Кейо в Токио, впервые удалось ввести микроэлектрод в колбочки сетчатки рыбы и получить столь неожиданный результат, что у многих современников он вызывал вначале серьезные сомнения. В темноте потенциал на мембране колбочки оказался необычайно низким для нервной клетки: приблизительно 50 милливольт вместо обычных 70. При освещении колбочки этот потенциал возрастал — мембрана гиперполяризовалась — в противоположность тому, чего следовало бы ожидать.
В темноте фоторецепторы позвоночных явно больше деполяризованы (имеют более низкий мембранный потенциал), чем обычные нервные клетки в состоянии покоя, и деполяризация вызывает непрерывное высвобождение медиатора из окончаний их аксонов — в точности так, как это происходит в обычных рецепторах при стимуляции. Свет, повышая потенциал на мембране рецепторной клетки (т. е. гиперполяризуя ее), уменьшает выделение медиатора. Таким образом, стимуляция, как это ни странно на первый взгляд, выключает рецепторы. Открытие Томита помогает нам объяснить, почему волокна зрительного нерва у позвоночных столь активны в темноте: спонтанную активность проявляют именно рецепторы; многие биполярные и ганглиозные клетки, вероятно, делают попросту то, что им диктуют рецепторные клетки.
В последующие десятилетия главные задачи состояли в том, чтобы выяснить, как свет вызывает гиперполяризацию рецептора и в особенности каким образом выцветание всего одной молекулы зрительного пигмента под действием одного фотона может привести в палочке к измеримому изменению мембранного потенциала. В настоящее время оба процесса достаточно хорошо поняты. Гиперполяризация на свету вызывается перекрытием потока ионов.
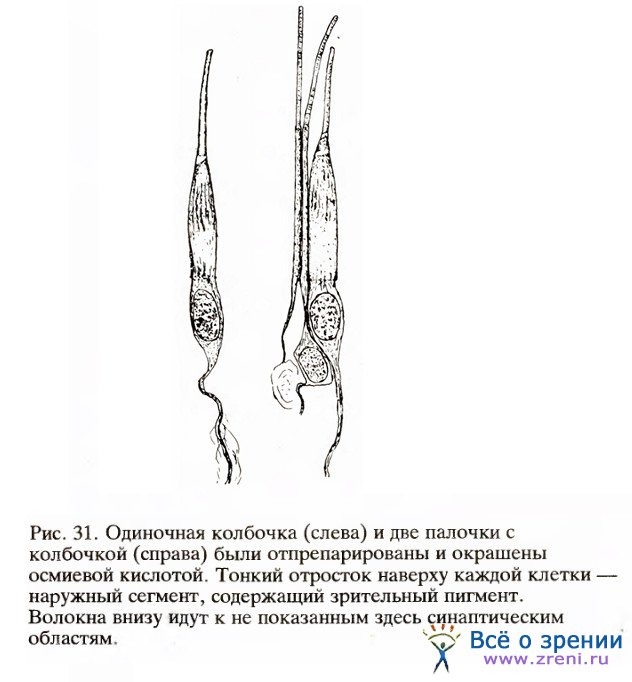
В темноте часть рецепторной мембраны более проницаема для ионов натрия, чем остальная мембрана. Поэтому ионы натрия непрерывно входят здесь в клетку, а где-то в другом месте ионы калия выходят наружу. Поток ионов в темноте, или темновой ток, открыли в 1970 году Уильям Хейгинс, Ричард Пенн и Шуко Йосиками в Национальном институте артрита и нарушений метаболизма в Бетезде. Он вызывает деполяризацию покоящегося рецептора и тем самым — его постоянную активность. В результате выцветания зрительного пигмента на свету поры для натрия закрываются, темновой ток уменьшается и степень деполяризации мембраны становится меньше, т. е. клетка гиперполяризуется. Ее активность (высвобождение ею медиатора) ослабевает.
В настоящее время в результате работ Евгения Фесенко с сотрудниками в Академии наук в Москве, Дениса Бейлора в Стэнфордском университете, Кин-Вай Яу в Техасском университете и других мы намного ближе подошли к пониманию связи между выцветанием пигмента и закрытием натриевых пор. Например, очень трудно было представить себе, как выцветание единственной молекулы могло бы привести к закрытию миллионов пор, необходимому для наблюдаемых изменений потенциала. В настоящее время выяснилось, что поры в рецепторе открываются с помощью молекул вещества, называемого циклическим гуанозинмонофосфатом (цГМФ). Выцветание молекулы зрительного пигмента приводит к целому каскаду событий. Белковая часть обесцвеченной молекулы пигмента активирует большое число молекул фермента трансдуцина, а каждая из них в свою очередь инактивирует сотни молекул цГМФ, обычно участвующих в открытии пор. Так в результате выцветания одной молекулы пигмента закрываются миллионы пор.
