Взаимодействие родопсина с цитоплазматическими белками (трансдуцином и ареатином) в фоторецепторной клетке (ЧАСТЬ 2)


Описание
Влияние модификации сулъфгидрильных групп на взаимодействие арестина с родопсином.Молекулу родопсина модифицировали MSL либо HgSL, а SH-группы арестина - HgSL. Фото- рецепторные мембраны, содержащие меченный или намеченный фосфорилированный родопсин, инкубировали с нативным или модифицированным арестином 20 мин при 20° С, затем мембраны саждали центрифугированием и определяли концентрацию не связавшегося арестина. Результаты опытов приведены в табл. 3.3.

Из таблицы видно, что модификация двух остатков родопсина MSL или двух других остатков HgSL практически не влияет на степень связывания арестина с родопсином, а константы диссоциации (Кд) комплексов арестина с родопсином, модифицированным MSL или HgSL, незначительно отличаются от величин Кл, определенных в контрольных экспериментах с немодифицированным родопсином. Таким образом, четыре поверхностных остатка цистеина Cys-140, Суs-316, Cys-322 и Cys-323, по-видимому, не участвуют в формировании центра связывания арестина на фосфорилированном родопсине. Примечательно, что и центр связывания cx-субъединицы транодуцина на родопсине не содержит SH-групп.
Модификация одной SH-группы арестина HgSL также не влияет на его связывание с фосфорилироваьным родопсином (см. табл. 3.3), а при более глубокой модификации связывание арестина лишь незначительно уменьшается: константа диссоциации возрастает не более чем на 25-30%. По-видимому, остатки цистеина не располагающая в области молекулы арестина, контактирующей с родопсином. Это принципиально отличает арестин от ?-субъединицы транедуцина, модификация одной из SH-rpynn которого нарушает его взаимодействие с родопсином.
Интересно, что в аминокислотной последовательности арестина имеется участок, гомологичный участку ?-субъединицы гранедуцина, который, как предполагается, связывается с родопсином. Так, степень гомологии участков 371-391 арестина и 326-350 транедуцина превышает 50%. Более того, секстапептид 384—389 арестина полностью совпадает с пептидом 343-3ч8 транедуцина, за исключением остатка Cys-347 гранедуцина. который заменен на Аlа-380 в арестине. Полученные нами данные не противоречат предположению о том, что именно этот участок арестина может быть центром связывания с родопсином. То обстоятельство, что модификация SН-групп арестина не влияет на его взаимодействие с родопсином, по-видимому, есть результат именно этой аминокислотной замены в структуре арестина, которая может иметь функциональное значение.
Определение Кд комплексов арестина и трансдуцина с фосфорилированными и нефосфорилированными формами родопсина. Чтобы объяснить ингибирующее действие арестина его непосредственным связыванием с фосфорилированным родопсином, следует предположить, что сродство между этими белками достаточно высоко. Нами; проведено сравнительное изучение степени связывания арестина с фосфорилированными и нефосфорилированными формами родопсина, метародопсина II и опсина. Оказалось, что с необесцвеченным родопсином связывается не более 2-5% арестина при концентрациях этих белков соответственно 40 и 2.5 мкМ, т.е. константа диссоциации (Кд) комплекса составляет 0,7-2 мМ. Фосфорилирование родопсина практически не влияет на сродство к нему арестина: степень связывания арестина с фосфорилированным и регенерированным родопсином не превышает 4-6%.
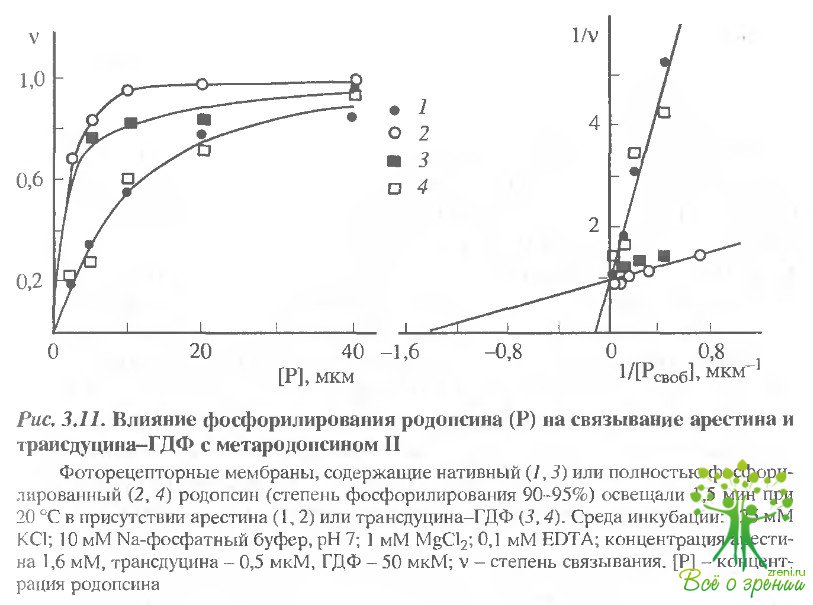
Образующийся при освещении фоторецепторных мембран метародопсин II существенно лучше связывает арестин (рис. 3.11). Кроме того, эффективность связывания значительно возрастает в результате фосфорилирования родопсина: Кд комплекса уменьшается от 11,7 ± 0,1 до 0,7 ± 0,1 мкМ, т.е. более чем на порядок.
