Развитие нервной системы и глазного бокала


Описание
Среди заболеваний органа зрения особое место занимает врожденная и наследственная патология. Проявляется она возникновением пороков, по-разному влияющих на функцию органа зрения. Для правильного понимания причин возникновения пороков развития и особенностей их проявления необходимы знания об особенностях эмбрионального развития органов и тканей организма человека, в частности глаза.Многими как отечественными, так и зарубежными исследователями было установлено, что наиболее важным этапом развития является эмбриональный морфогенез, включающий эмбриональный гистогенез и органогенез. Сущность эмбрионального гистогенеза сводится к возникновению в процессе развития специализированных тканей из малодифференцированных клеток. В результате органогенеза развиваются органы и системы органов. Этот процесс происходит на всех уровнях — клеточном. тканевом, межтканевом, органном и межорганном. Нарушение любого из этих механизмов влечет за собой отклонение от нормального развития.
На протяжении всего эмбрионального развития организма одновременно происходит ряд последовательно развивающихся процессов пролиферации клеток, их перемещение в пространстве, дифференциация и гибель. Перечисленные процессы управляются многочисленными механизмами индукции и супрессии, действующими на клеточном и молекулярном уровнях.
Благодаря процессам индукции на ранних этапах развития глаза образуются группы зародышевых тканей, из которых в последующем формируются различные части глаза. Индукция представляет собой цепь процессов, в результате которых в ответ на сигнал, исходящий из соседних клеток, происходит специфическая дифференциация эмбриональных клеток. Самые первые признаки индукции обнаруживаются уже на самых ранних этапах эмбриогенеза, а именно на этапе гаструляции. На этом этапе эктодерма охватывает дорсальную поверхность эмбриона в виде одного слоя и в последующем расщепляется на два слоя. Из наружного слоя клеток образуются эпителиальные производные, а из внутреннего — нервная пластинка. Нервная пластинка является предвестником центральной нервной системы, включая область, расположенную в передней ее части, из которой и образуется зачаток глаза.
После появления нервной пластинки наступает нейруляция, процесс, при котором нервная пластинка «сворачивается» в трубку по срединной линии эмбриона. При нейруляции происходят существенные изменения клеток в различных участках нервной пластинки под воздействием индукционных стимулов соседних тканей. При этом происходит перемещение тканей нервной трубки, которые дифференцируются в различных направлениях. Примером таких изменений является образование зрительного пузырька и в последующем зрительного бокала.
Образование глазного бокала имеет большое значение в дальнейшем развитии глаза, на чем мы остановимся несколько ниже. Основным в этом процессе является дифференциация различных структур глаза с появлением в них различных функций. Это образование оболочек глаза, зрительного нерва и др. Все эти процессы также происходят благодаря процессам индукции.
Классическим примером индуктивного значения глазного бокала является развитие хрусталика. Хрусталик развивается из участка эктодермы, который входит в контакт с глазным бокалом в течение нейруляции. Контакт между этой эктодермой и глазным бокалом приводит к уплотнению части эктодермы с последующей дифференциацией в хрусталик.
Таким образом, индуктивные взаимодействия представляют собой процесс, при котором после дифференциации одного участка эмбриональной ткани именно он и стимулирует дифференциацию рядом расположенных участков.
Возникает вопрос: благодаря каким механизмам осуществляется эмбриональная индукция? Природа индуктивного сигнала изучалась на протяжении многих десятилетий. Лишь недавно была установлена их химическая природа. Выявлена целая группа веществ, индуцирующих развитие эмбриональных тканей. Относятся они к полипептидам и называются полипептидными факторами роста. Установлено, что эти вещества принимают участие во многих функциях тканей, включая процессы пролиферации клеток, а также их дифференциации. Было выделено много классов полипептидов. отличающихся молекулярной массой и последовательностью аминокислотного состава. Первоначально способность к эмбриональной индукции одного из веществ этой группы, а именно «активина», была установлена in vitro. В настоящее время известно, что молекулы, схожие с «активином», присутствуют на ранних стадиях эмбриогенеза при индукции развития многих тканей эмбриона, в частности нервной ткани и хрусталика. К таковым можно отнести так называемый цилиарный нейротрофический фактор.
Таким образом, полипептидные факторы роста являются сигналами стимулирования развития, лежащими в основе эмбриональной индукции. При этом возникает вопрос: каким образом эктодерма отвечает на сигналы индукции, формируя при этом различные части глаза? Основной догмой эмбриологии является предположение о том, что возможность образования новой дифференцированной ткани возникает в результате экспрессии специфического гена. Именно экспрессия тканеспецифичных генов приводит к формированию дифференцированной клетки, в частности тканей глаза. Например, экспрессия генов родопсина приводит к развитию специализированных клеток зрительного рецептора, также как экспрессия генов кристаллина приводит к дифференциации клеток хрусталика.
По данным Bortoluzzi, d'Alessi, Danieli, анализировавшим сведения, приведенные в Internet, в развитии только сетчатой оболочки глаза участвует порядка 3152 генов.
Регуляторные гены выявлены у многих животных. Белковые производные этих генов являются факторами транскрипции, управляющими инициированием синтеза РНК и в последующем специфических белков.
Развитие глаза на различных этапах развития эмбриона можно представить как процесс взаимодействия генов, при котором происходит последовательная экспрессия различных регуляторных генов. Эта последовательность событий приводит к образованию зачатков тканей глаза с их собственным специфическим образцом экспрессии генов. Следующий этап развития глаза сводится к дифференциации этих зачатков с образованием специфических тканей благодаря процессам морфогенеза и дифференциации различных типов клеток.
Одним из таких генов является семейство Рах генов, играющих роль не только в развитии глаза, но и других тканей и органов. У многих животных экпрессия Рах2 гена выявлена в центральной нервной системе, глазном яблоке, ухе, мезенхиме. Мутации этих генов приводят к возникновению множественных аномалий, включая поражение глаза (синдром Ваарденбурга (Waardenburg), аниридия, колобома).
В развивающемся глазном яблоке экспрессия гена Рах2 первоначально определяется в дистальных областях зрительного пузырька, затем в вентральных частях зрительной чаши и зрительного стебелька. Позже эспрессия гена выявляется в диске зрительного нерва и вдоль него. Экспрессия гена выявляется и при дифференциации сетчатой оболочки.
