АНАЛИЗ СПЕКТРАЛЬНОГО СОСТАВА ИЗЛУЧЕНИЙ НЕЙРОНАМИ СЕТЧАТКИ | Горизонтальные клетки


Описание
Морфология.Горизонтальные клетки располагаются в сетчатке сразу за фоторецепторами в дистальных отделах внутреннего ядерного слоя. Размеры сомы этих клеток варьируют в пределах 50 ч-200 мкм. Часть клеток имеет аксоны, которые у большинства позвоночных контактируют с рецепторами и лишь у рыб (карп, карась) оканчиваются в проксимальной области внутреннего ядерного слоя.
Горизонтальные клетки залегают в несколько слоев, число которых у разных животных варьирует от 1 до 4. Для каждого отдельного слоя характерно упорядоченное расположение клеток в виде периодических решеток, согласующихся с мозаикой рецепторов. Функциональное назначение мозаичной организации рецепторного и нервных слоев остается пока невыясненным. Между клетками одного слоя имеются электрические синапсы типа «gap junction» (щелевой контакт — тесное смыкание мембран двух соседних клеток с шириной синаптической щели 20—100 А) и типа «tight junction» (плотный контакт — когда мембраны соседних клеток сливаются). В наружном синаптическом слое есть также химические синапсы между телами и отростками неидентифицированных клеток (возможно, из разных слоев).
В морфологической организации сетчатки млекопитающих имеются особенности, отличающие ее от сетчатки низших позвоночных: горизонтальные клетки здесь менее крупны, менее разнообразны по форме сомы и ветвлению отростков и образуют меньшее число слоев.
По типу преимущественных контактов с теми или иными видами рецепторов выделяют колбочковый, палочковый и «смешанный» типы клеток.
Передача сигнала от фоторецептора к горизонтальной клетке.
В темноте рецепторы позвоночных непрерывно выделяют медиатор, деполяризующий горизонтальные клетки. Гиперполяризация фоторецептора светом приводит к уменьшению выделения медиатора и, как следствие, к гиперполяризации горизонтальной клетки. Под влиянием медиатора в темноте субсинаптическая мембрана горизонтальной клетки проницаема для ионов Na+ и С1~. Каналы субсинаптической мембраны проницаемы, по-видимому, также для ионов К+. В отсутствие света мембранный потенциал горизонтальной клетки определяется в основном каналами субсинаптической мембраны и варьирует в пределах —5 ч- -5~25 мВ.
Несинаптическая мембрана горизонтальных клеток обладает резко выраженной нелинейностью: ее сопротивление падает при гиперполяризации на свету и возрастает при деполяризации в темноте. Нелинейность определяется работой потенциал-зависимых К+ и Са2+ каналов. Проницаемость для К+ возрастает при гиперполяризации и уменьшается при деполяризации. Проводимость для Са2+ возрастает при деполяризации ниже, чем —50 40 мВ.
Таким образом, сопротивление субсинаптической и несинаптической мембран меняется в противоположные стороны при изменении освещенности. При этом общее сопротивление мембраны клетки остается постоянным, что создает одинаковые условия для распространения по слою горизонтальных клеток потенциалов разной величины. Нелинейные свойства несинаптической мембраны позволяют клетке усиливать медленные потенциалы и способствуют передаче потенциалов по длинному аксону (300-М00 мкм) от которого отводятся примерно такие же ответы, как и от сомы.
Нелинейная соматическая мембрана горизонтальных клеток является частью системы с обратными связями «рецептор — горизонтальная клетка», которая в целом может вести себя линейно, компенсируя нелинейности отдельных своих компонентов (субсистем): процессов поглощения квантов света, мембраны рецептора синаптической передачи и др. Об этом свидетельствуют данные по линейной суммации реакций колбочек на мембране горизонтальных клеток.
[banner_centerrs] {banner_centerrs} [/banner_centerrs]
Согласно этим данным, функции спектральных реакций горизонтальных клеток L-, R+G- и G+R-B- (К+В-)-типов в линейном диапазоне работы клеток и при небольших энергиях света с высокой точностью (5%) аппроксимируются линейной комбинацией «взвешенных» значений функций спектральных реакций В-, G и R-колбочек. Степень линейности реакций горизонтальных клеток зависит не только от условий стимуляции, но и от того, какая фаза ответа берется в качестве критерия при построении спектральных функций — пик или плато.