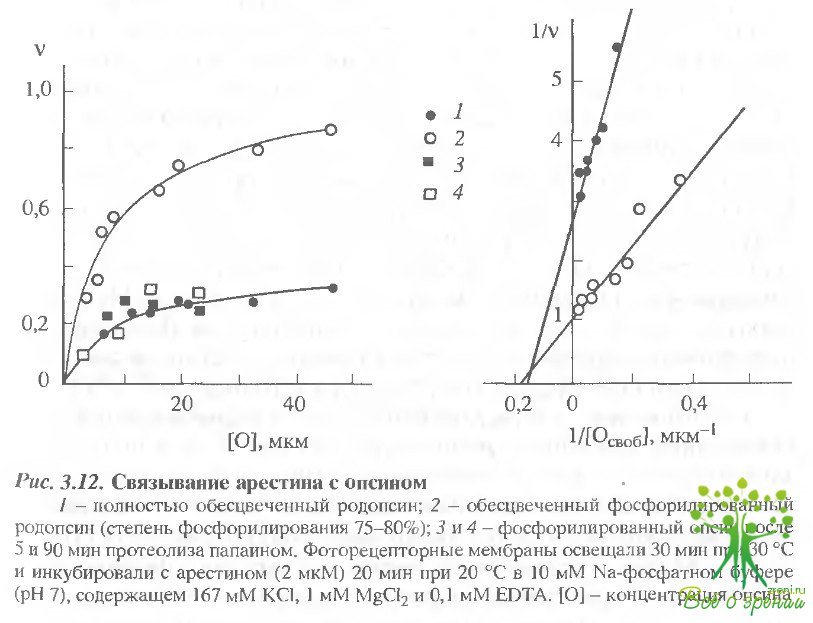
При распаде метародопсина II сродство арестина к родопсину снова падает: величины Кд для комплексов с нефосфорилированным и фосфорилированным опсином составляют соответственно 6,3 ± 2,5 и 5,3 ± 1,5 мкМ (рис. 3.12). Кроме того, при связывании с опсином проявляется структурная гетерогенность арестина: если с фосфорилированным опсином связывается до 100% молекул арестина, то с нефосфорилированной формой - не более 30- -35%.
Такая гетерогенность может быть обусловлена различием физико-химических свойств "популяций" арестина, возможно, наличием агрегированных форм белка. О гетерогенности молекул арестина по заряду свидетельствует картина его изоэлектрофокусирования (рис. 3.13): как отмечалось и ранее, обнаруживаются три основные полосы (pH 5,5, 5,65 и 5,7) и 4 минорные полосы (pH 5,43, 5,5, 6,0 и 6,1).

Аналогичным образом были определены величины Кд комплексов трансдуцина-ГДФ с метародопсином II в фосфорилированном и нефосфорилированном состояниях (см. рис. 3.11). Для комплекса с нефосфорилированным метаро-допсином II Кд составила 0,7 ± 0,2 мкМ, что близко к значению, полученному в работе путем измерения светорассеяния. При фосфорилировании метародопсина II его сродство к трансдуцину-ГДФ падает: Кд = 11,2 ± 0,2 мкМ. Таким образом, при фосфорилировании метародопсина II образуется новый высокоаффинный центр связывания арестина, и одновременно уменьшается сродство рецептора к трансдуцчну-ГДФ.
Исследование электростатических взаимодействий при связывании арестина с родопсином.На рис. 3.14, а приведены зависимости степени связывания арестина с фосфорилированным и нефосфорилированным опсином от ионной силы среды инкубации.