В процессах эмбрионального развития большую роль играют не только явления индукции, но и ряд других процессов. В первую очередь необходимо упомянуть о роли адгезивных молекул, которые впервые были обнаружены именно при изучении развития сетчатки. Установлено, что адгезивные молекулы появляются в местах активного взаимодействия между рядом расположенными клетками, в результате чего формируются межклеточные контакты и, в частности, синаптические образования.
В настоящее время выявлено относительно небольшое количество адгезивных молекул. Мы кратко остановимся на некоторых из них.
- Невральная адгезивная молекула клетки (NCAM) состоит из полипептидной цепи, закодированной отдельным геном.
- Кадгерин 3. Три основных типа кадгерина выявлены в зависимости от места их обнаружения. Это кадгерин N (нервная ткань), кадгерин Е (эпителиальная ткань), кадгерин Р (плацента). Характерной особенностью кадгеринов является их взаимодействие со специализированными мембранными структурами клеток, межклеточными контактами, связанными с цитоскелетом.
[banner_centerrs] {banner_centerrs} [/banner_centerrs] - Аксон-связанные адгезивные молекулы. Несколько отличающихся типов этих адгезивных молекул было выделено у разных животных. Наиболее полная информация существует относительно одного из них. а имен но L1. Предполагают, что эти адгезивные молекулы участвуют в ориентации растущих аксонов ганглиозных клеток. Необходимо отметить, что некоторые из адгезивных молекул относятся к иммуноглобулинам.
- Рецепторы типа интегринов. Эти адгезивные молекулы представляют собой мембранные белки и являются гетеродимерами, со стоящими из множества форм а- и (3-полипептидов. Эти адгезивные молекулы способствуют взаимодействию генов, участвующих в росте аксонов ганглиозных клеток сетчатки.
Завершая описание роли адгезивных молекул в развитии глаза, лишь укажем, что более подробные сведения об их значении мы приведем в разделах, посвященных развитию отдельных структур глаза.
Немаловажное значение в развитии глаза имеют и процессы ингибирования роста. Наиболее полно эти процессы изучены на модели развития сетчатки и формирования аксонов ганглиозных клеток сетчатки. При этом основное внимание при изучении процессов ингибирования было обращено на протеогликаны. Установлено, что внеклеточные матричные молекулы имеют большое значение в установлении направления роста аксонов ганглиозных клеток в эбриогенезе, а также при регенерации зрительного нерва после травмы. При этом матричные молекулы обладают ингибирующим действием. Из этих матричных молекул наибольшее значение имеют высоко сульфатированные протеогликаны. Многие структурные протеогликаны в большом количестве обнаруживаются в развивающейся ЦНС. Обнаружены они и в сетчатке. В зависимости от условий, сульфатированные протеогликаны способствуют росту или ингибируют рост аксонов. Наиболее подробные сведения об участии протеогликанов в регуляции роста аксонов ганглиозных клеток с формированием зрительного нерва известны относительно хондроитин сульфата и гепаран сульфата.
Ингибирующее значение протеогликанов в развитии сетчатки было показано на основании исследований, которые выявили, что наибольшее их количество обнаруживается в местах прекращения роста аксонов ганглиозных клеток сетчатки. С другой стороны, ряд протеогликанов, в частности ламинин, способствует в определенных условиях росту аксонов. Протеогликаны могут быть и регуляторами дифференциации аксонов ганглиозных клеток.
Развитие эмбриона тесно связано не только с размножением, дифференциацией, перемещением клеток, но и с их дегенерацией и гибелью. Гибель клеток четко прослеживается при формировании хрусталика, эмбриональной щели, зрительного нерва и при дифференциации каждого типа нейронов сетчатки. Примеров биологического значения смерти клеток в эмбриогенезе известно довольно много. Эти процессы происходят при дифференциации любых структур организма — центральной нервной системы, желудочно-кишечного тракта, конечностей и т.д. Проявления гибели клеток могут быть в нескольких формах. В сетчатке основной формой гибели клеток является апоптоз, который завершается фагоцитозом дегенеративно измененных клеток. Морфологические проявления апоптоза хорошо описаны. Основным отличием его от других типов гибели клеток является отсутствие реакции окружающих клеток и организма на гибель клеток. Никогда при этом не обнаруживаются клетки воспалительного инфильтрата и макрофаги. Эти особенности характерны для развивающейся сетчатки и для других структур глаза. Явления апоптоза выявляются на всех этапах эмбрионального развития глазного яблока.
Признаки апоптоза обнаруживаются уже при формировании зрительного бокала, зрительной щели, зрительного стебелька и хрусталика. Запрограммированная смерть клеток приводит к инвагинации и изменению формы зрительного зачатка. Участвует этот процесс и в направлении роста аксонов ганглиозных клеток по направлению наружного коленчатого тела.
Большое значение гибель клеток имеет в дифференциации сетчатки по слоям. Потеря клеток сетчатки в процессе дифференциации исключительно высокая. По данным разных авторов, у человека погибает 70% ганглиозных клеток сетчатки. Необходимо отметить, что эспериментально на мышах показано, что апоптоз нейронов сетчатки и наружного коленчатого тела стимулируется светом в ближайшие дни после рождения.
Апоптоз участвует и в «исправлении ошибок» эмбрионального развития. Например, апоптоз удаляет нейроны и их аксоны при неправильной их проекции на перекрест зрительного нерва. Правда, необходимо отметить, что такие ошибки встречаются лишь в 1—4%. Апоптоз участвует и в регуляции распределения ганглиозных клеток по площади сетчатки.
Приступая к изложению материала о закономерностях развития глаза, целесообразно первоначально напомнить начальные этапы развития эмбриона вообще.
После оплодотворения яйцо приступает к интенсивному делению. В течение первых суток оно делится на два почти одинаковых бластомера, а еще через сутки уже образуется 3—4 клетки (рис. 5.1.1).