Все это позволяет объяснить ряд явлений, бывших ранее загадочными. Во-первых, давно известно, что человек, адаптировавшийся к полной темноте, способен увидеть такую слабую вспышку света, при которой ни один рецептор не может получить более одного фотона. Как показывают расчеты, для ощущения вспышки нужно, чтобы в короткий промежуток времени около шести близко расположенных палочек были стимулированы фотонами. Теперь становится понятно, как одиночный фотон может возбудить палочку и заставить ее генерировать сигнал достаточной силы.
Во-вторых, мы теперь можем объяснить неспособность палочек реагировать на изменения освещенности, если свет уже достаточно ярок. По-видимому, чувствительность палочек столь высока, что при сильной освещенности, например при солнечном свете, все натриевые поры закрыты, и дальнейшее усиление света может не давать никакого дополнительного эффекта. Тогда мы говорим, что палочки насыщены.
Быть может, спустя несколько лет студенты-биологи будут видеть во всей этой повести о рецепторах просто еще одну вещь, которую нужно выучить. Надеюсь, однако, что этого не произойдет. Чтобы полностью оценить ее значение, надо было потратить многие годы, гадая о том, каким образом могут работать рецепторы, а затем внезапно — меньше чем за десяток лет — в результате эффектных исследований решить эту проблему. Волнение по этому поводу еще не стихло.
↑ Биполярные и горизонтальные клетки
Горизонтальные и биполярные клетки вместе с амакриновыми клетками расположены в среднем слое сетчатки. Биполярные клетки занимают в сетчатке стратегическую позицию, поскольку все сигналы, возникающие в рецепторах и поступающие к ганглиозным клеткам, должны пройти через них. Это означает, что они входят в состав как прямых, так и непрямых путей. В отличие от этого горизонтальные клетки входят только в непрямые пути. Как можно видеть на рис. 33, горизонтальные клетки встречаются намного реже биполярных, которые в целом преобладают в среднем слое.
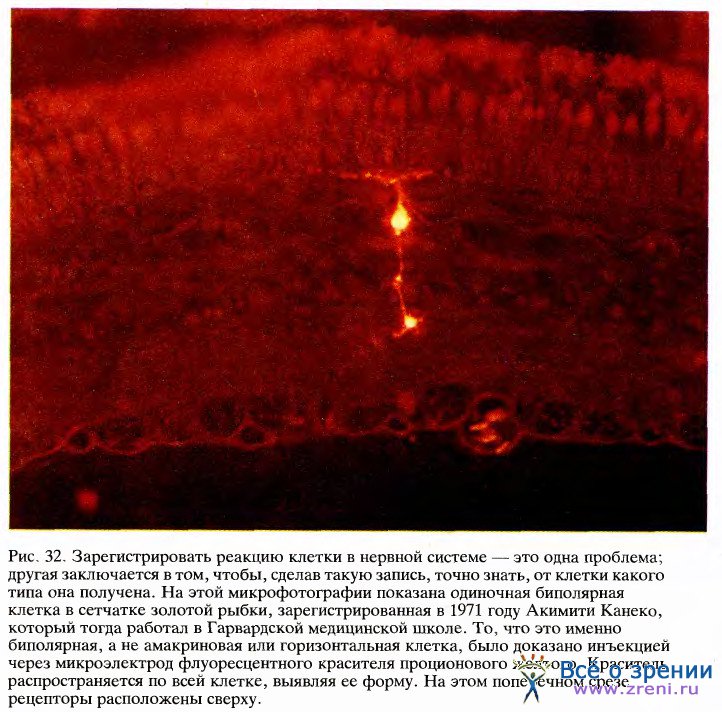
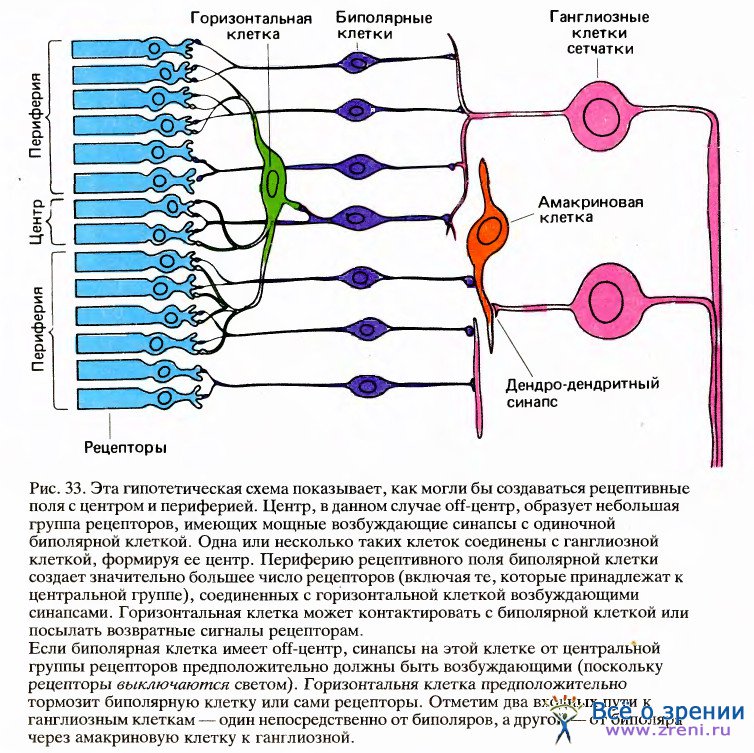
