Развитие роговой оболочки и склеры


Описание
Особенностям развития роговой оболочки посвящено большое количество исследований. В последние годы основное их число направлено на изучение генетических и гуморальных механизмов, контролирующих этот процесс. Задачей настоящего раздела является описание основных закономерностей развития этой структуры (рис. 5.2.1 — 5.2.5).
Рис. 5.2.1. Схема развития роговой оболочки (центральные участки): а — 39 дней развития. Двухслойный эпителий располагается на базальной мембране и отделен от трехслойного эндотелия узким межклеточным пространством; б — 7 недель развития. Мезенхима — будущая строма роговицы - перемещается в пространство между" передним и задним эпителием; в — 7,5 недели развития. Мезенхима (фибробласты) формируют 5 слоев. Появляются коллагеновые волокна; г — к 3-му месяцу развития передний эпителий состоит из 2 3 слоев клеток и приблизительно 25 30 слоев стромальных фибробластов (кератобластов). Появляются признаки начала формирования десцеметовой оболочки; д — 4,5 месяца развития. Отмечается дифференциация клеток переднего эпителия по слоям (появление крыловидных клеток). Четко видна базальная мембрана и боуменова оболочка. Строма в передних слоях еще нечетко организована; е— 7 месяцев развития. Строение роговой оболочки практически полностью соответствует строению роговицы взрослого. Отличием является лишь некоторая нечеткая организация передних слоев стромы
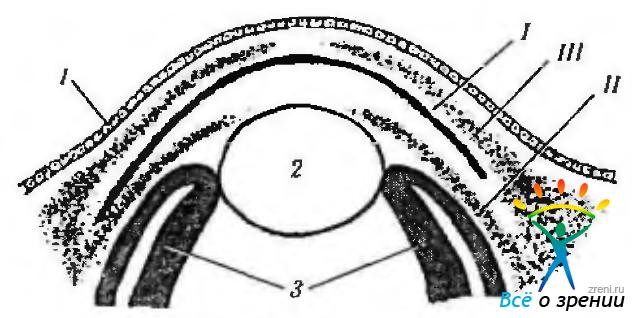
Рис. 5.2.2. Последовательность миграции клеток эктомезенхимы в передний отдел глазного яблока: I—первая волна миграции, приводящая к формированию эндотелия роговой оболочки; II— вторая волна миграции с образованием радужной оболочки и части зрачковой мембраны; III — третья волна миграции, завершающаяся образованием кератоцитов стромы роговой оболочки (1—передний эпителий роговой оболочки; 2 — хрусталик; 3 — сетчатая оболочка)

Рис. 5.2.3. Эмбриональное развитие роговой оболочки (световая микроскопия): а — строение роговой оболочки на 45 сутки (18 мм); б — строение роговой оболочки спустя 2 месяца (31 мм) эмбрионального развития: в — строение роговой оболочки спустя 5 месяцев эмбрионального развития (148 мм); г — строение передних слоев роговой оболочки новорожденного (51 см) (1 — передний эпителий: 2 — боуменова оболочка; 3— строма; 4—задний эпителий (эндотелий); 5 — хрусталик)
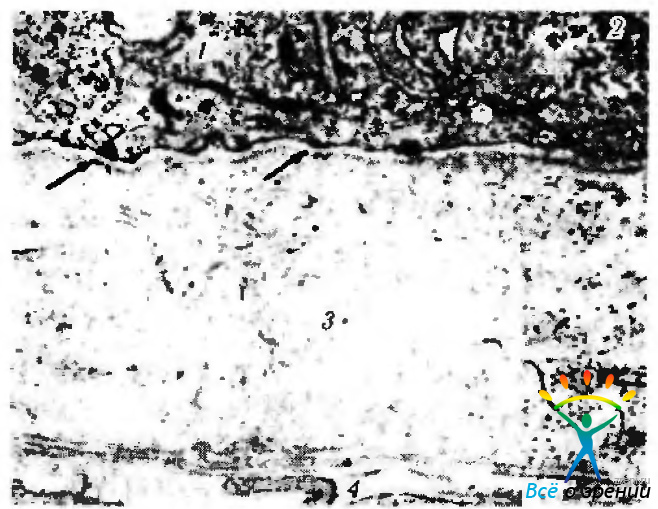
Рис. 5.2.4. Ультраструктурные особенности боуменовой оболочки на 7-м месяце эмбрионального развития. Четко определяется базальная мембрана эпителиоцитов переднего эпителия (тонкие стрелки) и полудесмосомы (широкая стрелка): 1 — эпителиальная клетка; 2— гранулы гликогена; 3— боуменова оболочка; 4 — строма роговой оболочки
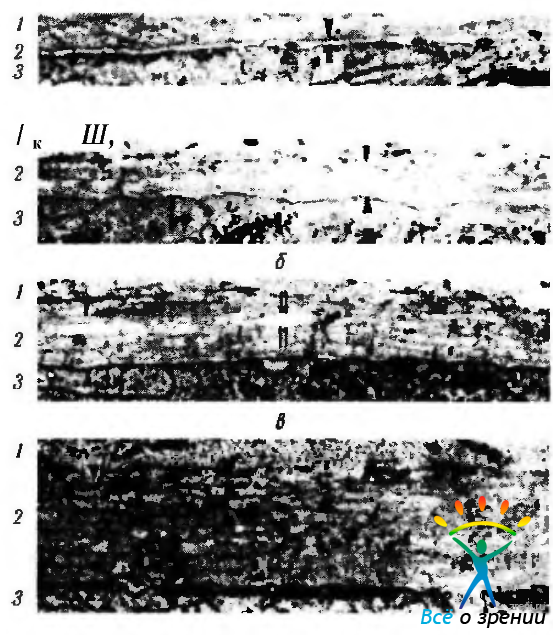
