Амакриновые и интерплексиформные клетки

Описание

В прошлом веке амакриновые клетки были морфологически исследованы и классифицированы сначала А. С. Догелем [Dogiel, 1888а, b], затем Ramon у Cajal [1972]. А. С. Догелю принадлежит приоритет открытия находящихся среди амакринов нейронов с длинным аксоном (клетка Догеля). Описание разновидностей амакриновых клеток методом Гольджи, данное Ramon у Cajal, до сих пор остается непревзойденным. Стоит лишь упомянуть, что в сетчатке рыбы (карпа) Ramon у Cajal увидел и описал 14 разновидностей этих интересных элементов.
В обзоре по морфологии сетчатки Stell [1972] заметил, что в нашем веке амакриновые клетки почти не изучались. Однако он не совсем прав, так как в 1941 г. обстоятельное описание сетчатки приматов, данное Polyak, частично заполнило эту брешь. В настоящее время многочисленные электронно-микроскопические исследования, а также микроэлектродные электрофизиологические, придали новый интерес проблеме амакриновых клеток.
Следующие факты можно считать установленными: 1) на телах амакриновых клеток в сетчатке птиц оканчиваются центробежные волокна, идущие из мозговых центров [Dogiel, 1895; Ramon у Cajal, 1972; Cowan, Powell, 1963]; 2) отростки амакриновых клеток образуют синапсы с отростками биполярных и ганглиозных клеток [Dowling, Boycott, 1965, 1966]; 3) синаптические связи, в которые вступают отростки амакриновых клеток, могут быть ленточными [Dowling, Boycott, 1965, 1966; Allen, 1969], сериальными [Kidd, 1962], реципрокными [Dowling, Boycott, 1966]; 4) одной из функций амакриновых клеток является торможение, которое они оказывают на соседние и последующие элементы сетчатки [Зепкин, Максимов, 1964; Бызов, Полищук, Зенкин, 1970]. И. А. Утина, Г. М. Зеикин, Э. А. Лебедев [1976] обнаружили во внутренней зоне внутреннего ядерного слоя элементы, активность которых выше при раздражении движущимися стимулами. Это исследование, а также работы Н. А. Полищук [1976] показали, что величина латерального торможения зависит от движения стимула и относительного контраста. Предполагается, что механизм детекции движения формируется на уровне амакриновых клеток.
Сетчатка рыб. Как сказано выше, в сетчатке карпа 14 разновидностей амакриновых клеток: однослойные различных размеров, ветвящиеся во всех подслоях внутреннего плексиформного слоя; двухслойные, многослойные и диффузные [Ramon у Cajal, 1972]. Асимметричная амакриновая клетка также наблюдалась Ramon у Cajal и названа им диагональной.
В морфофизиологической работе Chan, Naka [1976], посвященной амакриновым клеткам сетчатки рыбы Ictalurus punctatus, обнаружено два их типа: однослойные (дающие топические реакции на раздражения) и двухслойные (с фазическими реакциями). Более детальные описания морфологии этих типов были даны ранее [Naka, Carraway, 1975], хотя авторы отказались от термина «амакриновые клетки».
Однослойные нейроны названы в этой работе звездорасходящимися (starburst) по их виду на тотальной сетчатке, они делятся на симметричные и асимметричные. Двухслойные клетки названы веретенообразными по форме тела. Кроме звездорасходящихся и веретенообразных, авторы нашли разновидность однослойного нейрона с удлиненным телом и редкими длинными деидритами, названного спагетти-типом. Диффузные клетки отсутствуют в описаниях Naka, Carra-way [1975] и Chan, Naka [1976].
В сетчатке золотой рыбки посредством селективных меток амакриновых клеток выявлено несколько их типов, отличающихся размерами тела, дендритных полей и различным расположением отростков. Диаметр дендритного поля глицинэргических клеток равняется 26 мкм, ветвление двухслойное, распространяющееся в наружных отделах внутреннего плексиформного слоя.
От ГАМК-эргических амакриновых клеток отходит один основной нисходящий отросток,ветвящийся во внутренних отделах внутреннего плексиформного слоя. Индоламииэргические (серотонинсодержащие) амакриновые клетки образуют широкое иоле в подслое 1-го внутреннего плексиформного слоя [Маге, 1982, Зак, 1984]. Каждый тип нейрона метится лишь определенным медиатором.
Отростки, метящиеся ГАМК, группируются вокруг терминалей центр-деноляризациониых биполярных клеток. Эти терминали являются и пре- и постсинаптическими к ГАМК-эргическим амакриновым клеткам.
В связи с этими исследованиями представляет интерес показать здесь амакриновые клетки, найденные нами ранее в сетчатке щуки [Школьник-Яррос, Подугольникова, Дюбина, 1973, 1975] с некоторыми дополнениями. В сетчатке щуки импрегнировались диффузные и слоистые амакриновые клетки.