Последнее больше подвержено поздним меж- и пострецепторным взаимодействиям, связанным с адаптацией и вносящим основной вклад в нелинейность реакций. Поэтому связь между плато реакции колбочки и плато реакции горизонтальной клетки можно считать линейной лишь в диапазоне малых интенсивностей, где не функционируют потенциал-зависимые калиевые каналы несинаптической мембраны и механизм обратной связи.
Спектральные характеристики горизонтальных клеток.
На белый свет все клетки отвечают довольно медленным гиперполяри-зационным отклонением трансмембранной разности потенциалов (время достижения максимума у колбочковых клеток 50-100 мс), которая возвращается к исходному уровню темнового потенциала лишь после прекращения действия света. Такую реакцию, впервые зарегистрированную венесуэльским исследователем Светихиным в 1953 г., принято называть 5-потенциалом (от англ. «slow». — медленный). 5-потенциалы зарегистрированы в сетчатках всех позвоночных, имеющих горизонтальные клетки.
По реакциям на спектральные стимулы клетки делятся на два типа: яркостные L (от англ. «luminosity») — клетки, отвечающие гиперполяризацией на свет любой длины волны, и цветовые С (от англ. «colour») — клетки, полярность ответа которых меняется в зависимости от длины волны светового стимула (рис. 3.2.1а).

Функции спектральных реакций L-клеток (рис. 3.2.1 б) у разных животных и в сетчатке одного животного широко варьируют по форме и положению максимума. Так, в сетчатке костистых рыб обнаружены /.-клетки с пиками реакций на 460—495 нм, 525—580 нм и 600—680 нм. Часть L-клеток контактирует только с колбочками (колбочковые клетки), часть — только с палочками (палочковые L-клетки).
Антагонистическое взаимодействие палочковых и колбочковых сигналов в отдельных L-клетках обнаружено в сетчатках черепахи, кошки, саламандры, обезьяны. Одновременные контакты с фоторецепторами разного типа определяют сложную (с несколькими пиками) форму функций спектральных реакций L-клеток и их зависимость от условий световой и хроматической адаптации. Вклад колбочек разного типа » реакции клеток L-типа также варьирует.
Электрофизиологически — по функциям спектральной чувствительности, получаемым методом равных ответов, — выявлены клетки, связанные только с R-колбочками или с R- и G-колбочками, или со всеми тремя колбочками. В определенном диапазоне интенсивностей стимуляции функции спектральной чувствительности L-клеток аппроксимируются линейной комбинацией функций спектральной чувствительности соответствующих типов колбочек.
Располагаясь в один слой, клетки L-типа связаны между собой электрически в синцитиальную сеть. Об этом свидетельствуют большие размеры их рецептивных полей, а также прямые опыты с пропусканием тока через микроэлектрод в одну L-клетку и отведением потенциала через другой микроэлектрод из соседней L-клетки.
Размеры рецептивных полей L-клеток, варьирующие в диаметре от 300 мкм до 5 мм, значительно превышают протяженность их дендритных ветвлений. Принято считать, что рецептивные поля L-клеток круглые и не имеют деления на оппонентные друг к другу центре и периферию. Однако результаты отдельных работ последних лет свидетельствуют о более сложной организации рецептивного поля.
На лягушке и черепахе методами морфологии и электрофизиологии получены данные, что рецептивные поля клеток L-типа имеют форму эллипса, определенным образом ориентированного в плоскости сетчатки. Исследования методами тонкой морфологии и локальной световой стимуляции показали, что огромное по размерам рецептивное поле горизонтальной клетки обладает локальной анизотропией — неодинаковостью в морфологической организации и свойствах реакций в. разных участках. Локальная анизотропия свидетельствует о функциональной неоднородности рецептивных полей и возможности локальной обработки информации в разных его зонах.
Амплитудные функции L-клеток, так же как и амплитудные функции фоторецепторов, описываются уравнением Михаэлиса — Ментон. Однако амплитудные функции горизонтальных клеток имеют более крутой наклон: на одно и то же изменение в интенсивности цвета они реагируют большим изменением потенциала, нежели рецепторы.
Пространственная суммация спектральных сигналов в рецептивных полях L-клеток не всегда носит линейный характер и зависит от интенсивности света. О более сложном, нежели простая алгебраическая суммация, взаимодействии сигналов в рецептивных полях клетки L-типа свидетельствует и «эффект усиления».