До того как удалось зарегистрировать активность биполярных клеток, никто не знал, обладают ли их рецептивные поля, как у ганглиозных клеток, центром и периферией и делятся ли эти поля на два типа — с оп-центром и с off-центром. Утвердительный ответ на эти вопросы, почти несомненно, означал бы, что открытая Куффлером организация ганглиозных клеток пассивно отражает организацию биполяров. Данные о том, что рецептивные поля биполярных клеток действительно имеют центр и периферию и представлены двумя типами, были впервые получены путем внутриклеточной регистрации активности Джоном Даулингом и Фрэнком Верблином в Гарвардских биологических лабораториях и Акимити Канеко в Гарвардской медицинской школе.
Следующий вопрос — как устроены эти рецептивные поля. Для ответа на него мы должны начать с изучения связей между рецепторами, биполярами и горизонтальными клетками.
Биполярная клетка посылает по направлению к рецепторам единственный дендрит. Он либо образует синапс с одним рецептором (всегда с колбочкой), либо расщепляется на веточки, синаптически контактирующие более чем с одним рецептором. Если с одним биполяром связаны два или несколько рецепторов, они совместно занимают сравнительно малый участок сетчатки. В любом случае эти рецепторы должны составлять центр рецептивного поля, поскольку занимаемая ими площадь соответствует центру поля по величине. Возникает еще один вопрос: каковы синапсы между рецепторами и биполярными клетками — возбуждающие они, тормозные или обоих типов?
Биполярные клетки, подобно рецепторам и горизонтальным клеткам, не генерируют импульсов, но мы и здесь будем говорить об оп-реакции, подразумевая деполяризацию в ответ на световой стимул и соответственно усиленное выделение медиатора в выходных синапсах, и об off-реакции, разумея гиперполяризацию и уменьшение выброса медиатора. Что касается входных синапсов, передающих биполярам сигналы от рецепторов, то у биполяров с off-центром они должны быть возбуждающими, поскольку сами рецепторы выключаются (гиперполяризуются) светом; у биполяров с оп-центром входные синапсы должны быть тормозными. Чтобы понять, почему это так (если вас, как и меня самого, это путает), следует лишь представить себе влияния малого светового пятна. Рецепторы активны в темноте, а свет, вызывая гиперполяризацию, уменьшает их активность. Если синапс возбуждающий, биполяр будет активироваться в темноте, а инактивироваться на свету; если же синапс тормозный, биполяр в темноте тормозится, а свет, выключая рецептор, снимает это торможение, т. е. биполярная клетка активируется (это действительно нелегко сразу понять).