У диффузного нейрона (рис. 23, е) небольшое тело, от которого отходит короткий тонкий отросток, рассыпающийся в самом наружном отделе внутреннего плексиформного слоя на множество вторичных и третичных ветвей. По ходу и на концах извилистых конечных отросточков наблюдаются небольшие пуговки. Все образование имеет вид опрокинутого веера, распространяющегося по наружной половине внутреннего плексиформного слоя. Другая разновидность нейрона является как бы переходной между слоистым и диффузным типом (рис. 23, а). Маленькое в виде перевернутой груши тело продолжается длинным отростком, спускающимся к средней трети внутреннего плексиформного слоя. От основного отростка отходит в виде арки вторичный, дающий начало множеству конечных тончайших ветвлений, по ходу которых рассыпаны крошечные пуговки на концах. Это изящное диффузное ветвление занимает примерно середину внутреннего плексиформного слоя, размещаясь горизонтально.

[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Более полно импрегнировались слоистые нейроны (рис. 23, 24, 25), среди которых однослойные (рис. 23, f, d), двухслойные (рис. 23, с, 24) и многослойные (рис. 23, c.26). У миниатюрной однослойной клетки отростки распространяются горизонтально по самому наружному отделу внутреннего плексиформного слоя (см. рис. 23). У другого однослойного нейрона (см. рис. 23, d) крупный отросток спускается к середине плексиформного слоя, где в обе стороны от него отходят также крупные и весьма длинные ветки, простирающиеся до 900 мкм, покрытые варикозностями и единичными шипиками. Двухслойные амакриновые клетки импрегнировались недостаточно полно, но по характеру ветвлений их вполне уверенно можно отнести к этой разновидности (см. рис. 23, с; 24).
Наибольший интерес в сетчатке щуки представляет клетка «b» на рис. 23. Отдельная зарисовка этого нейрона дана на рис. 26.
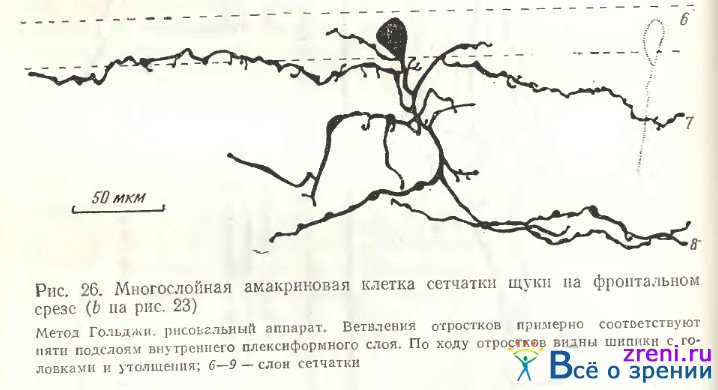
Основные отростки ветвятся по самой наружной и внутренней границам внутреннего плексиформного слоя. Однако и в центре этого слоя, и в самом внутреннем ядерном слое имеются дополнительные ветвления, так что эту клетку без всякого сомнения можно отнести к многослойным. По ходу отростков видны варикозности разной величины, а также шиповидные отросточки с типичными головками.
Кроме симметричных нейронов, наблюдаются и асимметричные (рис.27,28).

Если у двухслойного нейрона на рис.24 можно думать о неполной импрегнации, и асимметрия может быть кажущейся, то у клеток на рис. 27, 28 она действительно имеется. Длинное и мощное ветвление клетки на рис. 27 полностью обращено в одну сторону, вторичные и третичные отростки направлены точно так же. Описываемый нейрон, по всей вероятности, следует отнести к двухслойным, так как весьма крупный отросток в верхнем отделе внутреннего плексиформного слоя оборван. Также полностью асимметричными являются клетки на рис. 28, отростки которых направлены в противоположные стороны.
Сетчатка лягушки. Ramon у Cajal [1972] дал прекрасные зарисовки, очень полно представляющие все многообразие амакриновых клеток сетчатки лягушки. По его тексту и особенно рисункам можно увидеть не менее 13 различных их разновидностей. Это диффузные и слоистые — одно и многослойные клетки.
Импрегнированные нами сетчатки Rana temporaria и Rana ridibunda предоставили возможность довольно детально рассмотреть ганглиозные клетки и значительно менее полно биполярные и амакриновые нейроны. Из 35 импрегнированных по Гольджи амакриновых клеток лягушки 13 были отнесены нами к слоистым, 16 — к диффузным; тип остальных был либо переходным от слоистых к диффузным, либо его было трудно определить в связи с неполной импрегнацией.
Величина тела у амакриновых клеток неравномерна, размеры наименьшей клетки — 7X7 мкм, средние размеры — 16х10 мкм, наибольшая клетка — 31 мкм (рис. 29, 30).