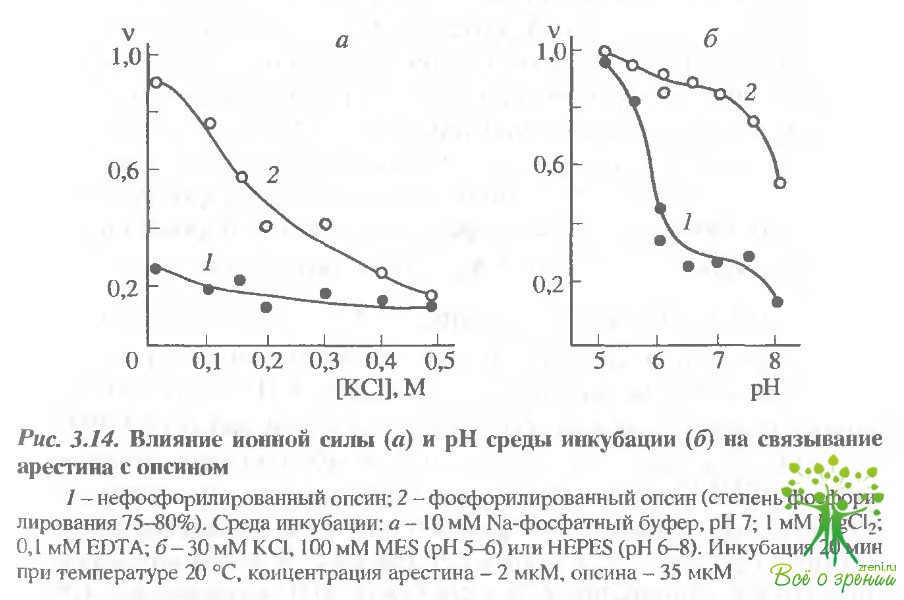
Видно, что при увеличении концентрации KCl от 30 до 500 мМ степень связывания падает, и в итоге обе формы о пси на становятся практически неразличимыми по своему сродству к арестину. Это означает, что взаимодействие между белками но сит в значительной степени электростатический характер, т.е. при увеличении концентрации электролита связывание арестина уменьшается за счет экранирования зарядов на молекулах белков. В то же время степень связывания арестина с нефосфорили-рованным опсином слабо зависит от ионной силы среды инкубации (см. рис. 3 14, а, кривая У), что указывает на отсутствие заряженных групп в местах взаимодействия этих белков.
Вывод об электростатической природе взаимодействия арестина с фосфорилированным опсином следует также из рН-зависимостей степени связывания (см. рис. 3.14, б). При уменьшении рН от 8 до 5 Кд комплекса арестина с фосфорилированным опсином уменьшается примерно в 30 раз, причем, наиболее резкие изменения наблюдаются при рН 5,6 и 7,6. Перегибы в этих точках, по-видимому, соответствуют изменению заряда именно молекул арестина, поскольку они наблюдаются как для фосфорилированного, так и для нефосфорилированного опсина. Кроме того, основных полос при изоэлектрофокусировании арестина соответствует диапазону 5,6-5,7 (см. рис, 3.13), а р групп родопсина, обнаруживаемых при титрование фоторецепторных мембран, как известно, равен 6,7.
Характер взаимодействия арестина с фосфорилированным и нефосфорилированным опсином. Характер связывания арестина с родопсином был изучен также методом ЭПР при помощи ртутьорганической спиновой метки, ковалентно связанной с одной из наиболее реакционноспособных SН-групп арестина. Введение метки не изменяет степени связывания арестина с родопсином. Спектр ЭПР спинмеченного арестина (рис. 3.15) состоит из двух сигналов, один из которых соответствует иммобилизованному относительно белка состоянию метки (S), а другой (W) - ее достаточно свободному вращению (время корреляции в модели изотропного вращения примерно соответствует 0,3 нc).
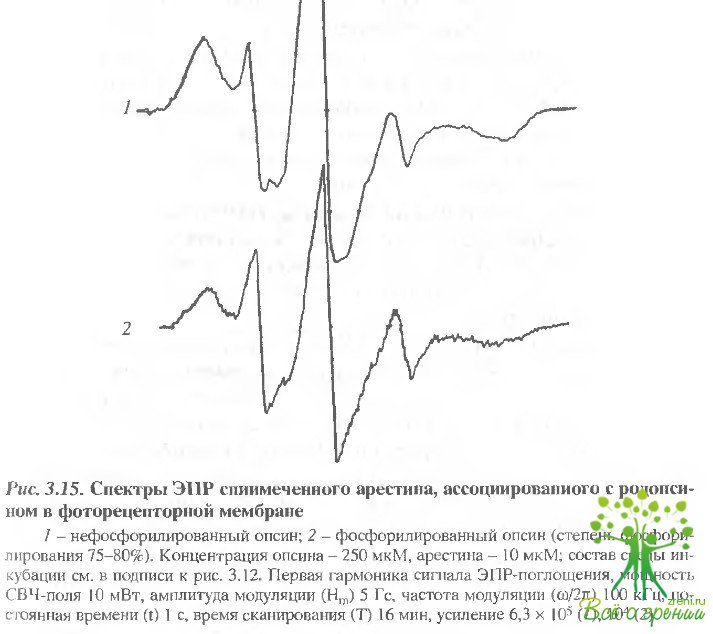
Оказалось, что связанный с опсином арестин сохраняет вращательную подвижность, причем она зависит от фосфорилирования: для комплекса с нефосфорилированным и фoсфорилированным опсином параметр 2 A’zz равен соответственно 64,8 ± 0,2 и 66,8 ± 0,2 Гс. Большее значение 2 A’zz для комплекса с фосфорилированным опсином нельзя объяснить большей полярностью окружения метки, поскольку спектр ЭПР этого комплекса характеризуется также и большим значением фактора СВЧ-насыщения Z+1, мало чувствительного к абсолютным значениям тензора СТВ: 2,14 по сравнению с величиной 1,49 для комплекса арестина с нефосфорилированным опсином при мощности поля СВЧ 100 мВт. В обоих случаях различаются и отношения W/8 в спектрах ЭПР комплексов: 1,80 и 0,94 соответственно для комплексов с нефосфооилированным и фосфорилированным родопсином. Это свидетельствует о конформационных изменениях в арестине при его связывании с оперном, особенно с его фосфорилированной формой.