Является ли рецепторно-биполярный синапс возбуждающим или тормозным, зависит либо от выделяемого рецептором медиатора, либо от типа каналов в постсинаптической мембране биполярной клетки. В настоящее время никто не думает, что один рецептор высвобождает два медиатора, и имеется много данных в пользу того, что биполяры двух типов имеют различные рецепторные молекулы.
Прежде чем обсуждать, как создается периферия рецептивного поля биполярного нейрона, мы должны рассмотреть горизонтальные клетки.
Горизонтальные клетки важны потому, что они, видимо, по меньшей мере частично ответственны за периферию рецептивных полей ганглиозных клеток сетчатки; они составляют ту часть непрямого пути, о которой мы знаем больше всего. Это крупные клетки, и они принадлежат к числу самых удивительных в нервной системе. Их отростки тесно контактируют с окончаниями многих фоторецепторов, которые распределены по площади, размеры которой велики по сравнению с участком, непосредственно связанным с одиночной биполярной клеткой. Каждый рецептор контактирует с обоими типами клеток второго порядка — биполярными и горизонтальными.
Имеется несколько подтипов горизонтальных клеток, которые могут сильно различаться у разных животных; их самое необычное свойство, общее с амакриновыми клетками, — это отсутствие чего-либо похожего на типичный аксон. Прочитав несколько упрощенное описание нервных клеток в предыдущей главе, вы с удивлением можете спросить: каким образом нерв без аксона будет передавать информацию другим нейронам? Когда начали использовать в нейроанатомии электронный микроскоп, вскоре выяснилось, что в некоторых случаях дендриты могут быть пресинаптическими отростками и образуют синапсы на других нейронах, обычно на их дендритах. (И наоборот, иногда окончания аксонов могут быть постсинаптическими структурами по отношению к другим аксонам, которые на них оканчиваются. Отростки, отходящие от тел горизонтальных и амакриновых клеток, могут, по-видимому, выполнять функции как аксонов, так и дендритов.
Своеобразны и синапсы, образуемые горизонтальными клетками с рецепторами: у них отсутствуют электронно-микроскопические признаки, обычно указывающие, в каком направлении передаются сигналы. Ясно, что рецепторы доставляют информацию горизонтальным клеткам через возбуждающие синапсы, поскольку в большинстве случаев горизонтальные клетки, подобно рецепторам, гиперполяризуются (т. е. выключаются) светом. Менее ясно, куда эти клетки посылают свои выходные сигналы. Известно, что у некоторых животных, например у черепах, они передают информацию обратно рецепторам; у других видов они образуют синапсы с дендритами биполярных клеток и, несомненно, передают им информацию; о приматах мы не располагаем какими-либо сведениями. Итак, горизонтальные клетки получают входные сигналы от рецепторов; их выход пока точно не известен, но он направлен либо к рецепторам, либо к биполярным клеткам, либо к тем и другим.
То, что рецепторы передают горизонтальным клеткам информацию с относительно обширного участка сетчатки, означает, что рецептивные поля этих клеток должны быть большими. Так оно и есть: их величина примерно соответствует размерам рецептивных полей биполярных или ганглиозных клеток, включая центр вместе с периферией. Они однородны, и освещение их в любом месте вызывает гиперполяризацию, которая тем сильнее, чем больше световое пятно. Многие данные указывают на то, что горизонтальные клетки ответственны за периферию рецептивных полей биполярных клеток; других кандидатов на эту роль, в сущности, нет, поскольку это единственные клетки, связанные с рецепторами на столь обширном пространстве. Там, где горизонтальные клетки непосредственно контактируют с биполярами, синапсы к on-биполярам должны быть возбуждающими (ввиду тормозящего влияния света на периферию), а к off-биполярам — тормозными. Если же влияние осуществляется через рецепторы, синапсы должны быть тормозными.
Резюмируем сказанное выше. Биполярные клетки имеют рецептивные поля с центром и периферией. Реакция центра определяется прямым входом от небольшой группы рецепторов; периферию определяет непрямой путь от более обширной области рецепторов, связанных с горизонтальными клетками, которые, вероятно, в свою очередь передают сигналы биполярам. Непрямой путь, кроме того, мог бы быть результатом обратной связи от горизонтальных клеток, тормозящей рецепторы.