Рис. 5.2.5. Развитие десцеметовой оболочки (электронная микроскопия). Стрелками указана динамика изменения толщины десцеметовой оболочки. При этом увеличивается объем волокнистого материала оболочки: а — 3-я неделя эмбрионального развития; б - 12-я неделя развития; в — 6-й месяц эмбриогенеза; г — момент рождения (1 — строма роговой оболочки; 2 — десцеметова оболочка; 3 — эндотелий роговицы)
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Развитие роговой оболочки представляет собой сложный морфогенетический процесс. Возникает она в наружной эктодерме под индуктивным воздействием зрительного бокала. Экспериментально показано, что площадь формирующейся роговой оболочки зависит от площади ее контакта со зрительным бокалом.
В морфологическом отношении эмбриогенез роговой оболочки сводится к следующему. На пятой неделе эмбрионального развития хрусталиковый пузырек отделяется от поверхностной эктодермы и вдавливает наружную стенку зрительного пузырька. Образуется зрительный бокал. На определенном этапе погружения хрусталикового пузырька слой поверхностной эктодермы над ним смыкается и образуется зачаток роговицы. При этом роговица представляет собой довольно толстый слой эпителиальных клеток наружной эктодермы, расположенных на базальной мембране (рис. 5.2.1—5.2.4). Базальные мембраны хрусталика и эпителия роговой оболочки на этом этапе развития временно «сплавлены». Несколько позже они разъединятся.
Между нейроэктодермальными краями зрительной чаши и хрусталиковым пузырьком начинается накопление недифференцированных клеток нейрального гребня (эктомезенхима), располагающихся спереди и по периферии хрусталика, а также по краям зрительной чаши. Определяется три волны миграции этих клеток (рис. 5.2.2). Первая волна приводит к накоплению клеток, которые в последующем дифференцируются в эндотелий роговой оболочки. Вторая волна обеспечивает закладку будущей стромы. Третья волна поставляет материал для формирования зрачковой мембраны и стромы радужной оболочки.
Изучение эмбриона цыпленка показало, что отделение хрусталикового пузырька стимулирует эпителиальные клетки базального слоя секретировать коллагеновые волокна и гликозаминогликаны, выполняющие пространство между хрусталиком и эпителием роговицы. Это пространство составляет первичную строму роговицы. Первичная строма состоит из нежных микрофиламентов, аморфного материала и небольшого числа коллагеновых волокон. Из клеточных элементов она содержит только макрофаги.
Мезенхимные клетки, исходящие из нейрального гребня, расположены по краям зрительного бокала. Затем они мигрируют глубже базальной пластинки и формируют эндотелий роговицы. Эти клетки используют заднюю поверхность первичной стромы будущей роговой оболочки и базальную пластинку клеток хрусталика как субстрат для своего перемещения. Это передвижение клеток нервного гребня представляет первую волну из трех последовательных волн врастания клеток нейрального гребня в переднюю часть глаза.