Количественную оценку вращательной подвижности арестина, связанного с опсином, можно провести приближенно в рамках модели изотропного вращения, используя в качестве предельного (в отсутствие вращения) значение 2 A’zz = 69 Гс, полученное для арестина в агрегированном состоянии при его высокой концентрации в растворе. Время корреляции вращения арестина в комплексе с нефосфорилирэванным опсином оказалось равным 27 ± 2 нм, а с фосфорилированным опсином - 60 ± 2 не (расчетное значение для свободного белка с молекулярной массой 48 кДа в водном растворе - 19 не). Таким образом, место связывания фосфорилированного опсина с арестином отличается от центра нефосфорчлированного опсина и большей степенью иммобилизации арестина на белке-рецепторе.
Влияние протеолиза фосфорилированного родопсина на связывание с ним арестина.Связывание арестина с фосфорилированным опсином исследовали через 5 и 90 мин протеолиза, т.е. когда удалялся С-концевой фрагмент или удалялась петля между V и VI ?-спиральными тяжами. При этом величина Кд возрастала соответственно до 7,6 ± 2,8 и 10,5 ± 3,3 мкМ по сравнению с исходным значением Кд = 5,3 ±1,5 мкМ. Кроме того, уменьшалось также предельное значение доли арестина, способного связываться с родопсином до величины 35-40%, соответствующей связыванию с нефосфорилированным опсином (см. рис. 3.12).
Таким образом, как отщепление С-концевого участка, несущего фосфорилированные аминокислотные остатки, так и удаление петли между V -VI «-спиральными тяжами, снижают эффективность связывания арестина с опсином. По-видимому, оба эти участка играют существенную роль в формировании специфического центра связывания арестина с фосфорилированным рецептором.
Влияние трансдуцина на спектры ЭПР меток, связанных с родопсином. Добавление трансдуцина к суспензии спинмеченных дисков приводит к изменению подвижности меток, связанных со всеми SН-группами. Однако, в отличие от арестина, характер этих изменений другой.
В частности, подвижность МЕSLувеличивается как для Суs- 140, так и для Сук-316: температура существенно снижается (см. табл. 3.3). Метки HgSLна Суз-322, 323, напротив, затормаживаются, о чем свидетельствуют, в частности, уменьшение полуширины линии (?Н+1) и возрастание фактора насыщения (Z+1). Это свидетельствует, во-первых, о том, что и в темноте имеет место связывание трансдуцина с родопсином. Во-вторых, различия в спектральных изменениях при добавлении трансдуцина и арестина позволяют предположить, что сами места связывания этих белков не идентичны. Хотя нельзя не учитывать также, что белки имеют различную форму, размер, субъединичное строение.
Существенное влияние оказывает добавление трансдуцина и на параметры фотоответа. Для меток, связанных со всеми остатками, наблюдается некоторое уменьшение фотоотклика, однако, на Суs-322, 323 этот эффект наиболее ярко выражен (рис. 3.16). При соотношении родопсин/трасдуцин, близком к эквимолярному, фотоотклик практически полностью исчезает.
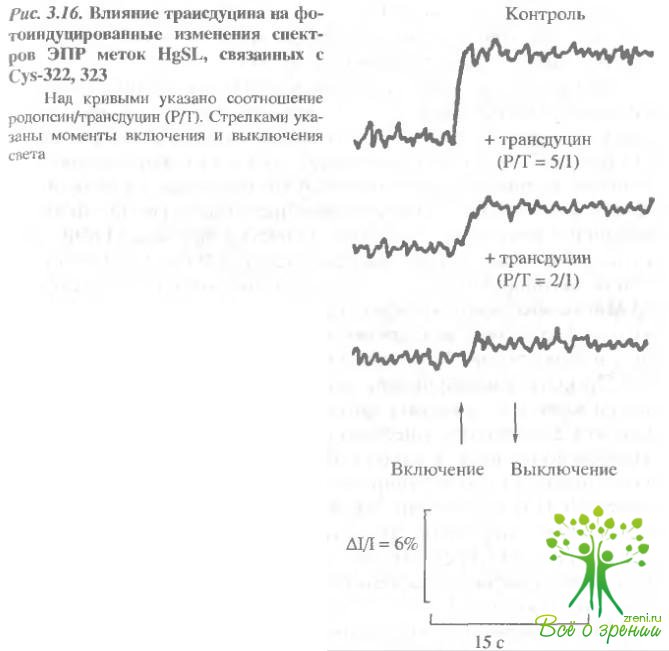
В этих же условиях изменения фотоотклика для меток МЕ8Ь, связанных с Суs-140, 316, составляют не более 30%. Возможно, это изменение связано с ограничением конформационных перестроек на поверхности родопсина при связывании трансдуцина. Добавление ГТФ приводит к восстановлению фотоответа, однако время нарастания его существенно замедляется.