↑ Амакриновые клетки
Эти клетки удивительно разнообразны по форме и используют необычайно большое число нейромедиаторов, которых может быть более двадцати. Все амакриновые клетки имеют ряд общих особенностей. Во-первых, тела их расположены в среднем слое сетчатки, а отростки — в синаптической зоне между этим слоем и ганглиозными клетками; во-вторых, они образуют связи и с биполярными, и с ганглиозными клетками и таким образом создают между теми и другими альтернативный, непрямой путь; и наконец, у них нет аксонов, но зато их дендриты способны к образованию пресинаптических окончаний на других клетках.
Амакриновые клетки, вероятно, выполняют много различных функций, большей частью неизвестных. Один их тип, по-видимому, участвует в специфических реакциях на движущиеся объекты, обнаруженных в сетчатке лягушки и кролика; другой тип участвует в создании пути, связывающего ганглиозные клетки с теми биполярами, которые имеют входы от палочек. Нет данных о том, чтобы амакрины участвовали в организации центра и периферии рецептивных полей ганглиозных клеток, но нельзя полностью исключить такую возможность. Функции большинства их форм остаются невыясненными; следует, вероятно, прямо сказать, что анатомические сведения об амакриновых клетках в целом значительно богаче физиологических.
↑ Связи между биполярными и ганглиозными клетками
Мы видели, что основные особенности рецептивных полей ганглиозных клеток проявляются уже у биполярных клеток. Это оставляет открытым вопрос о том, какие преобразования информации происходят между биполярами и ганглиозными клетками. Вряд ли тут совсем ничего не происходит, если судить по сложности синаптического слоя между средним слоем и слоем ганглиозных клеток, поскольку мы часто встречаем здесь конвергенцию на прямом пути от биполярных клеток к ганглиозным, а также включение амакриновых клеток, функция которых не вполне понятна.
Вероятно, все синапсы между биполярными и ганглиозными клетками возбуждающие; это означает, что биполярные клетки с оn-центром передают сигналы ганглиозным клеткам с оп-центром, а биполяры с off-центром — ганглиозным клеткам с off-центром. Это упрощает схему связей: ведь могло быть так, например, что клетки с оп-центром через тормозные синапсы воздействовали бы на клетки с off-центром. Ну что же, и на том спасибо!
До 1976 года не было известно, различаются ли по форме клетки с оп-центром и с off-центром, но в этом году Ральф Нелсон, Хельга Колб и Эдвард Фамильетти из Национальных институтов здоровья в Бетезде внутриклеточно зарегистрировали реакции ганглиозных клеток кошки, идентифицировав их как клетки с on- или off-центром, а затем через микроэлектрод ввели вещество, окрашивающее все дендритное дерево. Сравнив разветвления дендритов у клеток двух типов, они увидели четкое различие: две совокупности дендритов оканчивались на двух отдельных субслоях в пределах синаптической зоны между средним слоем и слоем ганглиозных клеток. Дендриты клеток с off-центром всегда оканчивались ближе к среднему слою сетчатки, а дендриты клеток с оп-центром — дальше от него. К тому времени в другой работе уже было показано, что биполярные клетки двух классов, образующие с рецепторами синапсы разной формы, различаются также и расположением своих аксонных окончаний: у одних аксоны оканчиваются там, где кончаются дендриты ганглиозных клеток с оп-центром, а у других — там, где кончаются дендриты клеток с off-центром. Таким образом, стало возможным реконструировать весь путь от рецепторов к ганглиозным клеткам для обеих систем — с on-и с off-центрами.
В результате был установлен неожиданный факт: на прямом пути именно система с off-центрами имеет возбуждающие синапсы на каждом уровне — от рецепторов к биполярам и от биполяров к ганглиозным клеткам, тогда как в системе с on-центрами синапсы между рецепторами и биполярами тормозные.
Разделение биполярных и ганглиозных клеток на категории с on- и с off-центром наверняка должно иметь свои корреляты в восприятии. Клетки с off-центром отвечают на темное пятно точно таким же образом, как клетки с оп-центром — на светлое пятно. Если наличие двух совокупностей клеток, отвечающих на темные и на светлые пятна, кажется нам неожиданным, то это, возможно, результат совершенно справедливых утверждений физиков, что темнота — это отсутствие света. Но темнота представляется нам вполне реальной, и теперь мы, видимо, находим биологическую основу этой реальности. Черное для нас так же реально, как и белое. Шрифт книги, которую вы сейчас читаете, в конце концов тоже черный.