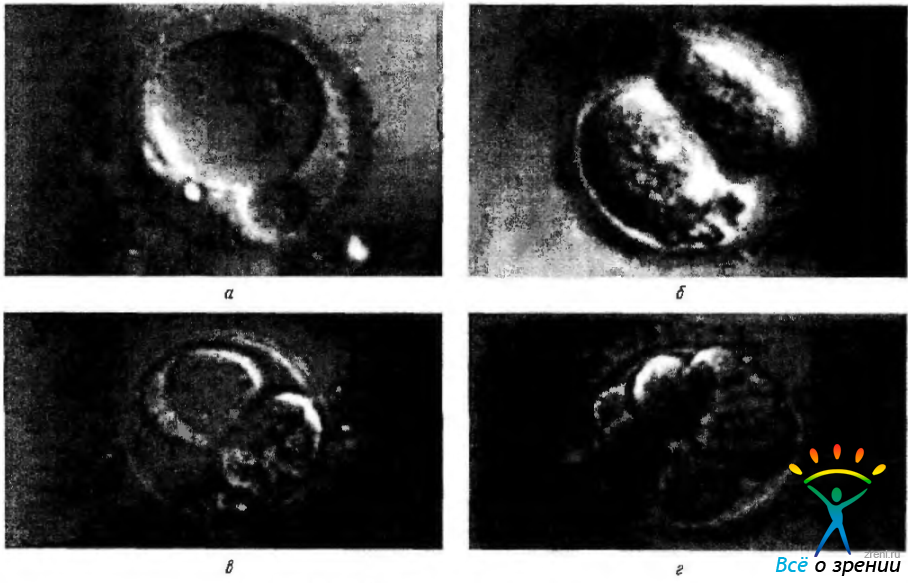
Рис. 5.1.1. Дробление яйцеклетки человека: а—оплодотворенная яйцеклетка; б—стадия двух бластомеров; в—стадия четырех бластомеров; г—стадия восьми бластомеров
На четвертые сутки зародыш достигает стадии клеточной морулы, складывающейся из 8—12 клеток.
Дальнейшее развитие зародыша происходит уже в матке. По мере быстрого размножения и дифференцировки клеток, отмечающихся в течение трех суток, к середине шестых суток зародыш состоит уже из более чем 100 клеток, а между клетками скапливается содержащая белок жидкость. При этом образуется полость и морула превращается в раннюю бластоцисту. Клетки бластоцисты становятся неоднородными и разделяются на поверхностные бластомеры, дающие начало трофобласту, и лежащие внутри более крупные и темные бластомеры. Совокупность этих клеток называется эмбриобластом (рис. 5.1.2, 5.1.3).

Рис. 5.1.2. Начальные этапы развития эмбриона: а—е — стадии интенсивного деления оплодотворенной клетки с образованием морулы (е); ж — бластоциста

Рис. 5.1.3. Ранние этапы развития эмбриона (по Пэттен, 1959): а — бластодермический пузырек; 6 — дифференциация внутренней клеточной массы (эмбрион Хергига—Рокка); в— 12-е сутки развития (эмбрион Миллера). Видны желточный мешок и амнион; г—13—14-е сутки (эмбрион Петерса). Внезародышевый целом выполнен мезодермальными клетками. Виден двуслойный эмбрион; д—15-дневный эмбрион. Видна первичная полоска и зачаток эмбриональной мезодермы (1 — плазмодиотрофобласт; 2 цитотрофобласт; 3 соединительная ткань хориона; 4 поддерживающий стебелек; 5 — полость амниона; 6—первичная полоска; 7 — зачаток эмбриональной мезодермы; 8 — энтодерма; 9 — желточный мешок; 10 — внеэмбриональная мезодерма; 11—эктодерма зародышевого щитка)
Из трофобласта в дальнейшем образуется плацента и поддерживающие ткани. Из эмбриобласта формируется эмбрион. В эмбриобласте можно четко различить два слоя — эпибласт (эктодерма) и гипобласт (энтодерма).
Таким образом, незадолго до имплантации в матку возникают два первых зародышевых листка: эктодерма, точнее, первичная эктодерма, и энтодерма. С образованием первых двух зародышевых листков и начинается процесс гаструляции.
На стадии бластоцисты зародыш человека вступает в период имплантации.
На двенадцатый день развития эмбрион состоит из двух неодинаковых по величине пузырьков. Эти пузырьки соприкасаются между собой в одном месте, образуя зародышевый щиток, из которого затем будет формироваться само тело зародыша.
К пятнадцатым суткам наступает вторая стадия гаструляции. Она характеризуется образованием среднего зародышевого листка (мезодермы), а также хорды и нервной трубки. Средний зародышевый листок образуется следующим образом. По средней линии зародышевого щитка, начиная от заднего его края, образуется сгущение клеток — первичная полоска, которая растет вперед. Дойдя до середины зародышевого щитка, она заканчивается первичным, или гензеновским узелком. Из первичной полоски путем врастания ее клеток в потенциальное пространство между эктодермой и энтодермой образуется средний зародышевый листок (мезодерма) (рис. 5.1.3—5.1.6).
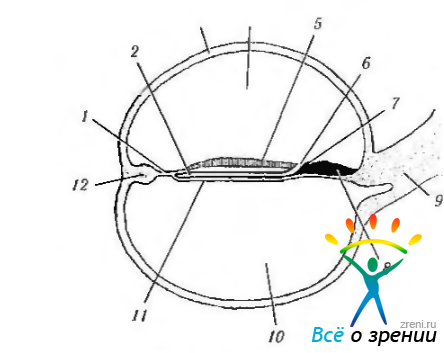
Рис. 5.1.4. Продольный срез эмбриона человека на 19-й день развития (по Duke-Elder, 1963): 1—прохордальная пластинка; 2—нотохордальная пластинка; 3— амнион; 4— амниотическая полость; 5 — нейральная пластинка; 6 бластопора; 7 узелок Хенсена; 8— примитивная полоска; 9 — зародышевый ствол; 10— желточный мешок; 11 — энтодерма; 12 — эктодерма
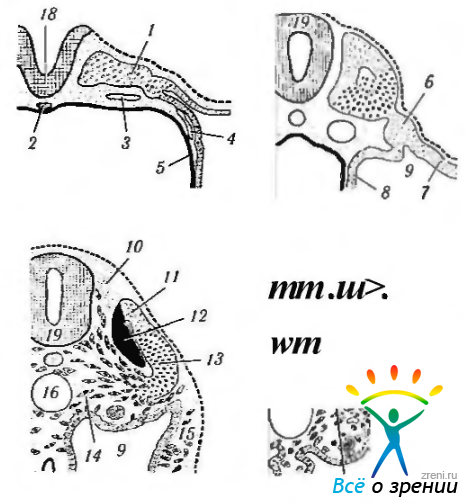
Рис. 5.1.5. Различные стадии образования нервной трубки из нервной пластинки, а также формирование мезодермы (по Clara, 1955): а — зародыш на стадии 5—6 сомитов; 6— зародыш на стадии 13—14 сомитов; в, г — зародыш на более поздних стадиях развития (1 — первичный сегмент; 2 — хорда; 3 — левая аорта; 4 — кровеносный сосуд; 5 — энтодерма; б — сегментная ножка; 7 — париетальный листок спланхнотома; 8— висцеральный листок спланхнотома; 9—вторичная полость тела; 10—спинальный ганглий; 11—дорзальный край первичного сегмента; 12 — миотом; 13 — кожная пластинка (дерматом); 14 — выселение клеток мезенхимы из склеротома; 15 — выселение клеток мезенхимы из париетального листка спланхнотома; 16 — аорта; 17 — выселение клеток мезенхимы из дерматома; 18 — нервная пластинка; 19 — нервная трубка)

Рис. 5.1.6. Схематическое изображение дорзальной поверхности эмбриона человека. иллюстрирующее этапы развития нервной пластинки, желобка и трубки (по Sadler, 1990): а—19-й день развития. Нервный желобок представляет собой углубление, расположенное в центре нервной пластинки; 6— 21-й день развития. Отмечается смыкание краев желобка с началом образования трубки; в, г 22-й и 23-й дни развития. Дальнейшее формирование нервной трубки путем прогрессивного распространения ее закрытия по направлению к переднему и заднему концам эмбриона (1 - обрезанный край амниона; 2 — нервная пластинка; 3—примитивный узелок; 4 — примитивная полоска; 5—нейральная складка; 6—перихондральное выпячивание; 7—слуховая плакода; 8—задняя нейропора)
На семнадцатый день развития из гензеновского узелка по направлению к переднему краю зародышевого щитка вырастает клеточный тяж — головной, или хордальный, отросток, представляющий собой зачаток будущей спинной струны (хорды).