Длинноволновый фон усиливает реакцию L-клетки на коротковолновое излучение.
Эффект усиления проявляется и при фоновом освещении нейтральным светом, возбуждающим все типы рецепторов: реакция на ахроматический тестовый стимул возрастает при условии слабого нейтрального фона. Эффект отсутствует, если тестовый и фоновый сигналы (например, длинноволновое излучение) возбуждают один тип рецепторов. Это доказывает, что суть дела именно во взаимодействии сигналов от разных рецепторов.
Форма функций спектральных реакций колбочковых L-клеток близко совпадает с формой кривой видности животного в фотопических условиях, оптимальных для различения спектральных излучений. Это, а также суммация L-клетками сигналов от колбочек всех типов на большой площади сетчатки являются основанием для отнесения их к «яркостным» каналам сетчатки. Однако форма спектральной характеристики L-клетки не полностью совпадает с кривой видности, которая определяется вкладом не только L, но и С-клеток. По современным данным, L-клетки принимают участие и в формировании спектральной оппонентности клеток С-типа через механизм обратной связи с разными колбочками.
Горизонтальные клетки С-типа меняют полярность реакций (гиперполяризация — деполяризация) в зависимости от длины волны света, и поэтому их относят к цветокодирующим клеткам. Три основные разновидности клеток С-типа описаны в сетчатке рыб и черепахи: R+G-, деполяризующиеся длинноволновым и гиперполяризующиеся коротко- и средневолновыми излучениями: G+R-В-, деполяризующиеся средневолновым и гиперполяризующиеся коротко- и длинноволновыми излучениями, и Y+В-, депо, ляризующиеся средне- и длинноволновыми и гиперполяризующиеся коротковолновым излучениями (рис. 3.2.1а) Y+B-клетки, по-видимому, не являются самостоятельным типом клеток и по ряду критериев представляют собой разновидность G+Я-В-, клеток с редуцированным длинноволновым входом.
В сетчатке высших животных, в том числе у приматов, клетки С-типа пока не обнаружены. Синаптические контакты С-клеток с рецепторами строго избирательны. По данным морфологии, R+G--клетки получают сигналы в основном от С-колбочек, G+R-В- и Y+В- — от В-колбочек.
Установлено, что каждый тип С-реакций генерируется своим морфологическим типом горизонтальных клеток, образующим отдельную синцитиальную сеть (электрический синцитий R+G--клеток, синцитий G+R-В--клеток). Рецептивные поля С-клеток принципиально сходны с рецептивными полями G-клеток: их размеры могут достигать 5-6 мм в диаметре, они не имеют пространственного деления на центр и периферию, изотропны — в каждой точке реадптивного поля спектральные свойства С-клетки примерно одинаковы. Зависимость амплитуды спектральных реакций С-клетки, так же, как и L-клетки, хорошо аппроксимируется уравнением Михаэлиса — Ментон.
Однако реакции С-клеток в большей степени, нежели реакции клеток L-типа, зависят от суммарной энергии излучения и условий световой и хроматической адаптации. Так, большие размеры или высокая интенсивность стимула могут приводить к нарушению спектральной оппонентности R+G--клеток (рис. 3.2.1), поскольку при увеличении и площади и интенсивности света происходит резкое увеличение общего потока квантов Р, достигающих сетчатки:
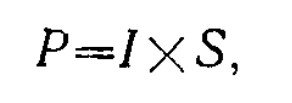
где I — число квантов, поступающих в единицу времени на единицу площади сетчатки, S — площадь засвета. При избирательном воздействии адаптирующим светом на один из реценторных входов С-клетки увеличиваются ее реакции с других входов: на средневолновом фоне увеличиваются реакции R+G--клетки на длинноволновые излучения. Все вышеперечисленное определяет сложный характер зависимости реакций клеток С-типа от размеров и интенсивности спектрального стимула, а также условий световой и хроматической адаптации.
Гиперполяризация R+G-, Y+B- и G+R-B--клеток возникает при освещении тех типов колбочек, с которыми они имеют прямые синаптические контакты. Ионные механизмы гиперполяризационных ответов С-клеток идентичны механизмам генерации гиперполяризационных реакций субсинаптической мембраной L-клеток. Относительно способов формирования реакций на цвета из других областей спектра существует две гипотезы.