Совершенно так же обстоит дело с теплом и холодом. Из курса физики мы с удивлением узнаем, что холод — это всего лишь отсутствие тепла, а между тем он кажется нам точно такой же реальностью, особенно если вы выросли, как это было со мной, в морозном Монреале. Наша интуиция подтверждается, когда мы узнаём, что в коже имеются температурные рецепторы двух типов: одни реагируют на повышение температуры, другие — на ее понижение. Поэтому с биологической точки зрения холод столь же реален, как и тепло.
Во многих сенсорных системах используются оппонентные пары: тепло/ холод, черное/белое, поворот головы налево/направо, желтое/синее и красное/зеленое. Причина этого, вероятно, в характере нервной импульсации и ее изменений. В принципе можно представить себе нервы с частотой разрядов, поддерживаемой на некотором достаточно высоком уровне, скажем 100 в секунду, и потому способных в ответ на оппонентные стимулы либо снижать, либо повышать частоту импульсов — вниз до нуля, а вверх, например, до 500 в секунду. Но поскольку на импульсы расходуется метаболическая энергия (весь входящий в нерв натрий должен откачиваться наружу), то для нервных клеток в состоянии покоя более экономным, вероятно, будет молчание или импульсация с низкой частотой, а организму выгодно иметь для каждой данной модальности две отдельные группы клеток, одна из которых разряжается при уменьшении, а другая при увеличении силы стимула.
↑ Значение рецептивных полей с центром и периферией
Зачем эволюции понадобилось создавать столь любопытные образования, как рецептивные поля с центральной и периферической зонами? Иными словами, какую пользу они приносят животному?
Отвечать на столь глубокие вопросы всегда нелегко, но мы можем попытаться высказать ряд правдоподобных соображений. Сообщения, которые глаз посылает мозгу, могут быть очень мало связаны с абсолютной освещенностью сетчатки, так как ганглиозные клетки сетчатки плохо реагируют на изменения силы рассеянного света. О чем действительно сигнализирует клетка, так это о результате сравнения количества света, падающего на определенный участок сетчатки, с его средним количеством в ближайшем окружении.
Мы можем проиллюстрировать это сравнение следующим экспериментом. Найдем сначала клетку с оп-центром и картируем ее рецептивное поле. Затем, равномерно осветив экран слабым постоянным фоновым светом, начнем включать и выключать пятнышко, точно заполняющее центр поля, начиная со столь слабого света, что его еще нельзя увидеть, и постепенно повышая интенсивность. При некоторой яркости мы начнем обнаруживать реакцию; отметим, что именно при этой яркости мы и сами начнем видеть это пятнышко. Измерив интенсивность фона и пятнышка фотометром, мы выясним, что пятнышко приблизительно на 2% ярче фона. Теперь повторим всю процедуру, начиная с фонового света в пять раз более яркого. Будем постепенно повышать интенсивность локального стимула. В какой-то момент мы снова начнем обнаруживать реакции, и опять это произойдет при такой яркости, когда мы сами будем едва замечать световое пятнышко на новом фоне. Измерив стимулирующий свет, мы найдем, что он тоже в пять раз ярче предыдущего, т. е. пятнышко снова на 2% ярче фона. Вывод таков, что как для нас, так и для клетки существенна относительная освещенность пятнышка и фона.
Неспособность клетки хорошо реагировать на что-либо кроме различий в локальной интенсивности может показаться странной, так как при рассматривании крупного равномерно освещенного пятна его внутренность представляется нам такой же яркой, как и его края. Ганглиозная клетка, если учесть ее физиологию, может передавать информацию только о границах пятна; внутренность его мы видим как однородную, поскольку ганглиозные клетки с полями, находящимися внутри пятна, не сообщают о локальных различиях в освещенности. Аргументация эта кажется достаточно убедительной, и все же мы испытываем некоторое сомнение: логика логикой, но ведь внутренность пятна видна все-таки абсолютно ясно! Мы вновь и вновь будем сталкиваться с этой проблемой в последующих главах, и нам придется признать, что нервная система часто работает по принципам, противоречащим нашей интуиции.
Рассуждая рационально, однако, мы должны согласиться, что видеть большое пятно с помощью только тех клеток, поля которых дежат на его границах (не нуждаясь в участии остальных клеток с центрами, распределенными по всему пятну), — это более экономный способ: если вы инженер, то вы, вероятно, именно так сконструировали бы соответствующую машину. В таком случае и машина, я думаю, тоже «представляла бы себе» пятно освещенным равномерно.