В начале третьей недели развития эмбрион имеет трехслойную структуру и состоит из эктодермы, мезодермы и энтодермы.
Между перечисленными листками располагаются рыхло лежащие клетки звездчатой формы — мезенхима, погруженные в межклеточную белоксодержащую жидкость с небольшим количеством волокон. Клетки мезенхимы исходят из различных участков мезодермы дерматомов, склеротомов, спланхнотомов. Мезенхима по клеточному составу довольно рано становится гетерогенной в результате миграции в нее клеток другого происхождения (нейробласты, меланоциты, миобласты и др.). В области головы и шеи мезенхима развивается из клеток нейрального гребня, о чем более подробно будет изложено ниже. Мезенхима является основным источником формирования соединительной ткани в организме.
В этот промежуток времени эктодермальные клетки, расположенные вдоль тела эмбриона на его дорзальной поверхности, начинают интенсивно размножаться. Первоначально формируется утолщение, называемое нейральной пластинкой. Нейральная пластинка, углубляясь, превращается в бороздку. Края (валики) нейральной пластинки постепенно становятся все выше, а затем срастаются между собой, превращая бороздку в нейральную трубку (рис. 5.1.4—5.1.6). Нейральная трубка представляет собой зачаток части центральной нервной системы.
Клеточный материал нейральной трубки, обособляясь от эктодермы в дальнейшем развивается в элементы нервной системы (нейроны, глия). По этой причине ткань нервной трубки называется нейроэктодермой.
В переднем отделе нейральной трубки благодаря интенсивному размножению клеток нейроэктодермы формируется три первичных мозговых пузырька, лежащих друг за другом:
- передний (prosencephalon),
- средний (mesencephalon)
- и задний (rhombencephalon).

Рис. 5.1.7. Схематическое изображение последовательности развития головного мозга (по Duke-Elder, 1963): а — первичный мозговой пузырь; б— стадия трех мозговых пузырей; в—стадия пяти мозговых пузырей; г—стадия конечной дифференциации теленцефалона (1—prosencephalon; 2— готbencephalon; 3 — mesencephalon: 4 — medulla oblongata, 5 — обонятельная доля; 6 — зрительный бугор: 7 — telencephalon; 8 — мозжечок; 9 — diencephalon; 10 — полушария мозга; 11 — полосатое тело: I, II, III, IV—первый, второй, третий и четвертый желудочки)
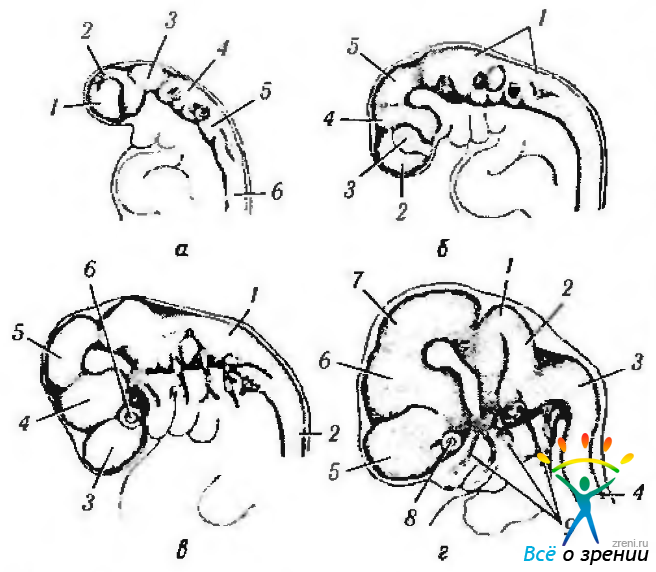
Рис. 5.1.8. Четыре стадии развития головного мозга и черепномозговых нервов у эмбрионов человека (по Пэттену, 1959): а — эмбрион Дэвиса (3,5 неделя развития) (1 — глазной пузырь; 2 — передний мозг; 3 — средний мозг; 4 — задний мозг; 5 — слу ховая ямка; 6 — спинной мозг); б— 4-я неделя развития эмбрио на (1 — задний мозг; 2 — конечный мозг; 3—глазной пузырь; 4 — промежуточный мозг: 5 — средний мозг); в — начало 6-й недели развития эмбриона (1 — задний мозг; 2 — спинной мозг; 3 — пузырь конечного мозга; 4 — промежуточный мозг; 5 — средний мозг; 6 глазной пузырь); г — 7-я неделя развития эмбриона (1—задний мозг; 2 — ромбическая губа; 3 — продолговатый мозг; 4 — спинной мозг; 5 — пузырь конечного мозга; 6 — про межуточный мозг; 7 — средний мозг; 8— зрительный пузырь; 9— черепномозговые нервы)
Наибольшей дифференциации подвергается передний мозговой пузырек (prosencephalon). Он подразделяется на заднюю часть, diencephalon (промежуточный мозг), и переднюю, telencephalon (конечный мозг). Именно из боковых стенок промежуточного мозга и развивается глазное яблоко.