Роль фосфорилирования родопсина в ингибирующем действии арестина на фосфодиэстеразу цГМФ. Нами показано, что арестин способен с достаточно высокой аффинностью связываться не только с фосфорилированным, но и с нефосфорилированным метародопсином II. Возникает вопрос, приводит ли связывание арестина с нефосфорилированным метародопсином II к ингибированию фосфодиэстеразы цГМФ.
Для ответа на этот вопрос определяли начальную скорость гидролиза цГМФ фосфодиэстеразой наружных сегментов палочек в условиях, когда V?Vмакс (концентрация цГМФ = 140 мкМ). На рис. 3.17 видно, что добавление арестина к нефосфорилированному родопсину в отсутствие АТФ не влияет на активность фосфодиэстеразы, а в присутствии предварительно фосфорилированного родопсина начальная скорость гидролиза цГМФ падает почти в 2 раза. Отсюда можно сделать вывод, что для проявления ингибирующего действия арестина на фосфодиэстеразу цГМФ необходимо фосфорилирование родопсина.
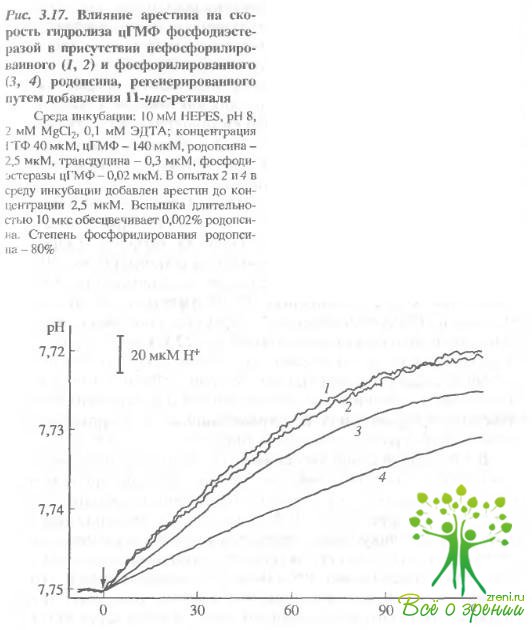
Тот арестин, который связывается с другим центром на неФосфорилированном метародопсине II, не проявляет ингибирующей активности.
Поскольку модификация остатков Cys -322 и Cys-323 существенно влияет на кинетику фотоотклика, можно предположить, что эти SH-группы существенны для связывания трансдуцина. Интересно оценить, в какой степени модификация этих цистеинов влияет на светоиндуцированную фосфодиэстеразную активность НСП (как известно, активация фосфодиэстеразы в НСП происходит при участи« трансдуцина)
На рис. 3.18 представлено влияние модификаций SH-групп родопсина на фосфодиэстеразную активность.
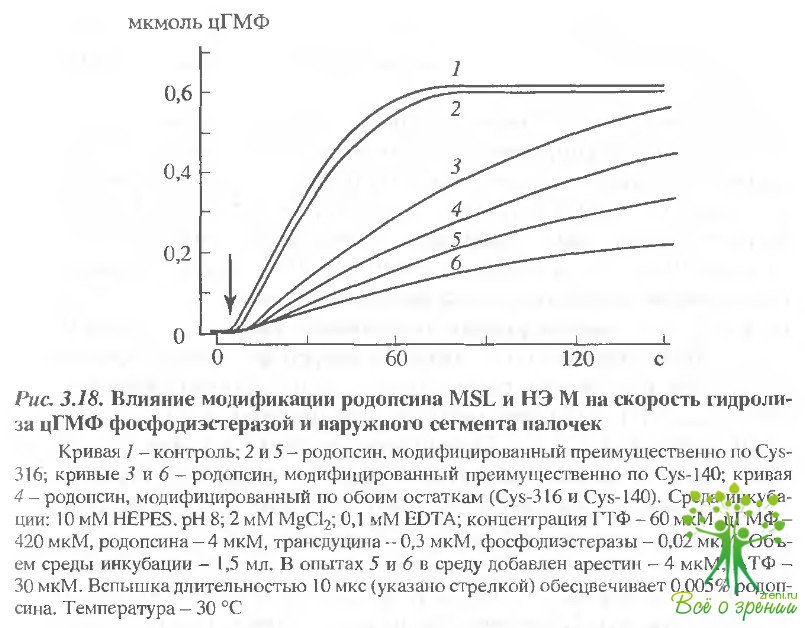
Преимущественная модификация Cys-316 не влияет на скорость гидролиза цГМФ. Модификация же второго остатка родопсина Cys-140 приводит к снижению фосфодиэстеразной активности (рис. 3.18, 3): начальная скорость гидролиза цГМФ уменьшается от 120 до 50 нмолей цГМФ/с. Наибольший эффект был получен при исчерпывающей модификации остатков Cys-322 и Cys-323 (рис. 3.19). Это приводило к снижению начальной скорости гидролиза цГМФ в 3,3 раза.