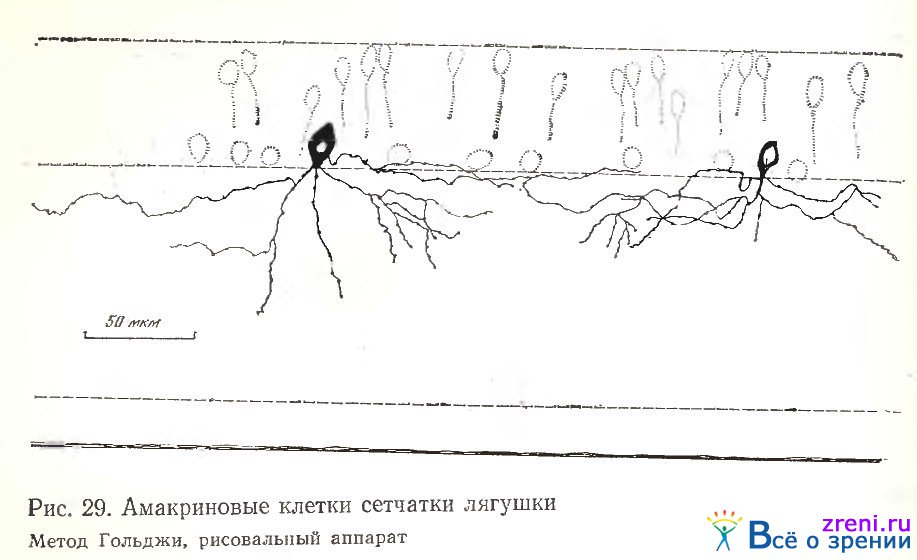

Наибольший размах ветвлений отростков у клеток на наших препаратах достигал 390 мкм, наименьший — 30. По данным Ramon у Cajal и нашим наблюдениям, этот размах может достигать 1 мм. Слоистые клетки полно представлены на рисунках Ramon у Cajal [1972]. Наши зарисовки отражают детали, не отмеченные им.. На рис. 29 однослойный нейрон, ветвящийся в наружном отделе внутреннего плексиформного слоя, виден на косо-тангенциальном срезе. Его редкие тонкие отростки извилисты, они простираются довольно далеко в горизонтальном направлении, по их ходу примерно па одинаковом расстоянии друг от друга видны небольшие округлые пуговки (см. рис. 29, слева). Из слоистых нейронов удачно (хотя недостаточно полно) импрегнировалась многослойная клетка (см. рис. 30, слева), с ветвлениями в самом верхнем и самом нижнем отделах внутреннего плексиформного слоя и в центре слоя, на трех его уровнях.
У всех трех рядом расположенных амакриновых клеток отростки переплелись (рис. 30, А) настолько, что трудно с достоверностью определить тип двух других клеток. Захождение самых наружных отростков во внутренний ядерный слой можно видеть на рис. 30, Б; видна также различная форма и величина тел у тесно рядом находящихся клеток и перекрытие отростков разных типов нейро нов. У двух клеток из девяти импрегнированных отмечается асимметрия в направлении отростков.
Диффузные амакриновые клетки сетчатки лягушки четко подразделяются на густо- и редковетвистые, узко- и широкоразмашистые. Наибольший размах на наших препаратах наблюдался у редковетвистой клетки (до 220 мкм); некоторые густо-ветвистые нейроны были весьма компактными, и изящный кустик их отростков занимал лишь 43 мкм. У диффузной густо-ветвистой клетки на рис. 30, В верхняя часть тела переходит в небольшое образование, как бы колпачок. Нижняя поверхность тела продолжается в крупный, но короткий отросток, рассыпающийся затем на множество тонких, слегка извилистых вторичных и третичных ветвлений. Все отростки, кроме основного, усыпаны синаптическими пуговками, часто располагающимися на равных расстояниях друг от друга. Этот нейрон производит впечатление как бы незавершенного, возможно, что какие-то из его нижних отростков недоимпрегнированы, хотя у большинства на концах видны терминальные пуговки.
В сетчатке черепахи амакриновые клетки весьма разнообразны. По данным Kolb [1982], их 27 типов; дендритные поля бывают узкие (30—150 мкм), малые (150—300 мкм), средние (300—500 мкм), широкие — более 500 мкм (до 2 мм). Величина тела амакриновой клетки не коррелирует с широтой размаха отростков. Обнаружена определенная корреляция характера и уровня ветвления отростков амакриновых клеток в сетчатке черепахи с их активностью. Так, клетки с отростками, ветвящимися в середине внутреннего плексиформного слоя, дают физические реакции на освещение, если же отростки ветвятся диффузно по всему внутреннему плексиформиому слою, реакции тонические [Wciler, Marchiafava, 1981].
Амакриновые клетки сетчатки млекопитающих. Амакриновые клетки ежа, так же как у других позвоночных и млекопитающих, могут быть подразделены на два основных типа: слоистые и диффузные. В наших исследованиях, несмотря на малое количество импрегнированных клеток, удалось видеть примеры обоих типов.

Однослойная клетка (рис. 31, Б, слева) характеризуется небольшим телом, от которого спускается тонкий короткий основной отросток, скоро делящийся на две веточки. По ходу более полно импрегнированного отростка расположены небольшие овальные утолщения и варикозности. Отростки этого нейрона перекрыты отростками соседнего, более крупного нейрона, вероятнее всего, относящегося к многослойным амакриловым клеткам. У диффузной клетки (рис. 31, А) миниатюрное тело плавно переходит в крупный нисходящий отросток, от которого во все стороны спускаются вторичные веточки, образующие в совокупности картину, сходную с кринолином. Площадь, занимаемая отростками нейрона, невелика, но по глубине охватывает весь внутренний плексиформный слой.
