Двухстадийная теория цветового зрения


Описание
Альтернативной одностадийной гипотезе Юнга—Гельмгольца и Геринга стала концепция, в которой предлагалась более сложная схема цветового анализа излучений. Она предусматривала уже две последовательные стадии анализа — рецепторную и нейрональную (рис. 1.4) и объединяла в себе две одностадийные модели — Гельмгольца и Геринга.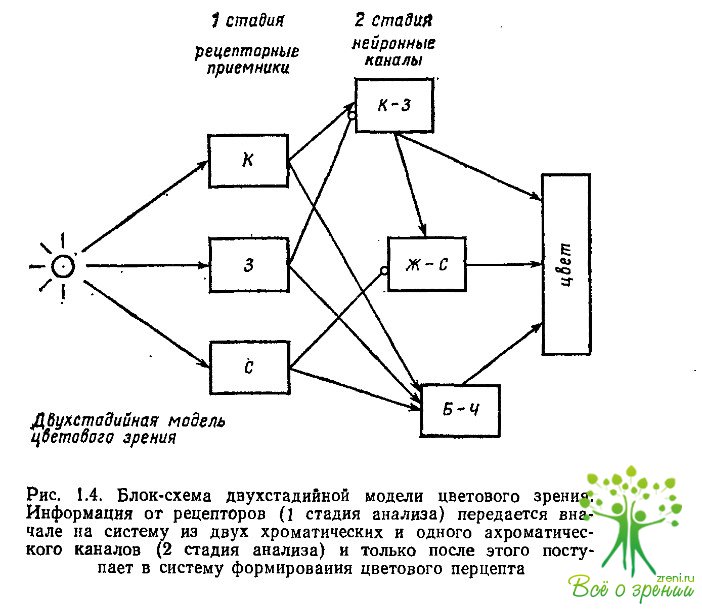
В двухстадийной модели на первом этапе анализ световых излучений осуществляется тремя рецепторними приемниками, а далее, на второй стадии, информация преобразуется нейрональной системой из двух цветооппонентных и одного яркостного механизмов. В настоящее время такая двухстадийная модель цветового зрения имеет наибольшее число приверженцев, хотя внутреннюю структуру каждой стадии разные исследователи представляют себе по-разному.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
В первой половине XX в. основное внимание обращалось на модификацию рецепторного звена. Обсуждалась очень остроумная гипотеза о возможности четырех разных рецепторных приемников, гипотеза об однородности сетчатки, согласно которой все приемники сетчатки являются одинаковыми, но сам приемник имеет сложную трехпигментную структуру; полихроматическая гипотеза Хартриджа предполагала наличие 6—8 разных приемников и т. д. Обзор и подробный анализ этих гипотез можно найти в работе Леграна.
Но во второй половине XX в. акценты смещаются, все теоретическое внимание исследователей переходит на обсуждение структуры второй, нейрональной, стадии цветового анализа излучений. Причин для этого было несколько. Самая главная причина заключалась в том, что в 50-е и особенно в 60-е гг. целый ряд исследователей в совершенно блистательных экспериментах выявили реальную структуру рецепторных приемников сетчатки, участвующих в анализе спектрального состава излучений.
Это, в первую очередь, биохимические исследования пигментов рецепторов, сетчатки, проведенные Брауном и Уолдом, электрофизиологические исследования спектральных характеристик отдельных рецепторов, проведенные с помощью микроэлектродной техники Томита с сотрудниками, микроспектрофотометрические измерения отдельных рецепторов, проведенные Брауном И УОЛДОМ! , а также Марксом, Доубелом и МакНиколем, микроденситометрические измерения характеристик отдельных рецепторов в интактной сетчатке.
Эти и последующие за ними работы выявили, что сетчатка человека и наиболее похожая на нее сетчатка приматов содержит только три типа колбочковых пигментов, которые распределены между тремя типами колбочек, так что одна колбочка содержит только один пигмент.
Другие, не менее важные причины, заключались в том, что в 50-е гг. были впервые разработаны психофизические и нейрофизиологические методы исследования цветовой оппонентности. В начале 50-х гг. американские исследователи Л. Харвич и Д. Джемсон разработали психофизический метод кансиляций для измерения цветовой оппонентности, и впервые сумели получить, экспериментально цветооппонентные характеристики зрения человека, а в конце 50-х гг. Светихин и МакНиколь зарегистрировали в сетчатке у рыбы клетки с цветооппонентными спектральными характеристиками.
Последующий за этими работами нейрофизиологический «бум» привел к открытию целого ряда цветооппонентных клеток не только в сетчатке рыбы, но и в сетчатке других животных, и в частности—приматов.
Вслед за этим цветооппонентные клетки были обнаружены в более центральных звеньях зрительной системы — наружном коленчатом теле и стриарной коре. Постепенно становилось понятным, что каждый цветооппонентный канал представляет собой не просто отдельную клетку, аналогично рецепторам сетчатки, а сложную нейрональную сеть.
Возникает вопрос — каким же образом информация, распределенная по этим сложным нейронным сетям, преобразуется в целостное и нераздельное ощущение, которое мы называем цветом?

Комментариев 0