Первые признаки развития глазного яблока выявляются уже на третьей неделе эмбрионального развития (эмбрион 2 мм). Первоначально с двух сторон от срединной линии в вентролатеральной области промежуточного мозга (prosencephalon) появляются два выпячивания, направленных в сторону эктодермы. Постепенно увеличиваясь, они формируют два шаровидных образования — первичные зрительные пузырьки (рис. 5.1.10—5.1.12).
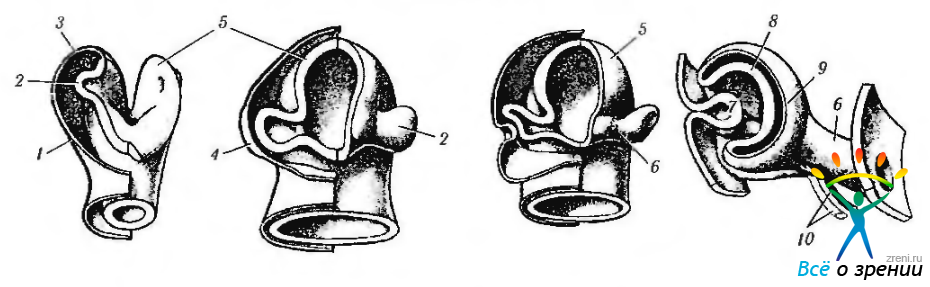
Рис. 5.1.10. Стадии формирования глазного пузырька и глазного бокала: а — 21-й день эмбрионального развития (2,0 мм); б — 27-й день эмбрионального развития (4,5 мм); в — 29-й день эмбрионального развития (7,0 мм); г — 31-й день эмбрионального развития (7,5 мм) (1— стенка переднего мозга; 2— зрительный пузырек; 3— поверхностная эктодерма; 4 — хрусталиковая плакода; 5— передний мозг; 6 — глазной стебелек; 7 — хрусталиковых пузырек; внутренний слой глазного бокала; 9 — наружный слой глазного бокала; 10—артерия и вена (гиалоидные))
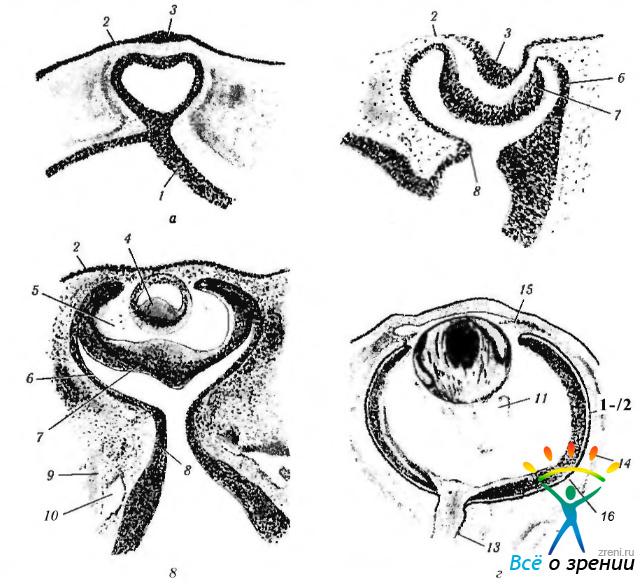
Рис. 5.1.11. Микроскопическая картина последовательных стадий развития глазного бокала (рисунок): а — стадия глазного пузырька: б — стадия глазного бокала: в, г — стадии, на которых отмечается формирование хрусталика и дифференциация других структур глаза (1—нейроэктодерма; 2—поверхностная эктодерма; 3—хрусталиковая плакода; 4—хрусталиковый пузырек; 5 — окружающая глазной пузырек мезенхима; 6 — наружная стенка глазного бокала; 7 — внутренняя стенка глазного бокала; 8—глазной стебелек; 9 — окружающая мезенхима; 10 — сосуды, располагающиеся в мезенхиме; 11— первичное стекловидное тело; 12 — пигментный эпителий сетчатки; 13 — зрительный нерв; 14 — склера; 15 — радужная оболочка; 16 — сосудистая оболочка)
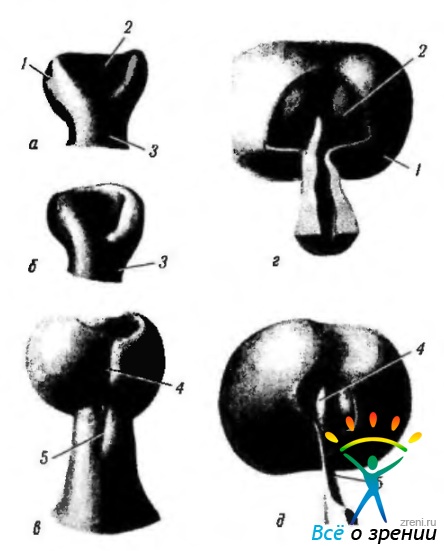
Рис. 5.1.12. Этапы развития глазного бокала (слепок). На подобных препаратах четко определяется образование и закрытие эмбриональной щели, распространяющейся на зрительный стебелек (по Velhagen, 1958): а—11-й день развития; б 11,5 дня развития; в—13-й день развития; г— 14-й день развития; д— 15-й день развития (а, б, в — вид с внутренней стороны; г, д— наружная стенка глазного бокала в области глазного стебелька удалена) (1—наружная стенка глазного бокала; 2 внутренняя стенка глазного бокала; 3 — зрительный стебелек; 4 — эмбриональная щель в области глазного бокала; 5 — эмбриональная щель в области зрительного нерва)
При закрытии нейральной трубки зрительные пузырьки увеличиваются и уже имеют вид полостей, симметрично расположенных на боковой стороне мозгового пузыря. Полость зрительного пузырька связана с полостью мозга. Первоначально эпителиальные клетки зрительного пузырька имеют призматическую форму. Их ядра образуют несколько слоев, число которых постепенно увеличивается. Все клетки, выстилающие зрительный пузырек, ориентированы апикальной поверхностью внутрь пузырька и содержат реснички. Базальная поверхность клеток располагается на тонкой базальной мембране.
Зрительный пузырек контактирует с поверхностной эктодермой и продолжает быть связанным с головным мозгом посредством сужения, которое называется зрительным стебельком. Зрительный стебелек соединяется с наиболее нижней частью стенки промежуточного мозга, а полость промежуточного мозга (будущий третий желудочек) сообщается с полостью зрительного стебелька.