Весьма многообразны амакриновые клетки сетчатки кролика. Они наблюдаются также двух типов: слоистые и диффузные. Из слоистых чаще всего встречаются однослойные с ветвлениями либо в самом нижнем отделе внутреннего плексиформного слоя, либо в среднем его отделе. В одном случае подобный нейрон обладал очень длинными, расходящимися в разные стороны извилистыми отростками, размахом более 1 мм. Как следует из работ Famiglietti [1983а], тело обнаруженной им однослойной амакриновой клетки сетчатки кролика, названной звездорасходящейся, может находиться либо, как обычно, в слое амакриновых клеток, либо в слое ганглиозных клеток (перемещенные амакриновые клетки). В первом случае отростки расположены в наружном подслое а внутреннего плексиформного слоя и клетка соответствует физиологическому типу on-. Во втором случае отростки расположены во внутреннем подслое b и клетка соответствует типу off-.
Диффузные амакриновые клетки сетчатки кролика на наших препаратах (рис. 32, 33) имеют либо очень узкое ветвление отростков (рис. 32, слева и справа), либо средних размеров (рис. 32, в центре; 33).
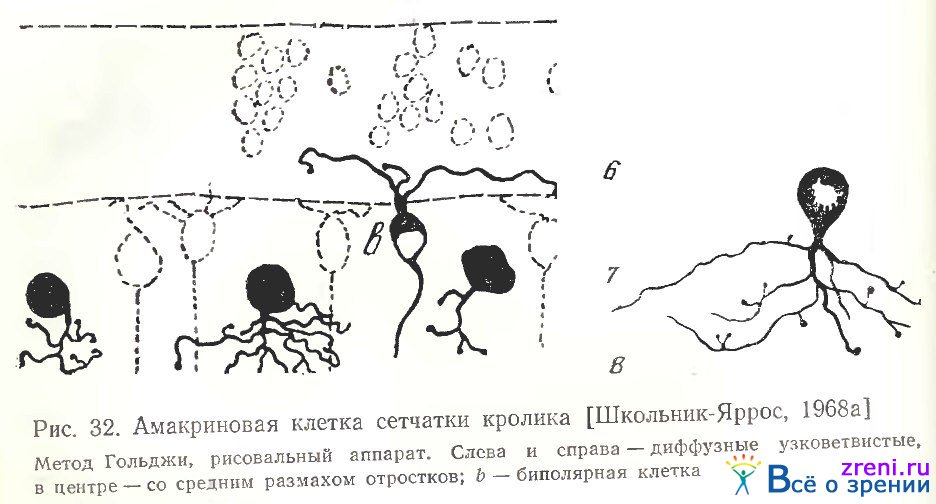
У части клеток отростки расходятся в виде небольшой редковетвистой грозди по внутреннему плексиформному слою (рис. 33). Отростки нейронов снабжены типичными шипообразными терминалями с головками-пуговками на концах. В некоторых случаях коротенькие выросты отходят непосредственно от тела клетки (рис. 32, в центре), а самые нижние ответвления расположены над слоем 8.
В сетчатке суслика обнаружено пять типов амакриновых клеток [West, 1976]. Чаще всего в материале West импрегнировались однослойные широковетвистые клетки (размах отростков более 1 мм) и узковетвистые диффузные клетки (размах — 30 мкм). Автор определил, что у диффузной клетки наибольшее количество амакриново-амакриновых синапсов 96%, тогда как у однослойных их лишь 43%. Остальные три типа амакриновых клеток, описанные West,— двухслойные узковетвистые, диффузные — стратифицированные с ветвлением во внутреннем отделе внутреннего плексиформного слоя, однослойные, ветвящиеся в подслое 1.
Сетчатка белки цитоархитектонически очень сходна с сетчаткой суслика. Однако амакриновые клетки, импрегнировавшиеся на наших препаратах, отличаются от типов, найденных в сетчатке суслика. Вероятно, это можно отнести за счет известной капризности методики Гольджи, так что в дальнейшем следует ожидать получения более полной картины нейронов.