Согласно первой, G-колбочки получают сигналы обратной связи с инверсией знака от L-клеток, а синие колбочки — от R+G--клеток (рис. 3.2.2 а, б). И тогда длинноволновое излучение, гиперлоляризующее R-колбочки и L-клетки, вызывает через обратную связь деполяризацию G-колбочек и связанных с ними клеток. Средневолновое излучение, гиперполяризующее R+G--клетки, вызывает через обратную связь деполяризацию В-колбочек и связанных с ними Y+В-- и G+R-В--клеток.

Длинноволновый сигнал, дважды пройдя через систему обратной связи с инверсией знака, снова становится гиперполяризующим в G+R-B--клетках (от L-клетки к G-колбочке, от нее к R+G--клетке через прямые синапсы и от R+G--клетки к B-колбочке через обратную связь, от B-колбочки к G+R-B--клетке). Эта гиперполяризация слабо выражена в G-R-B--клетках и совсем незаметна в Y+В--клетках.
Рядом исследователей разрабатывается гипотеза, согласно которой тормозные и возбуждающие медиаторы поступают на С-клетки прямо и независимо от колбочек разного типа. Баланс в притоке этих медиаторов, воздействующих на разные ионные каналы субсинаптической мембраны горизонтальных клеток, определяет нолярность ответа клетки.
При этом разница в латентпости реакций на разные цвета объясняется разницей в кинетике взаимодействия медиаторов с постсинаптической мембраной, как это известно для биполяров: в сетчатке черепахи латентный период деполяризационных реакций в центре рецептивного поля off-бииоляра заметно больше латентного периода гиперполяризационных реакций в центре рецептивного поля off-биполяра.
Существует тесная связь между наличием в сетчатке горизон-тальных клеток С-типа и способностью низших позвоночных к различению цветов. По данным Тамура и Нива, клетки С-типа обнаруживаются в сетчатках у рыб с хорошо выраженной способностью к цветоразличению (карп, золотая рыбка, кефаль) и не регистрируются в сетчатках у цветослепых рыб.
По мнению этих авторов, регистрация S-потенциалов является удобным методом изучения цветового зрения у рыб, особенно у тех, которые малодоступны для исследования поведенческими методами. Редукция системы колбочек в сетчатке рыб сопровождается редукцией слоев и типов горизонтальных клеток и, как следствие, приводит к разного рода нарушениям цветового зрения.
Так, у хищной рыбы Nannacara anomala с дихроматическим цветовым зрением имеется только один колбочковый С-тип горизонтальных клеток, возможно достаточный для генерации RG-потенциалов. У альбиносов форели потеря одного типа колбочек сопровождается потерей двух слоев горизонтальных клеток.
Таким образом, горизонтальные клетки не только обеспечивают латеральные связи между рецепторами, контролируя адаптационные изменения в сетчатке и участвуя в механизмах пространственного контраста и дирекциональной чувствительности. Результаты ряда работ указывают на их участие в формировании спектральных и пространственных свойств рецептивных полей цветокодирующих и ахроматических биполяров.
В настоящее время исследуются пути такого участия: через обратные связи с фоторецепторами или же через прямые контакты с биполярами. Имеются данные и об участии горизонтальных клеток в формировании реакций ганглиозных клеток сетчатки. В отдельных случаях реакции горизонтальных клеток на цвет могут коррелировать с реакциями на цвет ганглиозных и амакриновых клеток. На возможность участия горизонтальных клеток в генерации цветовых реакций выходных элементов сетчатки указывает и тот факт, что часть из них имеет аксоны, мембрана которых обладает способностью к бездекрементному проведению сигнала.
По данным некоторых авторов, L-клетки кодируют яркость излучения, так как форма их функций спектральных реакций близко совпадает с формой функции чувствительности глаза к спектральным стимулам.
Надо отметить, что функции спектральных реакций L-клеток по форме также близки к спектральному распределению энергии, характеризующему дневной «белый» свет в среде обитания животного.
Так, пресные воды на глубинах обитания карпа (1-4 м) имеют максимум пропускания излучения в длинноволновой области спектра (590-630 нм). В этой же части спектра локализуется максимум спектральных реакций L-клеток этих животных. Это значит, что белый свет для рыб отличается по спектральному составу от белого света для человека большим содержанием оранжевых и красных тонов.

Комментариев 0