В одном отношении слабые реакции нейрона или их отсутствие при воздействии диффузного света не должны вызывать удивления. Каждый, кто пытался фотографировать без экспонометра, знает, насколько плохо мы можем судить об абсолютной интенсивности света. Считайте, что нам везет, если мы можем устанавливать диафрагму своего фотоаппарата с ошибкой не более чем вдвое; да и это достигается не прямой оценкой «на глаз», а лишь в результате большого опыта, позволяющего, например, отметить, что в момент съемки на небе легкая облачность и что мы находимся на открытом пространстве в тени за час до захода солнца. Однако при пространственных сравнениях, когда нужно сказать, какая из двух соседних областей ярче или темнее, мы, как и ганглиозные клетки, судим очень точно. Как уже говорилось, мы способны производить такое сравнение при различии всего в 2% — в точности так, как это могут делать наиболее чувствительные ганглиозные клетки в сетчатке обезьяны.
Наряду с эффективностью эта система обладает еще одним важным преимуществом. Большинство объектов мы видим благодаря отраженному свету таких источников, как солнце или электрическая лампочка. Несмотря на изменения освещенности, создаваемой этими источниками, наша зрительная система сохраняет удивительную способность к неизменному восприятию объектов. Ганглиозная клетка сетчатки работает именно так, что это становится возможным. Рассмотрим следующий пример: газета выглядит примерно одинаково — белая бумага, черные буквы, — читаем ли мы ее в полумраке комнаты или на пляже в солнечный день. Предположим, что в обоих случаях мы измерим входящий в наши глаза свет от белой бумаги и от одной из черных букв заголовка. Вот какие цифры я получил, когда перешел из закрытого помещения на солнце во внутренний дворик Гарвардской медицинской школы:

Сами цифры не вызывают сомнений. Снаружи свет в 20 раз ярче, чем в комнате, а черные буквы отражают приблизительно десятую долю света, отражаемого белой бумагой. Но эти цифры, когда вы видите их впервые, тем не менее поражают, так как они показывают, что черная буква на открытом воздухе посылает в наши глаза вдвое больше света, чем белая бумага при комнатном освещении. Отсюда ясно, что восприятие черного и белого не определяется самим по себе количеством отражаемого объектом света. Существенно то, каково это количество в сравнении с окружающим фоном.
Экран выключенного черно-белого телевизора в нормально освещенной комнате выглядит белым или серовато-белым. Инженеры создали электронные механизмы, которые могут сделать экран ярче, но не способны затемнить его, и независимо от того, как он выглядит в выключенном виде, никакая его часть никак не может послать после включения меньше света. Мы, однако, хорошо знаем, что телевизор может дать нам ощущение насыщенного черного цвета. Самый темный участок изображения посылает в наши глаза по меньшей мере столько же света, что и при выключенном телевизоре. Отсюда следует, что «черное» и «белое» — не просто физические понятия: это биологические термины, они отражают результат вычислений, производимых нашей сетчаткой и мозгом при обработке воспринимаемой картины.
Как мы скоро узнаем, все соображения, которые я высказал здесь о черном и белом, применимы и к цвету. Видимый цвет объекта определяется не только приходящим от него светом, но также — в столь же значительной степени, как в случае черного и белого, — и светом, приходящим от всего остального. В результате то, что мы видим, становится независимым не только от интенсивности света, но и от его спектрального состава. И опять-таки это способствует постоянству восприятия видимой картины несмотря на заметные различия в яркости и спектральном составе падающего на нее света.
↑ Заключение
Выходные сигналы от глаза после двух или трех синапсов содержат намного более усложненную информацию, чем мозаичное представление мира, кодируемое палочками и колбочками. Что мне кажется особенно интересным, так это неожиданность новых результатов; до Куффлера никто не мог бы даже предположить, что существует что-либо вроде рецептивных полей с центром и периферией и что по зрительному нерву практически не передаются сведения о таком малосущественном параметре, как уровень диффузного освещения.
Точно так же никому не удалось даже приблизительно догадаться, что же поступает на следующие уровни зрительной системы — в головной мозг. Именно эта непредсказуемость и делает мозг столь привлекательным объектом — наряду с остроумием его механизмов (после того как нам удается их раскрыть).
---
Статья из книги: Глаз, мозг, зрение | Хьюбел Д.

Комментариев 0