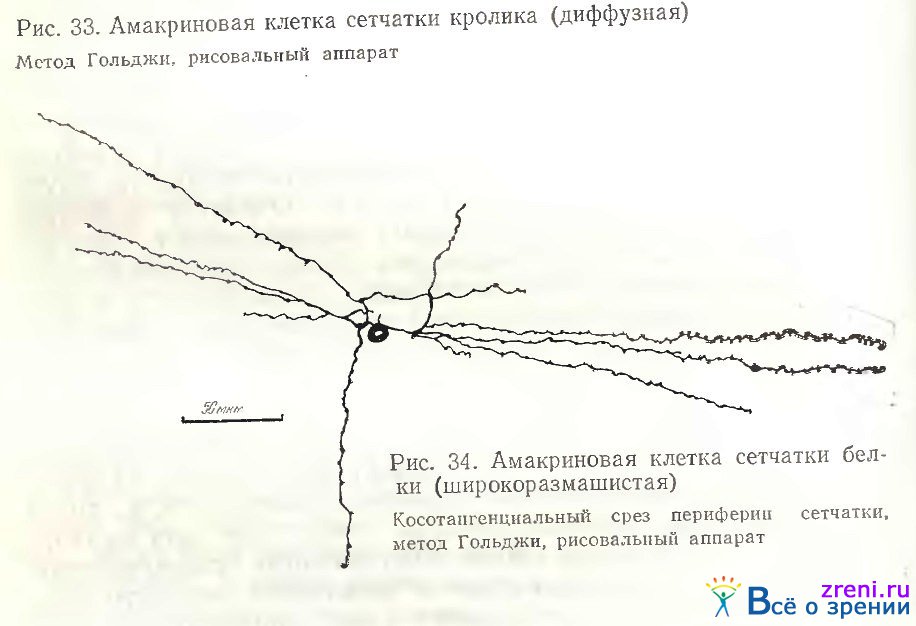
Примеры слоистых амакриновых клеток в сетчатке белки представлены на рис. 34, 35.
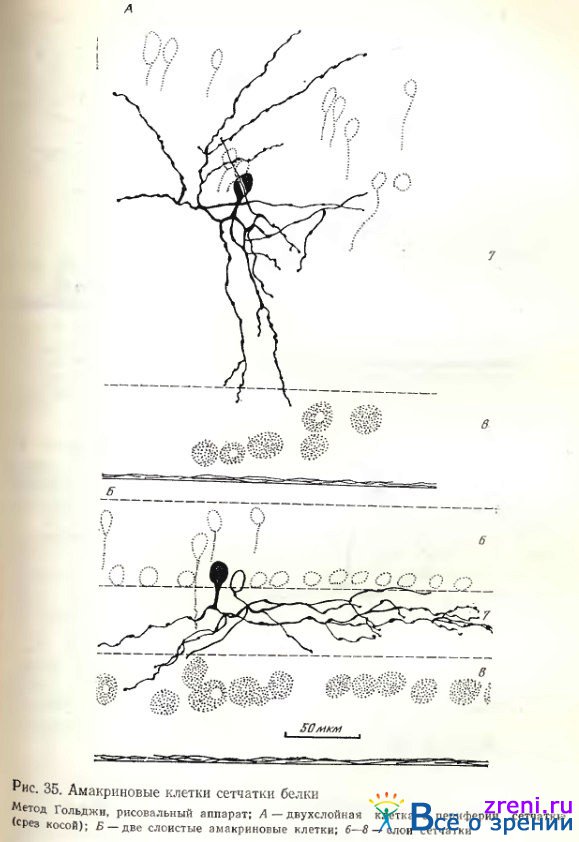
Широковетвистая клетка на рис. 34 характеризуется редкими, длинными отростками, миниатюрным округлым телом. Слегка извилистые тонкие отростки покрыты небольшими варикозностями, часто расположенными на одинаковых расстояниях друг от друга.
Признаком двухслойности на тангенциальных срезах является перекрещивание отростков, поэтому, вероятно, представленная клетка может быть причислена к двухслойным. То же относится к нейрону на рис. 35, А, отростки у которого несколько грубее, короче и компактнее расположены. Ветвления различных разновидностей диффузных клеток распространяются либо почти по всему поперечнику внутреннего плексиформного слоя (рис. 36, А), либо преимущественно в среднем его отделе (рис. 37, справа).