Развитие эпителия. Приблизительно к 40-му дню эмбрионального развития (эмбрион 17—18 мм) роговица состоит из поверхностного слоя плоских клеток, базального слоя кубовидных клеток, первичной стромы и двух слоев плоских эндотелиальных клеток, расположенных на ее задней поверхности (рис. 5.2.1—5.2.3).
К седьмой неделе беременности эпителиальные клетки поверхностного слоя имеют гексагональную форму. Их максимальный диаметр равняется 10 мкм. На апикальной поверхности клеток уже видны микроворсинки и складки. Клетки обладают довольно электронноплотной цитоплазмой, содержащей многочисленные гранулы гликогена. Поверхностно расположенные клетки соединяются между собой и клетками, лежащими глубже, при помощи единичных десмосом.
Между вторым и третьим месяцами эмбрионального развития большинство поверхностных клеток увеличивается в диаметре и достигает 15—20 мкм. Вся их апикальная поверхность уже покрыта многочисленными микроворсинками. Цитоплазма клеток несколько просветлевает, а количество зерен гликогена уменьшается. Увеличивается и количество десмосом.
В начале четвертого месяца различается три типа поверхностных клеток. Это самые маленькие клетки, отличающиеся наличием большого количества микроворсинок и микроскладок, клетки среднего размера с промежуточным количеством микроворсинок и крупные клетки. Крупные клетки отличаются наименьшей электронноплотностью цитоплазмы и находятся в состоянии десквамации, т. е. слущивания.
К 5—6-му месяцу эмбрионального развития передний эпителий роговицы приближается по строению к эпителию взрослых людей.
Развитие стромы. Развитию стромы роговой оболочки предшествует вторая волна миграции мезенхимных клеток нервного гребня в область «губы» зрительного бокала (рис. 5.2.2, 5.2.4). Эта миграция происходит в двух направлениях. На стадии развития эмбриона, достигшего 19 мм, клетки растут в пространство между эпителием хрусталика и эндотелием роговой оболочки и формируют первичную зрачковую мембрану. Примерно в это время основное вещество стромы роговой оболочки оттекает в результате гидратации гиалуроновой кислоты. Это способствует наступлению следующей волны миграции клеток нейрального гребня. На 7-й неделе эмбрионального развития (эмбрион 22—24 мм) мезенхимные клетки мигрируют и распределяются между эпителием роговой оболочки и эндотелием.
Центральная область стромы первоначально не содержит клеток. Врастающие клетки дифференцируются в направлении стромальных фибробластов или кератоцитов, активно секретирующих коллагеновые волокна 1-го типа и основное вещество стромы.
Первоначально строма богата фибронектином. Количество фибронектина постепенно уменьшается по мере увеличения числа мезенхимных клеток. Строма вскоре достигает своей максимальной толщины, которая приблизительно вдвое больше толщины постэмбриональной роговицы. Это происходит в результате уменьшения ее гидратации, уменьшения содержания гиалуроновой кислоты, а также ретракции (сокращения) коллагеновых волокон. Звездчатые мезенхимные клетки, беспорядочно рассеянные в строме, постепенно приобретают веретеновидную форму и ориентируются параллельно поверхности роговой оболочки. Это морфогенетическая трансформация начинается в задних слоях роговицы и сопровождается быстрым появлением коллагеновых волокон. Перед появлением волокон отмечается накопление гликозаминогликанов, богатых карбоксильными и сульфатными остатками. Кератансульфат не обнаруживается до 6-го месяца беременности. В последующем коллагеновые волокна образуют пластинки. Пластинки удлиняются и утолщаются в результате постепенного накопления коллагеновых волокон (внутритканевой рост).
К 8-й неделе развития (30 мм) строма роговой оболочки в центральных участках состоит из пяти—восьми рядов клеток. Периферические участки стромы сливаются с мезенхимной тканью, которая в последующем превращается в склеру. Количество слоев стромы быстро увеличивается и на стадии развития, соответствующей 35 мм длины эмбриона, состоит из 15 слоев клеток и небольшого количества коллагеновых волокон. Задняя поверхность стромы покрыта эндотелием, состоящим из двух слоев клеток.