Таким образом, модификация метками Cys-140, а особенно С-концевых цистеиновых остатков, влияет на взаимодействие метародопсина II с трансдуцином, что проявляется в снижении фосфодиэстеразной активности.
В следующей серии экспериментов аналогичный подход был использован для выяснения влияния модификации цистеиновых остатков родопсина на степень ингибирования фосфодиэстеразы арестином (см. рис. 3.18,5, 6). Как известно, добавление арестина и АТФ в среду инкубации, содержащую опсинкиназу, приводит к значительному снижению активности фосфодиэстеразы. Это наблюдается и в наших экспериментах. Однако, в отличие от предыдущих экспериментов, модификация остатков Cys-140 и Cys-316 не влияла на ингибирование фосфодиэстеразной активности арестином (сравнить кривые 2 и 5, 3 и 6 на рис. 3.18). Добавление арестина и А1Ф приводило к снижению начальной скорости гидролиза цГМФ в 2.5-4 раза как для нативного, так и для модифицированного по различным схемам родопсина. Эти результаты согласуются с описанными выше данными о том, что сульфгид-рильные группы родопсина не расположены в месте связывания арестина.
Итак, каковы основные выводы из описанных выше исследований? Фотоиндуцировгнные изменения в спектрах ЭПР спинмеченного родопсина были показаны ранее только в растворах детергента. В фоторецепторных мембранах такие изменения не были обнаружены ни с использованием обычной ЭПР-спектроскопии, ни с использованием переноса насыщения. В данной работе нам удалось наблюдать такие изменения. Для того чтобы описать характер фотоиндуцированных изменений в молекуле родопсина, полезно сравнить изменения в спектрах ^ПР, индуцированные освещением и протеолитическим гидролизом белка. Освещение приводит к увеличению вращательной подвижности метки на Cys-140 и Cys-316, причем, максимальный эффект достигается на Cys-140. С другой стороны, подвижность тех же меток существенно изменяется и в ходе протеолиза. Отщепление С-концевого пептида вызывает такие же изменения как на Cys-140, так и на Cys-316. Таким образом, можно полагать, что С-концевой пептид расположен непосредственно у поверхности белковой глобулы. Это предположение подтверждается также тем, что подвижность меток, связанных с обоими цистеинами, возрастает при удалении С-концевого фрагмента. Таким образом, можно полагать, что в натпвном родопсине их подвижность ограничена С-концевым пептидом.
То, что характер изменений одинаков при освещении и удалении С-концевого фрагмента, позволяет предположить, что при фотолизе происходит '"отодвигании" С-концевого фрагмента от поверхности белковой глобулы.
Фосфорилированис родопсина приводит к противоположному эффекту - подвижность метки уменьшается, что происходит, по-видимому, вследствие того, что С-пептид оказывается ближе к петлям, содержащим Cys-140 в фосфорилированном родопсине. Хотя нельзя исключить и прямого взаимодействия фосфорилированных остатков со спиновыми метками.
Интересно отметить что расщенление родопсина на два пептида не приводит к исчезновению фотоэффекта, он даже несколько возрастает. Особенно это касается MSL, связанной с Cys-140. который расположен на фрагменте F24. хотя хромофор расположен во фрагменте F12. Таким образом, как расщепление белка на два фрагмента, так и удаление гидрофильных петель не нарушают вызываемых светом изменений конформации белка и взаимодействия между тяжами.
Арестин обладает достаточно высоким сродством только к обесцвеченным формам родопсина: кд комплексов арестина с темновым и обесцвеченным рецептором различаются на 2-3 порядка, причем, сродство арестина к метародопсину II выше, чем к продуктам его превращения в ходе фотолиза родопсина.
Хотя арестин способен связываться как с фосфорилирсванными, так и нефосфорилированными формами обесцвеченного родопсина, характеристики центров связывания в обоих случаях существенно различны.
Во-первых, как для метародопсина II, так и для опсина величина Кд комплексов с арестином у фосфорилированных форм рецептор? меньше, что свидетельствует о большей аффинности центров связывания.