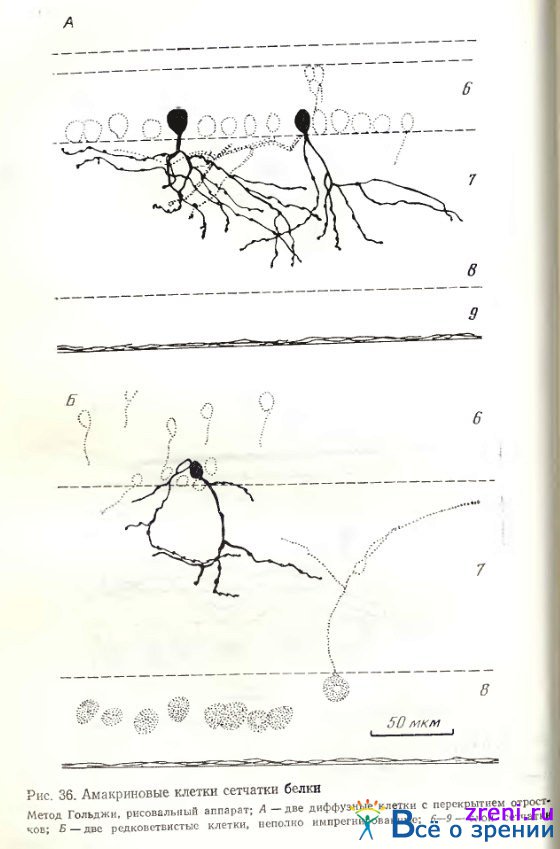
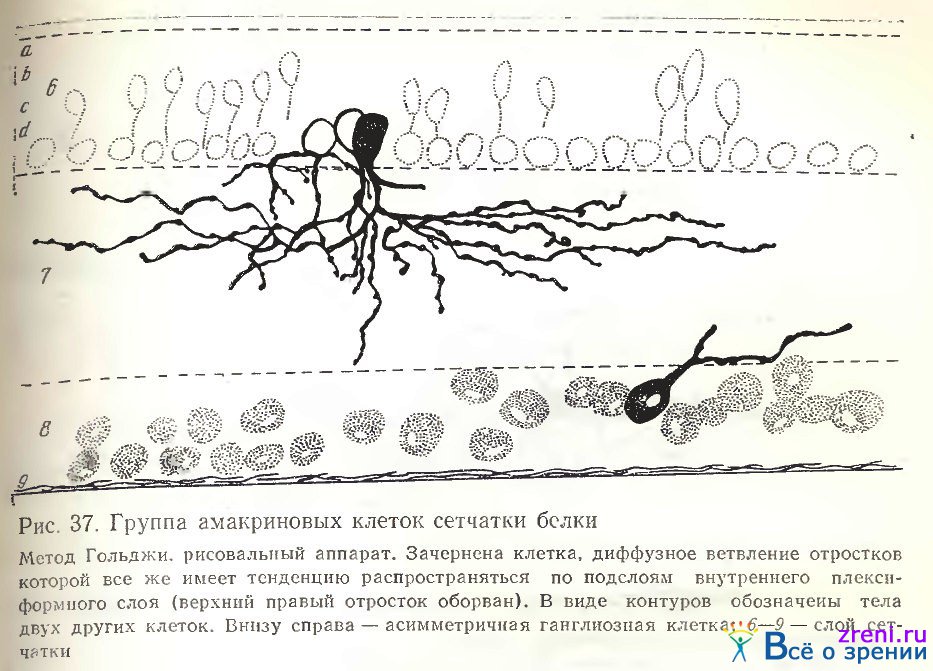
По ходу извилистых отростков и на их концах располагаются синаптические пуговки и единичные шипики. Зарисованные нами группы из трех (рис. 37) и двух (рис. 36, А, Б, 35, Б) амакриновых клеток позволяют видеть переплетение отростков различных их разновидностей, разную форму, величину тела и размах отростков.
Амакриновые клетки сетчатки кошки также делятся на диффузные и слоистые. В соответствии с шириной размаха ветвлений отростков они широковствисты, узковетвисты, либо промежуточны по величине между двумя крайними группами (рис. 38, 39).
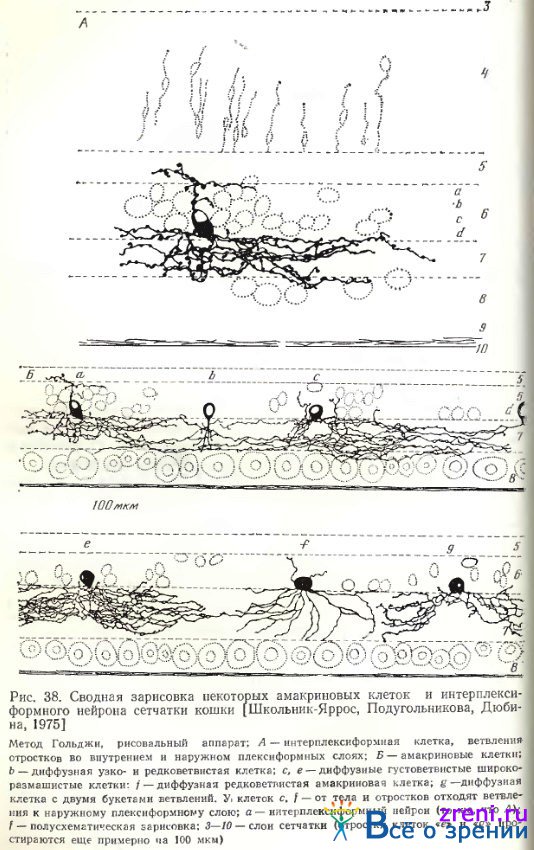
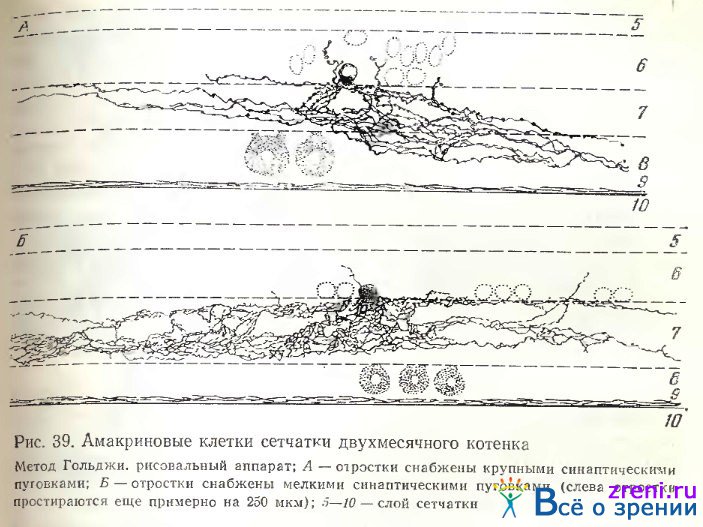
Среди слоистых наблюдаются однослойные (рис. 38, d); по материалам Kolb, Nelson [1980]; Kolb, Nelson, Mariani [1981], однослойные клетки располагаются по всем подслоям внутреннего плексиформного слоя. Авторы отметили также нейроны с более обширным ветвлением (по вертикали на радиальных срезах), занимающим один из двух выделенных ими основных подслоев — а или Ь. Всего эти авторы различают 22 типа амакриновых клеток (при импрегнации по Гольджи).