Как было указано выше, коллагеновые волокна растут в длину, в связи с чем увеличивается диаметр роговицы. Диаметр роговой оболочки эмбриона человека на 12-й неделе развития равен 2 мм, на 15-й неделе — 3,5 мм, на 17-й неделе — 4,5 мм, на 21-й неделе — 5,5 мм и на 35-й неделе — 9,3 мм. Как диаметр роговой оболочки, так и ее толщина определяется постоянным взаимодействием роговой оболочки с формирующейся сетчаткой и хрусталиком.
Развитие десцеметовой мембраны и эндотелия (рис. 5.2.5). К третьему месяцу развития (эмбрион 63 мм) эндотелий в центральной области роговицы становится однослойным и плоским. Располагается он на прерывистой базальной пластинке, будущей десцеметовой мембране. На этой стадии развития базальная пластинка состоит из электроннопрозрачной зоны (lamina lucida, толщина — 37,5 нм), смежной с эндотелиальными клетками, и электронноплотной зоны (lamina densa, толщина — 36,7 нм), прилежащей к строме роговицы. Дальнейшая дифференциация и утолщение десцеметовой мембраны происходят благодаря секреции эндотелиальными клетками «мембраноподобных структур». Процесс протекает довольно быстро. Если на 12-й неделе развития обнаруживается всего один слой, то к шестому месяцу слоев уже десять, а на момент рождения «мембраноподобных структур» тридцать—сорок. Параллельно с накоплением стромальных пластин появляются микрофиламенты длиной 170 нм и диаметром 40 нм. Эти филаменты располагаются перпендикулярно слоям стромальных пластин и связывают стромальные пластины, прикрепляя их к мембране. Именно благодаря этим филаментам происходит уплотнение области будущей десцеметовой мембраны.
Фибриллы будущей десцеметовой мембраны формируют гексагональные структуры, волокна в которых связаны «узлами» электронноплотного материала. В результате этого образуется равносторонний треугольник, стороны которого равны приблизительно 110 нм.
К четвертому месяцу эмбрионального развития апикальные поверхности эндотелиальных клеток соединяются при помощи десмосом. Образование контактов совпадает с началом секреции ресничным телом камерной влаги.
К шестому месяцу десцеметова мембрана уже четко различается при микроскопическом исследовании. При этом строма роговицы состоит из основного вещества, коллагеновых волокон и многочисленных активных кератоцитов.
Максимальной толщины десцеметова мембрана достигает на момент рождения.
Развитие боуменовой оболочки. На 18-й неделе эмбрионального развития базальная пластинка эпителия роговицы состоит из двух слоев — относительно тонкой lamina lucida (толщина— 41,68 нм) и полностью развитой lamina densa. Причем между эпителиальными клетками и базальной пластинкой полудесмосомы и «якорные» микрофиламенты развиты лишь частично. Постепенно микрофиламенты (средняя длина — 474 нм и диаметр — 17,6 нм) возникают у базальной поверхности эпителиальных клеток, пересекают lamina densa и lucida и заканчиваются в боуменовом слое.
Формируется боуменова оболочка в результате синтетической активности фибробластов передних слоев будущей стромы роговой оболочки. Структурные компоненты оболочки (основное вещество, волокна) также способны синтезировать клетки переднего эпителия роговицы.
К 26-й неделе беременности вблизи полудесмосом уже обнаруживаются «якорные» микрофиламенты типичного строения (рис. 5.2.4).
Необходимо отметить, что роговая оболочка эмбриона полупрозрачна, поскольку она поглощает воду в значительно большем количестве, чем роговица взрослого. Степень гидратации роговицы постепенно уменьшается по мере вызревания переднего и заднего эпителия, и на момент рождения она прозрачна. Нервные волокна в строме роговицы можно обнаружить уже на 3-м месяце развития, а к 5-му месяцу нервные окончания уже довольно равномерно распределены в эпителии. Электронномикроскопические исследования выявили, что созревание синаптических образований нервных окончаний роговой оболочки также происходит на пятом месяце развития.
Развитие склеры. Источником развития склеры являются клетки мезенхимы, которые постепенно уплотняются вокруг зрительного бокала (рис. 5.1.11). Исходят эти клетки, в основном, из нейрального гребешка. У млекопитающих задняя часть склеры, вероятно, развивается из парааксиальной мезодермы, особенно в местах прикрепления наружных мышц глаза.
Развитие склеры начинается в передних отделах на 6,5 недели развития эмбриона, и процесс постепенно распространяется назад. На этой стадии определяется восемь—девять параллельно расположенных клеточных слоев (фибробласты). Позади экватора клетки располагаются более беспорядочно.