Во-вторых, комплексы арестина с нефосфорилированным и фосфорилированным опсином различаются по структурно-динамическим параметрам связанного арестина. Так, подвижность спинмеченного арестина в комплексе с фосфорилированным опсином примерно в 2 раза ниже, чем с нефосфорилированным. Это свидетельствует о большей иммобилизации ареотина в центре фосформированного рецептора. Кроме того, различия в соотношениях W/S-сигналов ЭПР-метки можно интерпретировать как различия в конфирмационных состояниях арестина в этих комплексах.
В-третьих, связывание ареегина с фосфорилированным опсином является в основном электростатическим, так как существенно уменьшается при высокой ионной силе среды инкубации. В го же время на взаимодействие арестина с нефосфорилированным опсином ионная сила практически не влияет. Можно предположить, что отрицательно заряженные при рН 7 группы ортофосфата, связанные с остатками серина и треонина родопсина, принимают участие в формировании центра связывания арестина. Подтверждает это факт, что уже при 5-минутном протеолизе фосфорилированного родопсина, когда образуется преимущественно фрагмент без С-концевого участка, несущего фосфорилированные остатки, степень связывания арестина изменяется до значений, соответствующих взаимодействию с нефосфорилированным рецептором. При более глубоком иротеолизе (через 90 мин) т.е. при выщеплении цитоплазматической петли между V и VI ?-спиральными тяжами родопсина, когда преимущественно накапливаются два ковалентно не связанных фрагмента, происходит нарушение связывания арестина. Возможно, этот цитоплазматический участок также формирует центр связывания арестина. Ранее было показано, что именно этот участок существен и для связывании, транедуцина.
Наконец, наиболее существенно, что связывание арестина с нефосфорилированным метародопсином II, в отличие от связывания с его фосфопилированной формой, не приводит к ингибированию активности фосфодиэстеразы цГМФ, т.е., по-видимому, не препятствует связыванию и активации трасдуцина на рецепторе.
Ранее было показано, что фосфорилирование приводит к инактивации метародопсина II: в присутствии опсинкиназы и АТФ наблюдается быстрый спад "сигнала диссоциации". Этот эффект может быть обусловлен двумя причинами либо уменьшением сродства метародопсина II к трансдуцину-ГДФ, либо уменьшением скорости обмена ГДФ на ГТФ на трансдуцине. Наши данные указывают на то, что при фосфорилировании метародопсина II Кд возрастает почти на порядок - до 11,2 ± 0,2 мкМ. Таким образом, фосфорилирование рецептора приводит к нарушению его взаимодействия с трансдуцином.
При фосфорилировании метародопсина II образуется дополнительный высокоаффинный (Кд = 0,7 ± 0,1 мкМ) центр связывания арестина - центр, обеспечивающий сильную иммобилизацию арестина. В формировании этого центра принимают участие фосфорилированяые цитоплазматические области родопсина, отщепляемые при протеолизе папаином. Учитывая, что концентрация арестина в наружном сегменте палочки довольно велика и составляет примерно 15-100 молекул арестина на 100 молекул родопсина, т е. примерно в 2-10 раз превышает содержание трансдуцина, можно считать, что фосфооилирование метародопсина II создает эффективные условия для конкурентного вытеснения арестином трансдуцина из комплекса с рецептором. Можно предположить, что этот механизм является основным в ингибирующем действии арестина на фосфодиэстеразу цГМФ.
Эффект влияния трансдуцина и арсстина на спектры ЭПР позволяет рассмотреть характер взаимодействия родопсина с этими белками, а избирательная модификация сульфгидрильных групп - также локализовать их центры связывания.
Попытки оценить влияние модификации SH групп родопсина на его взаимодействие с трансдуцином предпринимались и ранее Так, в работе было показано, что модификация родопсина NEM не влияла на связывание родопсина с трансдуцином, наблюдаемое по сигналу "связывания" в светорассеянии. Можно предполагать, что в достаточно мягких условиях, используемых в работе, модификация Cys-140 была неполной. Кроме того, хотя измерение активности фосфодиэстеразы и не является прямым исследованием связывания трансдуцина, в действительности оно чувствительнее для определения влияния модификации на образование комплекса родопсин-трансдуцин.
Модификация Cys-322 и Cys-323 ранее не проводилась, однако можно сравнить полученные нами данные с результатами протеолитического отщепления С-концевого участка, содержащего эти остатки, что и будет сделано в дальнейшем.
Модификация Cys-316 в наших экспериментах не влияла на активность фосфодиэстеразы и константу связывания с арестином. Добавление трансдуцина также не влияло на спектры ЭПР MESL, связанной с Cys-316, однако некоторые изменения наблюдались после добавления арестина к фосфорилированному родопсину. Таким образом, можно заключить, что Cys-316 не включен в центр связывания трансдуцина, но расположен в непосредственной близости от центра связывания арестина. Это предположение согласуется и с результатами работы, в которой замена Cys-316 на серин в рекомбинантном родопсине не влияла на активацию фосфодиэстеразы.