В нашей работе [Shkolnik-Yarros, 1971] диффузные амакриновые клетки импрегнировались чаще слоистых (см. рис. 38, 39); можно выделить диффузные узковетвистые и широковетвистые с разновидностями густо- и редковетвистых. У диффузных узковетвистых нейронов ветвления занимают весь поперечник внутреннего плексиформного слоя, но размах их по горизонтали но превышает 30 мкм (рис. 38, b). Чрезвычайно сходные клетки наблюдаются в сетчатке кролика (см. рис. 32, 33).
Широковетвистые густые амакриновые клетки сетчатки кошки (см. рис. 10, 38, 39) отличаются множеством весьма тесно расположенных отростков, образующих как бы сплошное кружево, занимающих весь поперечник внутреннего плексиформного слоя, размах которых может превышать 600 мкм. Отростки усыпаны пуговками по ходу. У диффузных рсдковетвистых нейронов отростки расположены значительно реже, образуя крупные петли (рис. 38, f). Густоветвистая клетка импрегнировалась также в сетчатке 15- дневного котенка (рис. 11, am); можно видеть, что отростки занимают всю ширину внутреннего плексиформного слоя, но длина их по горизонтали еще незначительна. Часто можно наблюдать отхождение небольших терминалей кверху от основной массы, в глубь внутреннего ядерного слоя, иногда они оканчиваются типичными пуговками у тел других нейронов внутреннего ядерного слоя (рис. 11, am), иногда же распространяются по направлению к наружному плексиформному слою (см. рис. 38). В некоторых случаях отростки отходят кверху прямо от тела амакриновой клетки (см. рис. 38, 39, Б).
Очень крупную клетку, расположенную целиком во внутреннем плексиформном слое, ранее обозначенную как гигантская ганглиозная [Shkolnik-Yarros, 1971), вероятно, следует рассматривать как амакриновую интерстициальную (или интраплексиформпую). Это предположение подтверждается направлением отростков не вверх или горизонтально, что характерно для ганглиозных клеток, а вниз, что закономерно для амакриновых клеток.
Электронная микроскопия и реконструкция сериальных срезов позволила обнаружить специфичность амакриновых клеток: по структуре синаптических связей они относятся к одной из систем — палочковой, колбочковой, либо к системе, ответственной за реакцию центра рецептивного поля на включение или выключение [Kolb, Nelson, 1980]. К палочковой системе относится по меньшей мере пять различных морфологических типов амакриновых клеток, каждая из которых отличается особой реакцией за освещение [Kolb, Nelson, 1983]. У широковетвящейся амакриновой клетки (А 17), сходной с импрегнированными нами (см. рис. 38, 39), изучены с помощью метки пероксидазой хрена и последующей электронной микроскопии синаптические связи пуговок, покрывающих отростки. Каждая пуговка образует реципрокный синапс у синаптической ленты аксонной терминали палочкового биполяра; в совокупности количество таких реципрокных синапсов может достигать 1000, если считать пуговки на импрегнированной клетке.
В сетчатке кошки амакриновые клетки — это интернейроны между палочковыми биполярами и ганглиозными клетками. Возможно, что широкие амакриновые клетки палочковой системы интегрируют палочковую информацию с больших площадей, увеличивая уровень чувствительности во внутреннем плексиформном слое, а узкие амакриновые клетки концентрируют и ускоряют палочковые сигналы для избирательных выходов к разным типам ганглиозных клеток [Kolb, Nelson, 1983].
По цитологическим признакам амакриновые клетки сетчатки кошки разделяются на два основных типа — тип I с относительно светлой цитоплазмой и тип II с более плотной цитоплазмой [Famiglietti, Kolb, 1975]. Методом реконструкции с сериальных электронно-микроскопических снимков авторы проследили синаптические соединения клетки типа II (рис. 40).

Удачно проимпрегнированная узковетвистая двухслойная амакриновая клетка оказалась связанной преимущественно с терминальным ветвлением палочкового биполяра.