Приблизительно к 27-му дню развития (эмбрион 4,0—4.5 мм) поверхностная эктодерма, расположенная над зрительным пузырьком, утолщается и образуется хрусталиковая плакода. Взаимодействие внеклеточных элементов глазного зачатка стимулирует постепенную инвагинацию хрусталиковой плакоды в полость глазного пузырька (рис. 5.1.10, 5.1.11). Первоначально зачаток хрусталика соединен с поверхностной эктодермой при помощи хрусталикового стебелька. В конечном счете, хрусталиковый пузырек полностью отделяется от поверхностной эктодермы. Необходимо отметить, что образование хрусталикового пузырька можно увидеть на поверхности эмбриона. Выглядит он маленьким углублением (хрусталиковая ямка или пора) (рис. 5.1.9).
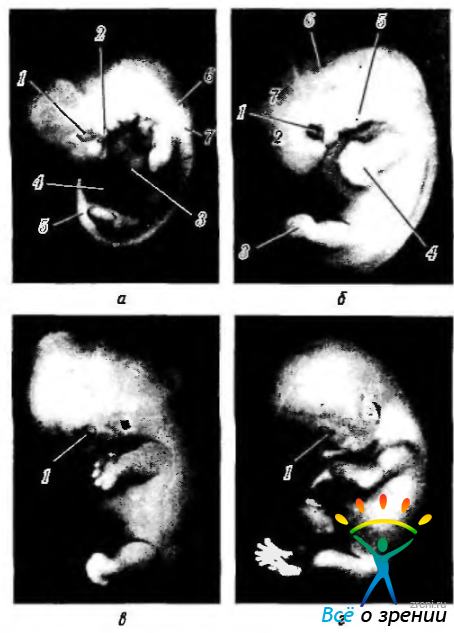
Рис. 5.1.9. Стадии формирования плода и развитие при этом глазного яблока: а — эмбрион на 5-й неделе (5 мм) (1 — закладка глазного бокала; 2 — первая жаберная дуга; 3 — сердечное выпячивание; 4 — закладка пуповины; 5 — хвост; 6 — вторая жаберная дуга; 7 — третья жаберная дуга); б — эмбрион в конце 6-й недели (10 мм) (1 — глазной бокал; 2 — конечный мозг; 3 — нога; 4 — рука; 5 — наружный слуховой проход; 6 — крыша IV желудочка: 7 — средний мозг); в, г — эмбрионы в конце 7-й (17 мм) и 8-й (31 мм) недель развития соответственно (1 — глазное яблоко с формированием век)
Одновременно зрительный пузырек превращается в зрительный бокал (рис. 5.1.12, 5.1.13).
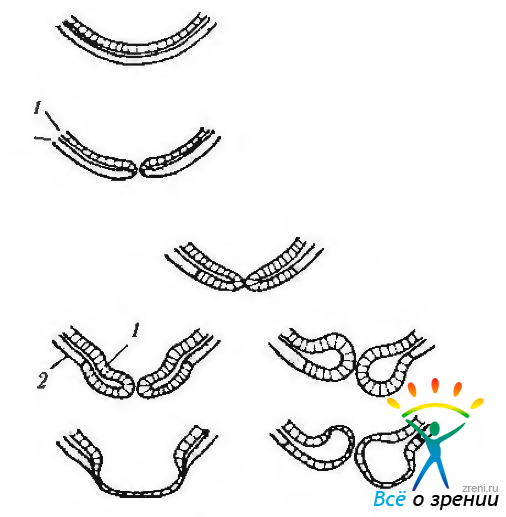
Рис. 5.1.13. Схема закрытия эмбриональной щели и нарушение этого процесса: а — закрытие эмбриональной щели в норме. Внутренний и наружный нейроэпителиальные слои приходят в соприкосновение, а затем срастаются. Формируется базальная мембрана; б— закрытие щели с образованием колобом. Эктропион внутреннего слоя нейроэпителия приводит к нарушению контакта, а затем и срастанию «губ» глазного бокала. В результате развиваются аномалии сенсорной части сетчатки и пигментного эпителия. Увеальный тракт и склера в этом месте также подвергаются недоразвитию. Справа изображены кистозные колобомы (1 — внутренний слой глазного бокала; 2 — наружный слой глазного бокала)
К 28-му дню развития (эмбрион 7,6—7,8 мм) движение клеток зрительного пузырька приводит к инвагинации височной и нижней стенок пузырька внутрь по направлению верхней и задней стенок, распространяющейся и на зрительный стебелек (рис. 5.1.13). Полость зрительного пузырька постепенно уменьшается. В том месте, где две боковые стенки бокала встречаются с вентральной стороны глазного пузырька, образуется щель, известная как эмбриональная щель (рис. 5.1.12—5.1.14).
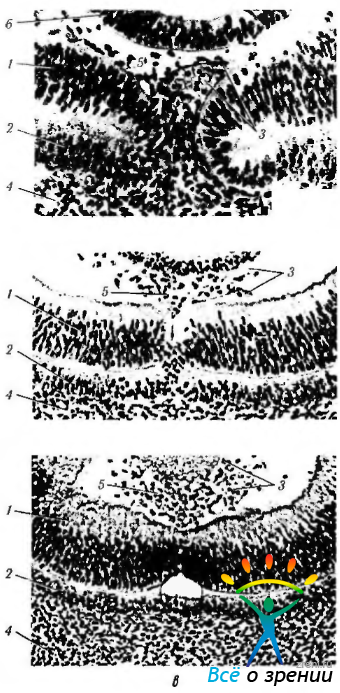
Рис. 5.1.14. Этапы закрытия эмбриональной щели в норме (световая микроскопия). Внутренний и наружный слои нейроэпителия глазного бокала сближаются, а затем и срастаются: а,б — сближение краев с сохранением между ними мезенхимы, формирующее сосуды гиалоидной системы: в — полное смыкание краев эмбриональной щели (1 — внутренний нейроэпителиальный слой глазного бокала; 2 — наружный нейроэпителиальный слой глазного бокала, постепенно превращающийся в пигментный эпителий сетчатки; 3 — мезенхима, проникающая в полость глазного бокала; 4 — мезенхима, окружающая глазной бокал и являющаяся источником образования сосудистой оболочки и склеры; 5 — кровеносные сосуды гиалоидной системы; 6 - задняя стенка хрусталиков ого пузырька)
Механизм закрытия эмбриональной щели пока полностью не изучен. Лишь в эксперименте Geeraets, Suzuki, Shirai, Majima получены определенные сведения относительно этого процесса. Он сводится к следующему (рис. 5.1.14). Край эмбриональной щели состоит из внутреннего слоя сенсорной части сетчатки и наружного слоя клеток, дифференцирующихся в пигментный эпителий. На самых ранних стадиях слияния нейроэпителиальных слоев наружный слой глазного бокала (будущий пигментный эпителий сетчатки) проникает в щель. Эти недифференцированные клетки участвуют в процессе слияния нейроэпителиальных слоев эмбриональной щели. Только после того, как слияние произошло, в клетках наружного слоя появляются меланосомы. По мере сближения краев зрительного бокала развиваются многочисленные оппозиционные контакты между клетками. Поскольку нейральная часть сетчатки отделена от пигментного эпителия, формируется непрерывный ряд межклеточных контактов со смежными клетками наружного слоя (будущий слой фоторецепторов). Смежные клетки пигментного эпителия сетчатки соединяются при помощи десмосом. расположенных на их апикальной поверхности. На боковых поверхностях этих клеток видны контакты промежуточного типа.