Модификация Cys-140 приводит к значительному ингибированию активности фосфодиэстеразы, хотя и не влияет на спектры ЭПР, Это свидетельствует о том, что центр связывания трансдуцина непосредственно включает этот цистеиновый остаток Интересно отметить, что действие антител на пептид, содержащий Cys-140, также приводило к ингибированию активности фо сфодиэстеразы.
Парные С-концевые группы (Cys-322, 323), по-видимому, находятся в непосредственной близости от места посадки трансдуцина: их модификация существенно сказывается на активации фосфодиэстеразы. Однако связывание трансдуцина при спинмеченьи родопсина, по-видимому, не нарушается, поскольку метки все же чувствуют присутствие трансдуцина (как в темноте, так и при освещении). По-видимому, при модификации метками парных SH-групп может пооисходить изменение в диссоциации ?-субъединицы трансдуцина от родопсина. Существенное замедление кинетики фотоотклика в присутствии трансдуцина-ГТФ по сравнению с контрольными образцами без трансдуцина может быть проявлением этого факта.
Кроме того, можно предположить также, что при модификации парных SH-групп может измениться конформация всего С-концевого фрагмента, поскольку, как было показано в работе, эти группы в родопсине быка пальмитгрованы и заякоривают тем самым С-коьцеьой участок на мембране. При модификации этих групп алкилирующими ртутьорганическими агентами С-конец теряет фиксацию на мембране, и центр связывания трансдуцина может существенно изменить свою конформацию.
Интересно отметить также, что относительное уменьшение величины фотоответа нелинейно зависит от соотношения трансдуцин/родопсин (при т/р = 0,2 уменьшение интенсивности составляло 50%, а при т/р = 0,5 - примерно 80%). То же касается и замедления кинетики фотоответа. Этот факт предполагает кооперативность во взаимодействии транедуцина с родопсином, т.е. одна молекула родопсина может взаимодействовать с 2-3 молекулами родопсина. Насыщение сигнала связывания трансдуцина в работе наблюдалось при соотношении т/р примерно 0,25-0,30, что хорошо соотносится с нашими данными.
Для более точного определения мест связывания трансдуцина и арестина интересно сравнить полученные нами данные с эффектами протеолиза, а также с действием антител на взаимодействие родопсина с аранедуцином.
Отщепление от 12 до 19 (348-339) аминокислот не приводит к ингибированию активности фосфоциэстеразы. в то время, как синтетический пептид, включающий остатки 317-339, конкурирует с трансдуцином. Результаты, полученные с рекомбинантным родопсином с замененными Asp-330, Asp-333 и Lys-248, показывают, что эти замены сильно снижают функциональную активность родопсина.
Суммируя наши данные с приведенным выше, можно заключить, что центр связывания трансдуцина включает район 317-339, особенно петлю между III и IV тяжами, а также включает Cys-14H и Lys-248 в петле между V и VI тяжами. Заякоривание С-концевого пептида также имеет, по-видимому, важное значение для ГДФ-ГГФ-обмена на транедучине.
Хотя удаление пептида 330-348 не влияет на активность фосфодиэстеразы. фосфорилирсвание этою пептида существенно влияет на константу связывания трансдуцина, связывание ГТФ с транедуцином и активность фосфоциэстеразы. Таким образом, хотя этот участок прямо и не входит в центр связывания, однако появление отрицательных зарядов при фосформировании серинов и треонинов, локализованных в этой области, оказывает существенное влияние на взаимодействие с трансдуцином.
Модификация малеимидными и ртутьорганическими метками любой из поверхностных SН-групп родопсина, как показано нами, не влияет на связывание его с арестином. К тому же блокирование Суs-140 и Суs-316 не снимает активности арестина как ингибитора фосфодиэстеразы. Можно поэтому заключить, что при связывании арестина с родопсином участки вблизи Суs-140, Суs-316, Суs-322 и Суs-323 не затрагиваются. В то же время фосфорилирование С-конца родопсина существенно увеличивает связывание арестина, а отщепление фосфорилированного С-конца уменьшает его связывание и снимает ингибирующее действие арестина на фосфодиэстеразу. Это указывает на то что, в отличие от трансдуцина. значительная область С-концевой последовательности родопсина формирует место связывания арестина.
Таким образом, можно полагать, что оба белка связываются с С-концевой областью, однако связывание происходит в разных ее участках. Приведенные нами данные по химической модификации SН-трупп родопсина в совокупности с ранее известными данными позволяют предположить, что ингибирующее действие арестина не связано с его конкуренцией с трансдуцином за общие места связывания на родопсине, а осуществляется другим путем.
Статья из книги: Молекулярные механизмы зрительной рецепции | Каламкаров Г.Р., Островский М.А..

Комментариев 0