У интерплексиформных клеток тело расположено в слое амакриновых клеток, а отростки ветвятся не только во внутреннем плексиформиом слое, как у обычных амакриновых клеток, но и в наружном плексиформиом слое (рис. 38, А). Подобные нейроны с ветвлениями в обоих плексиформных слоях наблюдались Ramon у Cajal [1972] в сетчатке собаки, но редко. Множество этих клеток видел Polyak [1941] в сетчатке приматов, он относил их к центробежным биполярам. Gallego [1971b] обратил внимание на нейроны с ветвлениями в обоих плексиформных слоях в сетчатке кошки и предложил назвать их интерплекси- формными. Затем наличие таких клеток было подтверждено в сетчатке кошки Е. Г. Школьник-Яррос, Т. А. Подугольниковой,А.П. Дюбиной [1975] и проведено детальное изучение их с помощью электронной микроскопии и гистохимических методов [Dowling, Ehinger, 1975; Boycott et al., 1975].
Из работы Dowling, Ehinger [1978] следует, что отростки аминосодсржащих иитерплексиформных клеток образуют постсинаптические соединения с амакриновыми клетками во внутреннем плексиформном слое и являются лишь иресипаптическими в наружном плексиформном слое. Такие нейроны обнаружены в сетчатке золотой рыбки (рис. 41) и у обезьяны Cebus. Довольно часто наблюдающиеся в сетчатке кошки интерплексиформные клетки [Gallego, 1971b; Boycott et al., 1975; Школышк-Яррос, Подугольникова, Дюбина, 1975] не содержат катехоламины [Boycott et al., 1975]. Однако клетки, сходные с амакриновыми, часто богаты катехоламинами в сетчатках рыб [Ehinger, Falk, Laties, 1969], птиц [Ehinger, 1967], млекопитающих [Ehinger, Falk, 1969].
Таким образом, интерплексиформные клетки, по-видимому, бывают двух разновидностей [Boycott et al., 1975]. Содержащие же катехоламины нейроны не обязательно являются интерплексиформными [Ehinger, Falk, Laties, 1969; Dowling, Ehinger, 1978]. Предполагается, что содержащие дофамин интерплексиформные клетки регулируют латеральное взаимодействие, зависящее от деятельности L-типа горизонтальных клеток в наружном плексиформном слое и фазических амакриновых клеток во внутреннем плексиформиом слое [Hedden, Dowling, 1978]. Авторы думают, что интерплексиформные клетки влияют на биполярные клетки, изменяя их реакцию с периферии рецептивного поля.
Как упоминалось выше, в сетчатке кошки нами [Shkolnik-Yarros, 1971] наблюдались также клетки с отростками, целиком расположенными во внутреннем плексиформном слое (рис. 42).

Так как и тело находится там же, справедливо отнести этот тип нейрона к интраплексиформным клеткам. По конфигурации и расположению все отростки сходны, т. е. нет четкого деления на дендриты и аксоны при световой микроскопии. Возможно, что представленный на рис. 42 интраплексиформный нейрон является смещенной амакриновой клеткой. Смещенные во внутренний плексиформный слой и в слой ганглиозных клеток ама- крнновые клетки наблюдались также Kolb, Nelson, Mariani [1981].
Амакриновые клетки сетчатки приматов тщательно изучены [Dowling, Boycott, 1965, 1966; Boycott, Dowling, 1969]. Эти авторы приняли классификацию Ramon у Cajal [1972] как более объективно отражающую действительные соотношения в морфологии амакриновых нейронов, чем классификация Polyak [1941]. boycott, Dowling [1969] выделили однослойные, двухслойные, многослойные, диффузные узковетвистые и диффузные широковетвистые амакриновые клетки (рис. 43).

Дополнительно введено новое обозначение — диффузно стратифицированные клетки, что соответствует кустовидным нейронам по нашему обозначению. Чрезвычайно убедительно проиллюстрированы однослойные, двухслойные и диффузные различных разновидностей клетки. Многослойная амакриновая клетка в сетчатке приматов отображена лишь в одном рисунке и легко может быть принята за диффузную широковетвистую.
Сделанное нами выше заключение о биполярных клетках действительно и для амакриновых. Чем шире в сетчатке данного вида внутренний плексиформный слой и чем больше в нем подслоев, тем больше должно быть разновидностей амакриновых нейронов. Так, несмотря на то что и в сетчатке кошки найдено много их разновидностей [Kolb, Nelson, 1980; Kolb, Nelson, Mariani, 1981], узость и слабая дифференцированность внутреннего плексиформного слоя обусловливают меньшее их разнообразие по сравнению с сетчаткой лягушки, щуки и изученной Т. А. Подуголышковой [1981, 1985] ставриды.
----
Статья из книги: Нейроны сетчатки | Е.Г. Школьник-Яррос, А.В. Калинина

Новость отредактировал: Dr_Michael - 30-01-2012, 20:36
Причина: Отредактировал новость, вставил фото
Комментариев 0