В процессе слияния краев эмбриональной щели происходит довольно интенсивная гибель части клеток. При этом используется механизм «запрограммированной смерти», т. е. апоптоз. Способствуют гибели клеток и выведению продуктов распада также многочисленные амебоподобные клетки, отличающиеся выраженной фагоцитарной активностью. Наибольшее их количество выявляется именно в месте слияния краев глазного бокала.
Эмбриональная щель имеет большое значение в дальнейшем развитии глазного яблока. Ее наличие обеспечивает рост аксонов ганглиозных клеток сетчатки по направлению к головному мозгу с образованием зрительного нерва. Кроме того, именно благодаря эмбриональной щели в зрительный бокал проникает мезенхима, являющаяся источником образования первичного стекловидного тела.
Как указано выше, эмбриональная щель постепенно закрывается. Первоначально это происходит в ее средних участках. Затем закрытие распространяется кпереди и кзади. В тех случаях, когда эмбриональная щель не закрывается полностью, образуются типичные колобомы оболочек глаза (рис. 5.1.13). Более часто встречаются колобомы радужки, а также заднего отдела глаза и зрительного нерва. В экваториальной области колобомы редки. Эта закономерность связана именно с последовательностью закрытия эмбриональной щели.
К моменту закрытия эмбриональной глазной щели (седьмая неделя) большинство структур глаза уже существует. С этого момента развитие глаза сводится к дифференциации и модификации различных частей глазного яблока, что подробно будет рассматриваться далее.
Перед тем как приступить к изложению особенностей развития отдельных структур глаза, необходимо привести сведения относительно динамики изменения размера, веса глазного яблока и эмбриона (табл. 5.1.1, 5.1.2).
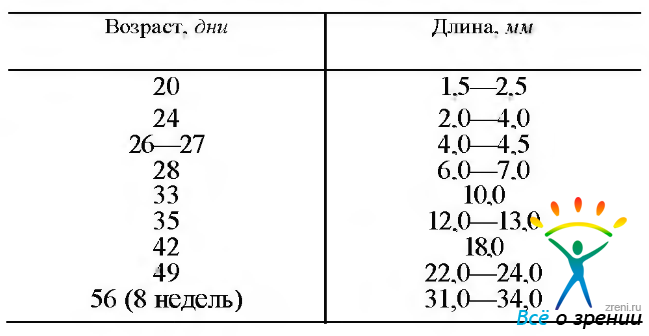
Tаблица 5.1.1. Усредненные размеры эмбриона на ранних стадиях развития человека

Таблица 5.1.2. Относительные размеры и вес эмбриона и глаза человека
Эти цифры упоминаются во многих работах, поскольку они характеризуют качественно отличающиеся этапы развития эмбриона. Получены они на основании изучения эмбрионов человека многими исследователями.
Завершая этот раздел, необходимо подчеркнуть, что в отличие от других органов в развитии глаза принимают участие только два из трех зародышевых листка: эктодерма (нейроэктодерма, поверхностная эктодерма) и мезодерма. Важно отметить, что мезодерма в области головы и шеи имеет свои особенности происхождения. На этом необходимо остановиться более подробно.
Термином «мезенхима» обозначают «примитивную» соединительную ткань, которая не образует зародышевого листка (слоя), а представляет собой рыхлое скопление клеток звездчатой формы, выполняющих пространства между зародышевыми листками. Мезенхима в области туловища развивается из мезодермы путем выселения клеток на ранних стадиях развития. Мезенхимные структуры головы (включая структуры глаза и глазницы) исходят из двух источников — клеток нейрального гребня и мезодермы. Именно в последние три десятилетия было выявлено значение клеток нейрального гребня в органогенезе глаза и структур глазницы.
Клетки нейрального гребня возникают в момент закрытия нейральной трубки. Они выявляются у вершины нервных валиков, расположенных вдоль нервной пластинки (рис. 5.1.15).
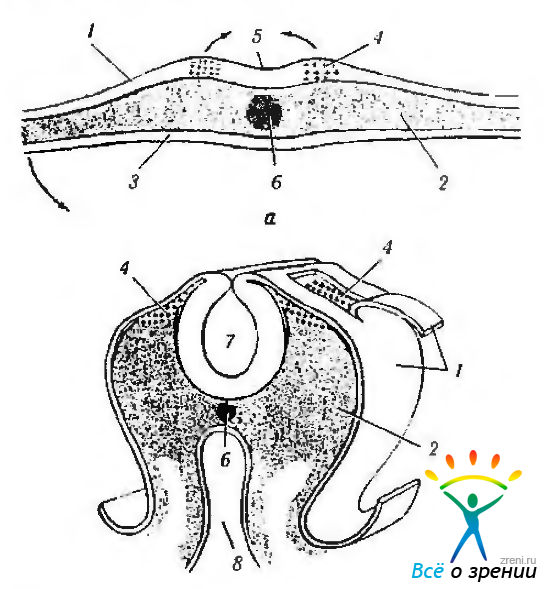
Рис. 5.1.15. Поперечный срез эмбриона. Миграция клеток нейрального гребня (на схеме обозначены крестиками) (по Johnston, Sulik, 1980): а — начальная стадия формирования нейрального желобка; б— образование нервной трубки (1—эктодерма; 2—мезодерма; 3 — энтодерма; 4 — нейральный гребень; 5 — нейральная пластинка; 6 — нотохорда: 7 — нейральная трубка; 8 — желудочно-кишечная трубка)
Клетки нейрального гребня дают начало ряду образований организма человека. К этим образованиям относятся нейробласты (внутричерепные, спинальные и периферические ганглии), глия и спонгиобласты (все глиальные элементы периферической нервной системы), парасимпапткобласты (парасимпатические элементы центральной и периферической нервной системы), меланобласты (меланоциты, невусные клетки), центральные и периферические нервные элементы (с-клетки, гипофиз, лобная доля, клетки желудочно-кишечного тракта), феохромобласты (симпатические ганглии, адреналовая система) и производные нейрального гребня в области головы и шеи.