Цитоплазма клеток содержит многочисленные свободные рибосомы, небольшое количество шероховатого эндоплазматического ретикулума с хлопьевидным материалом. Обнаруживается и плохо развитый аппарат Гольджи.
Гранулы гликогена и капли липидов, являющиеся источником энергии, более многочисленны в клетках, расположенных в переднем отделе будущей склеры. В межклеточном пространстве можно обнаружить беспорядочно распределенные коллагеновые фибриллы (диаметр — 27—29 нм). Эластические волокна на ранних стадиях развития склеры не обнаруживаются.
К середине седьмой недели эмбрионального развития уже четко определяется граница между склерой и окружающими тканями. Число клеточных слоев в передних отделах достигает 15. Клетки, прилежащие к сосудистой оболочке, отличаются большей длиной и более компактным расположением. Шероховатая эндоплазматическая сеть склеральных клеток хорошо развита. Определяется аппарат Гольджи, вблизи которого видны многочисленные зерна гликогена и капельки липидов.
Количество коллагеновых волокон к этому моменту также увеличивается. При этом их диаметр увеличивается до 30—40 нм. Появляются первые отложения эластина в виде микрофибрилл диаметром 10—12 нм.
По мере накопления межклеточного вещества исчезают контакты между цитоплазматическими отростками соседних склеральных клеток.
В течение девятой недели эмбрионального развития фибробласты задней части склеры отдают длинные отростки, ориентирующиеся параллельно слою хориокапилляров сосудистой оболочки. Нарастает количество эластических волокон.
К одиннадцатой неделе (60—65 мм) существенных различий в строении внутренних и наружных слоев склеры уже не выявляется. В это время часть мезенхимных клеток распространяется между нервными волокнами зрительного нерва, они ориентируются поперечно и формируют решетчатую пластинку.
В начале четвертого месяца склера в пре- и постэкваториальных областях состоит из 30 слоев клеток, а к шестому месяцу — из 50 слоев. В дальнейшем количество слоев не увеличивается. Увеличивается лишь объем межклеточного вещества и количество коллагеновых (диаметр — 100 нм) и эластических волокон. Необходимо отметить, что, хотя клеточные элементы роговой оболочки и склеры имеют общее происхождение (мезенхима), они ведут себя по-разному в отношении синтетической активности. Herrmann установил, что интезируемый роговичными клетками коллаген отличается по своему составу от коллагена, синтезируемого клетками склеры. Более того, различно соотношение коллагеновых и неколлагеновых белков и скорость их синтеза.
Несколько позже (эмбрион 70 мм) во внутреннем слое передней части склеры развивается клиновидной формы выступ, склеральная шпора. Форма этой структуры объясняется натяжением участка склеры развивающимися волокнами ресничной мышцы. К середине седьмого месяца передние концы волокон продольной ресничной мышцы прикрепляются к склеральной шпоре. Коллагеновые и эластические волокна в области склеральной шпоры ориентированы циркулярно.
Интрасклеральные сосудистые сплетения, водяные вены и коллекторные каналы, пересекающие склеру в лимбальной области, впервые появляются на 12-й неделе эмбрионального развития. Эти сосуды дифференцируются из примитивной мезодермы.
Поскольку склера определяет размер и форму глазного яблока, факторы, регулирующие ее развитие, играют основную роль в формировании рефракционных особенностей глаза. Факторов, индуцирующих развитие склеры, как и других структур глаза, большое количество, и секретируются они многими структурами. Одним из таких факторов, принимающих особое участие в дифференциации склеры, является матричная РНК, контролирующая синтез инсулин-подобного фактора роста. В наибольшем количестве обнаруживается он в клетках нейрального гребня, откуда и направляется к развивающимся структурам. В зрелой склере, роговице и сетчатке эта РНК не выявляется. Дальнейший рост склеры после рождения контролируется другими факторами роста, которые синтезируются и высвобождаются из сетчатки. Сигнал, побуждающий к развитию склеры, относится к пептидным гормонам, синтез и выделение которых стимулируется светом, попадающим на развивающуюся сетчатку. Именно по этой причине у новорожденных, лишенных, по разным причинам, зрительных стимулов, отмечается неполное созревание склеры, что может явиться причиной развития близорукости.
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0