Процесс выселения и миграции клеток нейрального гребня происходит следующим образом (рис. 5.1.16).
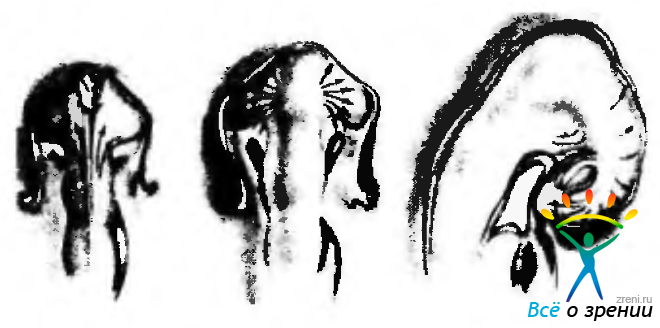
Рис. 5.1.16. Миграция клеток нейрального гребешка (по Johnston, 1966) (направление миграции указано стрелками): а — в процессе закрытия нервной трубки клетки нейрального гребешка перемещаются вперед: б— на следующем этапе клетки нейрального гребешка направляются к глазному пузырьку и окружают его; в — в последующем клетки мигрируют в вентральном и дорзальном направлениях, участвуя в формировании соединительной ткани лицевого черепа
Клетки нейрального гребня, лежащие в передних отделах промежуточного и среднего мозга, начинают передвигаться (мигрировать) вперед вдоль дорзальной поверхности эмбриона. Эта миграция происходит «волнообразно». При этом пузырек окружается ими с дорзальной, вентральной и медиальной сторон. В дальнейшем часть клеток перемещается каудально. формируя при этом верхнечелюстной отросток. Другие клетки мигрируют рострально, образуя лобно-носовой отросток. Две волны мигрирующих клеток встречаются и смешиваются вокруг глаза. Указанные отростки (верхнечелюстной и лобно-носовой) являются основой формирования части лицевого черепа и мягких тканей, о чем будет подробней изложено в соответствующих разделах (эмбриогенез глазницы, век).
На ранних стадиях миграции клетки нейрального гребня плюрипотентны, т. е. в дальнейшем они могут дифференцироваться в различных направлениях. Из них развиваются соединительнотканные компоненты глаза и глазницы. Это строма роговой оболочки и эндотелий, большинство структурных элементов склеры, зрительного нерва, строма сосудистой оболочки, ресничного тела, включая мышцы, строма радужки, кости глазницы, жировая клетчатка, хрящ блока, нервные, мышечные оболочки и соединительная ткань глазницы (табл. 5.1.3).
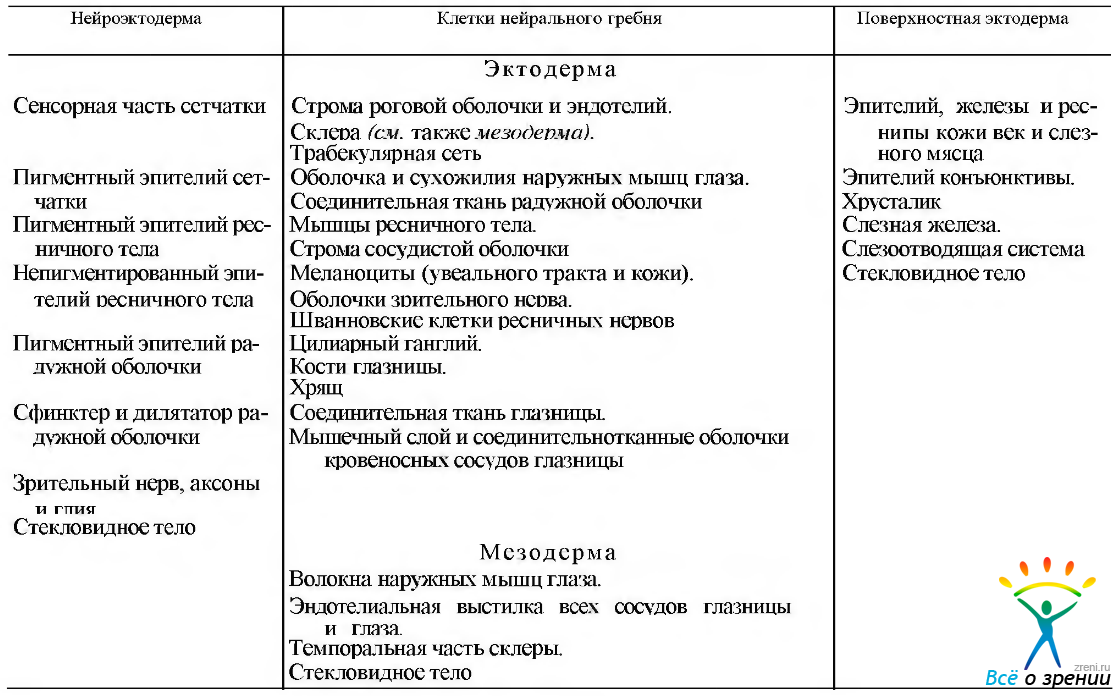
Таблица 5.1.3. Особенности происхождения структур глаза и орбиты
Исключением являются поперечнополосатые мышечные волокна наружных мышц глаза и эндотелиальная выстилка всех сосудов глаза и глазницы. Наружные мышцы глаза, как и круговая мышца глаза, развиваются из мезодермы. но соединительнотканные элементы этих мышц — из клеток нейрального гребня. Клетки нейрального гребня также принимают участие в развитии сосудистой системы эмбрионального глаза, о чем более подробно приведены данные в разделе, посвященном увеальному тракту.
----
Статья из книги: Строение зрительной системы человека | Вит В. В.

Комментариев 0